Longisquama
Longisquama („lange Schuppe“) war eine Gattung diapsider Reptilien, deren Fossilien in Sedimentgesteinen der mittel- bis obertriassischen Madygen-Formation im Südwesten Kirgisistans entdeckt wurden. Eine Anzahl mutmaßlicher verlängerter Schuppen, die am Rücken des einzigen bekannten Skelettexemplars ansetzten, wird als Anpassung an den Gleitflug gedeutet.
| Longisquama | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
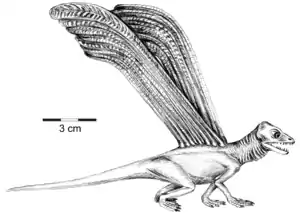
Lebendrekonstruktion von Longisquama auf der Grundlage des einzigen bekannten Skelettexemplars. Anordnung der verlängerten Rückenschuppen in Anlehnung an Haubold & Buffetaut (1987).[1] | ||||||||||
| Zeitliches Auftreten | ||||||||||
| Mitteltrias bis Obertrias | ||||||||||
| vor ca. 230 bis 220 Mio. Jahre | ||||||||||
| Fundorte | ||||||||||
| ||||||||||
| Systematik | ||||||||||
| ||||||||||
| Wissenschaftlicher Name | ||||||||||
| Longisquama | ||||||||||
| Sharov, 1970 | ||||||||||
| Art | ||||||||||
|
Fundlokalität und Typus-Exemplar
Die triassischen klastischen Sedimentgesteine der Madygen-Formation sind nahe der Ortschaft Madgyen im Lyailyaksii-Distrikt im äußersten Südwesten Kirgisistans (Gebiet Batken) an mehreren Stellen aufgeschlossen. Nach bisherigen Ergebnissen repräsentieren die Gesteine der auf 500 m Mächtigkeit geschätzten Abfolge Fluss- und Seeablagerungen, in denen auch einige Kohleflöze vorkommen.[2][3]

Neben Longisquama hat die Madygen-Formation eine Vielfalt an Makrophyten (Pflanzen-Großfossilien) und Insekten sowie Knochenfische (einschließlich eines Lungenfisches), Muscheln, Krebstiere, das Amphib Triassurus, die Therapsiden-Gattung Madysaurus sowie Sharovipteryx, einen weiteren kleinen Diapsiden, hervorgebracht. Bei Sharovipteryx und Longisquama, die beide aus den Ton-/Schluffsteinen der „Oberen Grauen Folge“ der Madygen-Formation stammen, sind die Umrisse des Körpers sehr detailliert als Abdruck erhalten (Hautschattenerhaltung), so dass man einzelne Schuppen unterscheiden kann.[4]
Das 1970 von Sharov als Longisquama insignis beschriebene ca. 15 cm lange Teilskelett (PIN 2584/4, Holotypus) ist das einzige bisher gefundene Skelettexemplar der Gattung. Es umfasst Platte und Gegenplatte, auf denen ein unvollständiger Schädel, der Nacken, der vordere Teil des Brustkorbs, der Schultergürtel, die Vordergliedmaßen sowie Hautabdrücke am Hals, entlang der Arme und des Rückens erhalten sind. Besonders auffällig sind längliche blattartige Körperanhängsel, die von der Mittellinie des Rückens ausgehen. Fünf weitere Exemplare dieser Körperanhängsel ohne Zusammenhang zu Skelettfunden sind bisher beschrieben worden.[5][4] Die Fossilien befinden sich im Paläontologischen Institut der Russischen Akademie der Wissenschaften in Moskau.
