Totenkopfschwärmer
Der Totenkopfschwärmer (Acherontia atropos) ist ein Schmetterling (Nachtfalter) aus der Familie der Schwärmer (Sphingidae), dessen Hauptverbreitungsgebiet die Tropen Afrikas sind. Er kommt auch im äußersten Süden Europas vor, von wo aus er als Wanderfalter nach Mittel- und Nordeuropa fliegt. Die Art ist gemeinsam mit ihren beiden Verwandten aus der Gattung Acherontia innerhalb der Schwärmer einzigartig, da sie durch einen Mechanismus in der Mundhöhle pfeifende Geräusche erzeugen kann. Auch die Lebensweise der Falter, sich von Honig zu ernähren und dafür in Bienenstöcke einzudringen, ist sehr ungewöhnlich.
| Totenkopfschwärmer | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Totenkopfschwärmer (Acherontia atropos) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Acherontia atropos | ||||||||||||
| (Linnaeus, 1758) |


Auf Grund seiner Lebensweise und der imposanten Erscheinung mit dem namensgebenden „Totenkopf“ auf dem Thorax galt er lange Zeit als unheilbringend. Er wurde 1988 in dem Roman Das Schweigen der Lämmer von Thomas Harris für das Böse stilisiert. Der Erstbeschreiber, Carl von Linné, gab der Art den wissenschaftlichen Namen Atropos nach einer griechischen Göttin.
Merkmale
Falter
Die Falter erreichen eine Flügelspannweite von 90 bis 115 mm (Männchen) bzw. 100 bis 122 mm, maximal 130 mm (Weibchen), wobei in Mitteleuropa aufgewachsene Tiere in der Regel etwas größer sind als jene, die aus dem Süden zuwandern. Ihr vollständig und nahezu pelzig dicht beschuppter Körper ist bis ca. 60 mm lang und erreicht einen Durchmesser von etwa 20 mm. Damit ist der Totenkopfschwärmer die größte in Europa vorkommende Schwärmerart und zählt hier auch zu den größten vorkommenden Schmetterlingen überhaupt. Männchen erreichen ein Gewicht von 2 bis 6 g und Weibchen von 3 bis 8 g. Die verhältnismäßig kurzen Fühler sind beim Männchen 10 bis 14,5 mm, beim Weibchen 10 bis 13 mm lang. Sie sind schwarz gefärbt und haben eine weiße Spitze.[1] Das Männchen besitzt ein spitz zulaufendes Hinterleibsende und ist hierdurch vom Weibchen zu unterscheiden, bei dem dieses stumpf abgerundet ist.[2]
Der Kopf und die Oberseite des Thorax sind schwarzbraun oder nahezu schwarz. Auf dem Thorax kann man die charakteristische totenkopfähnliche Zeichnung erkennen, der die Art ihren deutschen Namen verdankt. Diese gelbe Zeichnung ist variabel und fehlt manchmal vollständig.[3] Die Unterseite des Thorax und des Hinterleibs ist ockerfarben. Jedes Sternit auf der Unterseite des Hinterleibs trägt eine breite dunkle Querbinde; die Terga auf der Rückenseite des Abdomens sind seitlich ocker bis nahezu orangefarben und tragen mittig einen graublauen Längsstreifen. Der Hinterrand der Terga ist schwarz. Dadurch ergibt sich eine markante Doppelreihe aus ocker- bzw. orangefarbenen Flecken am Hinterleib. Die letzten zwei, seltener drei Hinterleibssegmente sind beim Männchen entweder komplett graublau oder schwarz. Beim Weibchen hat nur das letzte Segment eine solche Färbung.[4]