Anatomie
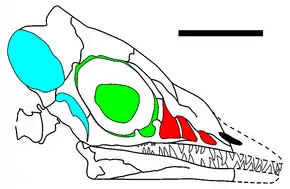

Longisquamas Schädel zeichnet sich durch den stark erweiterten Rand des oberen Temporalfensters (einer hinter dem Auge gelegenen Schädelöffnung) aus. Sharov beschrieb zwei Merkmale, die für die Gruppe der Archosaurier, zu der auch Krokodile und Dinosaurier gehören, als charakteristisch gelten: ein Schädelfenster, das sich zwischen Augen- und Nasenöffnung befindet (das Antorbitalforamen), sowie eine Öffnung im Unterkiefer (das Mandibularforamen). Das Vorhandensein dieser Öffnungen und die daraus abgeleitete systematische Zuordnung wurden unter anderem von Unwin & Benton (2001) in Zweifel gezogen.[6] Jones et al. (2001) hingegen bestätigen ausdrücklich das Vorliegen eines Antorbitalforamens,[7] nach der Interpretation von Peters (2000) sind sogar drei nach vorn kleiner werdende Antorbitalfenster zu unterscheiden (siehe Abbildung).[8]
Anders als bei Archosauriern, aber ähnlich wie bei Schuppenechsen (Lepidosauriern), waren Longisquamas Zähne offenbar acrodont, das heißt ohne Zahnwurzeln an den Kieferknochen befestigt.[4] Der kurze Hals besteht aus acht Halswirbeln, für die der Nachweis von Rippen uneindeutig ist. Die ersten der länglichen blattartigen Körperanhängsel setzen oberhalb der hinteren Halswirbel an. Es liegen acht dünne nach hinten in der Länge zunehmende Rippen des Brustkorbs und eine entsprechende Anzahl von Rückenwirbeln vor, der sich schwanzwärts anschließende Teil des Körpers ist nicht erhalten.[8]
Der Schultergürtel besteht aus langen schmalen Schulterblättern, kurzen stabförmigen Rabenbeinen, zwei robusten Schlüsselbeinen, die zusammen eine U-Form bilden und mit einem Zwischenschlüsselbein (Interclavicula) verbunden sind. Unwin & Benton (2001) verweisen darauf, dass das Vorhandensein eines verknöcherten Zwischenschlüsselbeins wiederum untypisch für Archosaurier sei.[6] Jones u. a. (2001) entgegnen, dass Zwischenschlüsselbeine auch in einer Anzahl von Archosauriern wie zum Beispiel Euparkeria vorkommen. Sie bezeichnen die beiden Schlüsselbeine aufgrund ihrer Form und ihres Überlappens als ein Gabelbein, das in seiner Form demjenigen des Urvogels Archaeopteryx gleiche.[7] Peters (2000) beschreibt ein Brustbein, das von den beiden Schlüsselbeinen eingefasst sei, sowie eine Art Kiel, der vom Zwischenschlüsselbein ausgehe. Er sieht in der besonderen Morphologie des Schultergürtels von Longisquama jedoch keine Affinität zu Vögeln, sondern betont die Übereinstimmung des Komplexes aus Schlüsselbeinen, Zwischenschlüsselbein und Brustbein mit dem eines juvenilen Exemplars des urtümlichen Flugsauriers Eudimorphodon.[8]
Die Oberarme sind schlank, leicht S-förmig gebogen und sind in etwa genauso lang wie die Unterarm-Knochen. Longisquamas Hände sind asymmetrisch - für die Finger Nr. I bis V beträgt die Zahl der Fingerglieder 2, 3, 4, 5 bzw. 4. Die zweitäußersten Glieder der Finger (diejenigen, an denen die Krallen ansetzen) sind überproportional lang, ein Merkmal das oft infolge der Anpassung an das Klettern auftritt. Ein für die Gattung Longisquama charakteristisches (autapomorphes) Merkmal ist der extrem verlängerte vierte Finger, der dem Oberarm in der Länge gleicht.[4]
Schuppen oder Federn?
Die bis zu 15 cm langen blattartigen vom Rücken abstehenden Körperanhängsel sind nur von den Longisquama zugeordneten Exemplaren aus der Madygen-Formation bekannt. Sharov (1970) deutete sie als verlängerte Hornschuppen und gab der neuen Gattung einen entsprechenden Namen. Er nahm an, dass sie eine aerodynamische Funktion hatten und ähnlich wie Fallschirme funktionierten. Haubold & Buffetaut (1987) entwickelten ein Modell, nach dem die Anhänge paarig angeordnet waren und als ausklappbare Tragflächen den Gleitflug ermöglichten (siehe Abbildung in der Box).[1]