Die Vorderflügel sind tiefbraun bis dunkelgrau schattiert und mit rotbraunen bis ockerfarbenen, mitunter weißlichen Flecken marmoriert. In der Flügelmitte befindet sich ein kleiner heller Punkt, der sich zwischen zwei schwarzen Querbinden befindet. Direkt an der Flügelbasis sind die Falter hell ockergelb behaart. Nahe der Flügelbasis und in der Postdiskalregion liegen helle ockergelbe Querbinden, von denen die letztere zum Flügelaußenrand deutlich rostbraun verfärbt ist. Die Enden der Flügeladern sind am Flügelaußenrand auch rostbraun gefärbt.[1] Die variable Musterung der Flügel kann sehr deutlich ausgebildet sein oder nahezu vollständig fehlen, wodurch die Vorderflügel dann nahezu gleichmäßig braun wirken. Auch der helle Punkt ist variabel ausgebildet. Die Hinterflügel sind auf der Oberseite ockerfarben und haben zwei auffällige dunkle Querbinden. Ihre Färbung ist ebenfalls sehr variabel. Die Binden können graubraun aufgehellt oder so stark ausgeprägt sein, dass sie nahezu miteinander verschmelzen. Die innere Binde kann gleich breit ausgebildet sein wie die Binde nahe am Flügelaußenrand, man findet jedoch auch Individuen mit unterschiedlich breit ausgebildeter innerer Binde, bis hin zum völligen Fehlen dieser.[2] Die Unterseite beider Flügelpaare ist überwiegend ockerfarben. Mittig auf den Vorderflügeln findet sich eine feine schwarze Binde, der Flügelaußenrand ist dunkel. Die Binde setzt sich auf der Unterseite der Hinterflügel fort, hinzu kommt dort anstelle des dunklen Außenrandes eine undeutlich abgegrenzte dunkle Submarginalbinde, die über die Flügeladern an den Flügelaußenrand ausstrahlt. Die innere Binde ist nahe dem Innenrandwinkel zum Außenrand hin gekrümmt.[1] Die Vorderflügelfärbung ermöglicht den Tieren eine perfekte Tarnung auf Baumrinde oder trockenem Laub am Boden, wo sie tagsüber mit dachförmig aneinandergelegten Flügeln ruhen.[3]
Wie für alle drei Arten der Gattung Acherontia typisch, aber unter den Schwärmern sonst einzigartig, haben die Tiere mit 12 bis 18 mm Länge einen verhältnismäßig kurzen, aber sehr breiten und stabilen Saugrüssel. Er besteht nicht wie bei anderen Schmetterlingen aus einer Röhre, sondern ist bandartig und füllt auch nur die Hälfte der Rüsselscheide aus. An der Basis hat er eine Breite von 0,75 bis 1,25 mm und verjüngt sich zu einem spitzen Ende, das anders als der restliche Rüssel ungezähnt ist. Diese Rüsselspitze ist auch besser beweglich als bei anderen Schwärmerarten. Die Tiere besitzen weiters innenseitig an den Tibien (Schienen) der Vorderbeine eine Putzschuppe, die aus einem schwammig wirkenden, mit Borsten besetzten Plättchen besteht. Die Fühler werden geputzt, in dem sie zwischen dem Plättchen und der Tibia hindurchgezogen werden.[2]
Ei
Die Eier des Totenkopfschwärmers sind matt hellgrün oder blaugräulich und haben auf ihrer Oberfläche eine sehr feine polygonale Netzstruktur, die jedoch nur bei starker Vergrößerung erkennbar ist. Sie sind 1,5 bis 1,7 mm breit, 1,7 bis 1,9 mm lang und damit schwach oval. Ihre Hülle ist sehr elastisch, so dass die Eier nach einem Herabfallen vom Boden wieder hochfedern. Mit der Entwicklung des Embryos verfärbt sich das Ei nach und nach von gelblich zu gelb und ist zwei Tage vor dem Schlupf der Raupe etwas eingedellt. Kurz vor dem Schlupf kann man durch die Eischale hindurch die Mundwerkzeuge und das Analhorn der Raupe erkennen.[3][5]
Raupe