Erst Jones et al. (2000) zweifelten daran, dass die Anhänge tatsächlich Reptilienschuppen repräsentierten - sie erklärten, es handele sich um Federn eines „Nichtvogels“, die homolog zu Vogelfedern seien und diskutierten verschiedene Beobachtungen, die diese These stützen sollten. Die Mittelachse jeder Struktur repräsentiere demnach den Federschaft (Rhachis) und die sonst als Fältelung oder Streben gedeuteten Abzweigungen von der Mittelachse wurden als Strahlen (Radii) gleich den Strahlen einer Vogelfeder gedeutet. Die Federfahne, die sich wie bei Vögeln aus einzelnen Strahlen zusammensetze, sei dieser Vorstellung nach bei bisherigen Beschreibungen als massive Schuppe fehlinterpretiert worden, tatsächlich seien die „Strahlen“ bei Longisquama nur am äußeren Rand miteinander verwachsen, so dass die „Federn“ einen glatten Umriss und nicht, wie zu erwarten, einen ausgefransten Rand zeigen.[9]
Jones et al. (2001) führten als Beleg die Überkreuzung mancher „Strahlen“ an, die bei der Deutung als massive Schuppe nicht zu erklären sei. Die hohlzylindrische Basis der Anhänge, die gleich der Spindel (Federkiel, Calamus) echter Vogelfedern intern transversal gegliedert seien, ist ihrer Ansicht nach einer der wichtigsten Beweise. Solche Strukturen könnten nur durch das Wachstum aus einem Federfollikel heraus erklärt werden, der gleiche Wachstumsmechanismus wie bei Federn sei daher anzunehmen. Des Weiteren sei nahe der Basis eine Umhüllung der „Federn“ mit einer Struktur, die der Federscheide moderner Federn gleiche, zu beobachten. Angesichts des Vorliegens einer „follikularen Federbasis“, eines „Federkiels“, einer „Federscheide“, eines „Federschafts“ sowie einer aus „Strahlen“ bestehenden „Federfahne“ schlussfolgerten Jones et al. (2000, 2001), dass es sich bei den Körperanhängen von Longisquama um Federn handeln müsse.[9][7]
Ein großer Teil der mit dem Ursprung der Vögel vertrauten Paläontologen hält diese Ansicht für eine Fehlinterpretation, die unter anderem auf dem Überinterpretieren beobachteter Gewebeabdrücke oder auch auf der Missachtung widersprüchlicher Befunde beruht. So seien zum Beispiel nirgends Häkchen (Rami) nachzuweisen, die bei Kontur- und Schwungfedern die einzelnen Strahlen miteinander verbinden (so dass eine kontinuierliche Federfahne vorliegt). Randlich fusionierte Vogelfedern seien in der rezenten Vogelwelt ein sehr seltenes Phänomen und im vorliegenden Fall eine weit hergeholte Erklärung. Die als „Strahlen“ gedeuteten Strukturen der Anhängsel hätten z. T. keine Ähnlichkeit mit denen von Vogelfedern und seien an vielen Stellen nicht nachzuweisen. Prum (2001) stimmt mit Jones et al. (2000) überein, dass die Körperanhängsel von Longisquama eine zylindrische Basis haben, sieht jedoch nicht die Notwendigkeit, dass sie von Federfollikeln (oder von mit Federfollikeln homologen Strukturen) gebildet wurden.[10][11]
Ergänzend zur Skepsis gegenüber den von Jones et al. zu Longisquama veröffentlichten Beobachtungen und Schlussfolgerungen, ist eine Mehrheit der Paläontologen von der Vielzahl gemeinsamer abgeleiteter Merkmale und von den Ergebnissen vieler kladistischer Analysen überzeugt, die Vögel als direkte Nachfahren theropoder Dinosaurier ausweisen. Fossilien gefiederter Dinosaurier zeigen verschiedene Zwischenstadien zwischen haarartigen Körperanhängseln und komplexen Vogelfedern mit Federfahne, die sich mit Hilfe der Entwicklungsbiologie erklären lassen.[12] Die für Dinosaurier diskutierten Stadien der Federentwicklung lassen sich auch im Laborversuch durch Manipulation der verantwortlichen Morphogene erzeugen.[13] Aus diesen Gründen und da keine phylogenetische Analyse vorliegt, die Longisquama tatsächlich in die nähere Verwandtschaft der Vögel stellt, findet die These, die mutmaßlichen verlängerten Schuppen hätten etwas mit der Entwicklung der Federn zu tun, in der Regel nur unter Gegnern der Dinosaurierabstammung der Vögel Unterstützung.[11]
Systematik