Die Raupen erreichen eine Körperlänge von 120 bis 130 mm und treten ausgewachsen im Wesentlichen in einer grün-gelblichen, braunen oder gelb-orangen Grundfarbe auf. Sie sind nach dem Schlupf rund sechs Millimeter lang und haben dann eine hellgelbliche Färbung, die durch feine, sehr eng beieinander liegende, blassgelbe Tuberkel und Linien matt überdeckt wird. Ihr Körper ist spärlich behaart. Das mit 2,6 bis 2,8 mm Länge zur Körperlänge verhältnismäßig lange Analhorn hat eine gegabelte Spitze und verfärbt sich ein bis zwei Stunden nach dem Schlupf schwarz. Die Kopfkapsel hat dieselbe Färbung wie der Körper und ist 0,9 bis 1 mm breit. Nach Beginn der Nahrungsaufnahme an den Blättern verfärbt sich der Körper der Raupen rasch grün. Vor der ersten Häutung haben die Raupen eine durchschnittliche Körperlänge von 12 mm.[6]
Im zweiten Raupenstadium sind die Tiere 12 bis 17 mm lang, haben eine gelblichgrüne bis grüne Körperfärbung und tragen auch auf der 1,5 bis 1,6 mm breiten Kopfkapsel deutlich erkennbare, eng beieinander stehende, feine, helle, dornförmige Tuberkel und Körnchen, insbesondere am Thorax. Ab diesem Raupenstadium treten nach und nach die seitlichen Schrägstreifen am Körper auf. Das schwarze Analhorn verläuft gerade und ist 4 bis 5 mm lang. Seine Spitze läuft in ein oder zwei Enden aus.[7]
Nach der zweiten Häutung haben die Tiere eine Körperlänge von 18 bis 30 mm und sind hellgrün bis gelblich. Sie tragen nach wie vor helle Tuberkel sowie Warzen, die am Thorax mehr und mehr hervortreten. Am Hinterleib haben sich nunmehr markante, unterseits weiße und oberseits gelbe Schrägstreifen ausgebildet, die sich am Rücken beidseits jeweils zu einem „V“ treffen. Zwischen dieser Zeichnung befinden sich dunkelgrüne bis bläuliche Schrägstreifen. Das Analhorn ist dicker als im vorhergehenden Raupenstadium und zum meist ein-, selten zweispitzigen Ende hin gekrümmt. Es ist basal dunkel gefärbt und wird zur Spitze hin hell gelb-grünlich. Die erste Hälfte ist vor allem auf der Rückenseite mit dunklen Stacheln versehen. Die Kopfkapsel ist 2,7 bis 2,8 mm breit.[7]

Ab dem vierten Raupenstadium sind die Tiere bereits 40 bis 45 mm, maximal 50 mm lang und wiegen vor der letzten Häutung etwa vier Gramm. Sie haben eine gelbe oder grüne Grundfarbe. Die Kopfkapsel ist 4,2 bis 4,5 mm breit, am Kopf sind seitlich schwarze Streifen ausgebildet. Die Thorakalbeine sind schwarz und mit feinen Warzen weiß punktiert. Die Schrägstreifen am Hinterleib sind gelb und blau bis violettblau gefärbt. Am Rücken des ersten Hinterleibssegments sind dunkelviolette Punkte ausgebildet. Der Thorax ist am Rücken deutlich hell beborstet. Die Stigmen sind schwarz. Das 6 bis 7 mm lange, nunmehr gelbe Analhorn ist an der Spitze bereits nach oben und somit S-förmig gekrümmt. Es tragt auffällige gelbe Stacheln.[7]
Im fünften und letzten Raupenstadium sind die Raupen nach der Häutung 50 bis 70 mm lang und entwickeln sich bis zur Verpuppung zu ihrer eingangs beschriebenen Länge; ausgestreckt erreichen sie sogar 150 mm. Sie wiegen dann 18 bis 22 g. Ihre Kopfkapsel ist 7 bis 8 mm breit, der Kopf ist seitlich schwarz gefärbt, die Stirn trägt oberseits zwei dunkle, nach unten etwas auseinanderzeigende Streifen. Die Mundwerkzeuge sind ebenso schwarz gefärbt. Die Körperoberfläche ist anders als in den vorhergehenden Stadien glatt. Sie ist in den eingangs erwähnten drei Farbvarianten gefärbt, wobei dunkelbraune Tiere sehr selten auftreten. Die „V“-Zeichnung am Hinterleib ist rückenseitig blau bis violett, bauchseitig leuchtend hellgelb umrandet. Am Hinterleibsrücken finden sich zahlreiche schwarze Punkte, die Stigmen sind deutlich schwarz hervortretend. Die Thorakalbeine sind wie zuvor schwarz und tragen weiße Warzen. Das nur 5 bis 7 mm lange, gelbe Analhorn ist charakteristisch für die Art wulstig, grob bestachelt und deutlich S-förmig gekrümmt. Nach jeder Häutung dauert es immer eine kurze Zeit, bis die neue Körperfärbung ausgebildet ist.[8]