Jones et al. (2001) gehen davon aus, dass der Nachweis eines Antorbitalfensters (eine vor der Augenhöhle liegende Schädelöffnung) die Einordnung der Gattung Longisquama in die Gruppe der Archosaurier, wie sie bereits von Sharov bei der Erstbeschreibung vollzogen wurde, rechtfertigt. Ihrer Ansicht nach legen die Übereinstimmung der Körperanhängsel mit Vogelfedern und die Konfiguration der Schlüsselbeine, die dem Gabelbein der Vögel gleicht, nahe, dass innerhalb der Archosaurier eine Verwandtschaft Longisquamas mit den Vögeln bestehe.[7]
Im Gegensatz dazu hält Peters (2000) das bzw. die Antorbitalfenster von Longisquama und einigen anderen Diapsiden nicht notwendigerweise für ein Merkmal der Zugehörigkeit zu den Archosauriern, sondern für eine Folge konvergenter Evolution. Er untersucht Longisquama, Sharovipteryx, verschiedene Gattungen der Prolacertiformes und urtümliche Flugsaurier mit dem Ziel einer Revision bisheriger kladistischer Analysen und erhält das folgende Ergebnis (vereinfacht):
| Archosauromorpha |
| ||||||||||||||||||||||||||||||||||||||||||
Neben dem Vorhandensein von drei Antorbitalfenstern, die in späteren Formen zu einem zusammenwachsen, diskutiert Peters eine Anzahl weiterer Gemeinsamkeiten, auf denen das neue Taxon „Fenestrasauria“ beruhen soll.[14]
Zweifel an der Robustheit dieser Klassifikation Longisquamas sind aufgrund der Versehrtheit und Mehrdeutigkeit des Schädels und Schultergürtels, auf deren Interpretation das Analyseergebnis im Wesentlichen beruht, angebracht. Der mutmaßliche Prolacertiforme Sharovipteryx zeigt Ähnlichkeiten mit Flugsauriern, deren phylogenetische Signifikanz jedoch umstritten ist. Nach derzeit mehrheitlicher Auffassung gehören Flugsaurier zu den Archosauriern und bilden gemeinsam mit den Dinosauriern sowie Scleromochlus und Lagosuchus die Gruppe Ornithodira.[15]
Eine im Jahr 2004 von Senter veröffentlichte Analyse stellt Longisquama als fragliches Mitglied in die Gruppe Avicephala, einer neu definierten Untergruppe der Diapsiden, der auch der Gleitflieger Coelurosauravus angehören soll. Das folgende Kladogramm stellt diese Verwandtschaftshypothese vereinfacht dar:
| Diapsida |
| ||||||||||||||||||||||||||||||
Die Einheit der Ornithodira als Teilgruppe der Archosaurier bliebe demnach unangetastet.[16]
Lebensweise
Longisquama war ein kleiner arboricoler (baumbewohnender) Insektenfresser, dessen spitze gekrümmte Zähne zum Knacken von Chitin-Exoskeletten geeignet waren. Der relativ hoch entwickelte Schultergürtel, die Proportionen der Gliedmaßenknochen und die verlängerten äußeren Fingerglieder, an denen die leicht gekrümmten Krallen ansetzten, zeigen gute Kletterfähigkeiten an. Wenn die mutmaßlichen verlängerten Schuppen Longisquama tatsächlich zum Gleitflug befähigten, könnten sie sich besonders bei der Fortbewegung von Baum zu Baum als vorteilhaft erwiesen haben. Andernfalls könnten sie der Abschreckung von Feinden oder der Zurschaustellung (als Teil des Paarungsverhaltens) gedient haben.[4]
Siehe auch
Populärwissenschaftliche Literatur
- Barry Cox, Dougal Dixon, Brian Gardiner, Robert J. G Savage: Dinosaurier und andere Tiere der Vorzeit. Die große Enzyklopädie der prähistorischen Tierwelt. Mosaik-Verlag, München 1989, ISBN 3-570-03928-5.
Referenzen
- Hartmut Haubold, Éric Buffetaut: A new interpretation of Longisquama insignis, an enigmatic reptile from the Upper Triassic of Central Asia. In: Comptes Rendus de l'Académie des Sciences. Série 2: Mécanique, Physique, Chimie, Astronomie, Sciences de la Terre et des Planètes. Bd. 305, 1987, ISSN 1477-2019, S. 65–70.
- Inna A. Dobruskina: Keuper (Triassic) Flora from Middle Asia (Magygen, Southern Ferghana) (= New Mexico Museum of Natural History and Science. Bulletin. 5, ISSN 1524-4156). New Mexico Museum of Natural History and Science, Albuquerque NM 1995, S. 1–49, online.
- Dmitry E. Shcherbakov: Madygen, Triassic Lagerstätte number one, before and after Sharov. In: Alavesia. Bd. 2, 2008, ISSN 1887-7419, S. 113–124, Digitalisat (PDF; 586,68 kB) (Seite nicht mehr abrufbar, Suche in Webarchiven) Info: Der Link wurde automatisch als defekt markiert. Bitte prüfe den Link gemäß Anleitung und entferne dann diesen Hinweis. .
- David M. Unwin, Vladimir R. Alifanov, Michael J. Benton: Enigmatic small reptiles from the Middle-Late Triassic of Kirgisztan. In: Michael J. Benton, Mikhail A. Shishkin, David M. Unwin, Evgenii N. Kurochkin: The Age of Dinosaurs in Russia and Mongolia. Cambridge University Press, Cambridge u. a. 2000, ISBN 0-521-55476-4, S. 177–186.
- Александр Г. Шаров: Своеобразная рептилия из нижнего триаса Ферганы. In: Палеонтологический Журнал. Nr. 1, 1970, ISSN 0031-031X, S. 127–131, [Ein ungewöhnliches Reptil aus der Unteren Trias von Fergana.], (in russischer Sprache und kyrillischer Schrift).
- David M. Unwin, Michael J. Benton: Longisquama Fossil and Feather Morphology. In: Science. Bd. 291, Nr. 5510, 2001, S. 1900–1901, doi:10.1126/science.291.5510.1899c.
- Terry D. Jones, John A. Ruben, Paul F. A. Maderson, Larry D. Martin: Longisquama Fossil and Feather Morphology. In: Science. Bd. 291, Nr. 5510, 2001, S. 1901–1902, doi:10.1126/science.291.5510.1899c
- David Peters: A reexamination of four prolacertiforms with implications for pterosaur phylogenesis. In: Rivista Italiana di Paleontologia e Stratigrafia. Bd. 106, Nr. 3, 2000, ISSN 0035-6883, S. 293–336.
- Terry D. Jones, John A. Ruben, Larry D. Martin, Evgeny N. Kurochkin, Alan Feduccia, Paul F. A. Maderson, Willem J. Hillenius, Nicholas R. Geist, Vladimir Alifanov: Nonavian Feathers in a Late Triassic Archosaur. In: Science. Bd. 288, Nr. 5474, 2000, S. 2202–2205, doi:10.1126/science.288.5474.2202.
- Richard O. Prum: Longisquama Fossil and Feather Morphology. In: Science. Bd. 291, Nr. 5510, 2001, S. 1899–1900, doi:10.1126/science.291.5510.1899c.
- Erik Stokstad: Feathers, or Flight of Fancy? In: Science. Bd. 288, Nr. 5474, 2000, S. 2124–2125, doi:10.1126/science.288.5474.2124.
- Richard O. Prum: Development and Evolutionary Origin of Feathers. In: Journal of Experimental Zoology. Bd. 285, Nr. 4, 1999, S. 291–306, doi:10.1002/(SICI)1097-010X(19991215)285:4<291::AID-JEZ1>3.0.CO;2-9.
- Mingke Yu, Ping Wu, Randall B. Widelitz, Cheng-Ming Chuong: The morphogenesis of feathers. In: Nature. Bd. 420, Nr. 6913, 2002, S. 308–312, doi:10.1038/nature01196.
- Einige Synapomorphien der Fenestrasauria nach Peters (2000): Verschmelzen des Ectopterygoids und Pterygoids (Knochen, die am Aufbau des Gaumens beteiligt sind), ein ausgedehntes Rostrum (Schnauze) und Palatinum (Gaumenbein) infolge der Verlängerung des Prämaxillare (Zwischenkieferknochen); Rückenwirbel mit vergrößerten seitwärts gerichteten Fortsätzen (Quer- oder Transversalfortsätzen); mindestens vier Beckenwirbel; Überlappen der Schlüsselbeine; Zwischenschlüsselbeine mit reduziertem Schaft; ein strebenartiges Rabenbein, das mit dem Brustbeinkomplex verbunden ist; ein verlängerter vierter Finger, der mindestens dreimal so lang ist wie der vierte Mittelhandknochen. Darüber hinaus sei nur bei Longisquama und den Flugsauriern das Brustbein mit den Schlüsselbeinen fusioniert.
- zur Gegenüberstellung siehe zum Beispiel Christopher A. Brochu: Progress And Future Directions In Archosaur Phylogenetics. In: Journal of Paleontology. Bd. 75, Nr. 6, 2001, ISSN 0022-3360, S. 1185–1201, Digitalisat (PDF; 685,05 kB).
- Phil Senter: Phylogeny of Drepanosauridae (Reptilia: Diapsida). In: Journal of Systematic Palaeontology. Bd. 2, Nr. 3, 2004, S. 257–268, doi:10.1017/S1477201904001427.