Puppe


Die Puppe ist beim Männchen 50 bis 65 mm lang, beim Weibchen 65 bis 70 mm, maximal 80 mm. Die frische Puppe ist gelb bis cremefarben und kann am Rücken leicht grünlich angehaucht sein. Nach etwa 12 Stunden ist sie stark glänzend und mahagonifarben. Sie wiegt beim Männchen 7 bis 10 g, beim Weibchen 7 bis 12 g. Der Saugrüssel ist mit dem Hinterleib bündig verschmolzen. Der Kremaster ist unter dem Rasterelektronenmikroskop als stark gefaltet erkennbar. Am siebten Segment finden sich in der Puppenhaut einige wenige mikroskopisch kleine Senken, in deren Mitte Borstenhärchen sitzen.[9]
Ähnliche Arten
Der Totenkopfschwärmer ist kaum mit anderen Schwärmerarten zu verwechseln, lediglich Acherontia styx, dessen Verbreitungsgebiet sich im Nahen Osten mit dem des Totenkopfschwärmers überschneidet, sieht ihm ähnlich. Diese Art unterscheidet sich vom Totenkopfschwärmer durch zwei statt einer mittigen dunklen Binde auf der Unterseite der Vorderflügel, einen orangen, statt gelben Punkt auf der Mitte der Vorderflügel, eine dunklere Totenkopfzeichnung sowie einen schwach bläulichen Fleck im Analwinkel jeweils auf der Oberseite der Hinterflügel, der von der schwarzen Submarginalbinde umschlossen wird.[10]
Verbreitung und Lebensraum
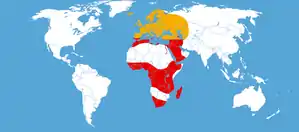
Gesamtverbreitung
Der Totenkopfschwärmer ist im Wesentlichen eine Art der Afrotropis, kommt aber auch in Nordafrika, dem Mittelmeerraum und dem Nahen Osten vor. Seine Verbreitung reicht im Osten von der Türkei bis in den Nordosten des Irans sowie in die Ukraine, nach Turkmenistan, Kuwait und den Saudi-Arabien. Ein einzelnes Männchen wurde 2006 im Nordosten Kasachstans (Pawlodar) gefunden.[11] In Europa kommt die Art an den südlichsten Küsten des Mittelmeeres und auf den mediterranen Inseln sowie auf Madeira, den Kanarischen Inseln und den Azoren dauerhaft vor. Sie fliegt jedoch gelegentlich im Sommer als Wanderfalter weiter nach Norden und kann dabei auch weit über Frankreich und Italien nach Nordeuropa und auch nach Island vordringen. Die nördlichste Verbreitung in Russland ist aus dem Ort Izvail in der Republik Komi nachgewiesen. Die Wanderflüge der Art reichen in den afrikanischen Tropen bis zur Insel Ascension.[12]
Lebensraum
Die Tiere besiedeln offene, verbuschte Lebensräume, in denen Nachtschattengewächse wachsen, insbesondere auch Regionen, in denen Kartoffeln landwirtschaftlich angebaut werden. Sie bevorzugen dabei trockene und sonnige Gegenden.[3] Zumindest in Mitteleuropa ist die Art überwiegend zum Kulturfolger geworden und besiedelt fast ausschließlich Kartoffelfelder.[13]
Die Falter halten sich bevorzugt in den niederen Höhenlagen auf,[14] so sind etwa aus Baden-Württemberg Falterfunde vom Flachland bis in die hochmontane Stufe des Schwarzwalds um 900 Meter Seehöhe dokumentiert.[15] Während ihrer Wanderflüge fliegen die Tiere aber auch deutlich höher, so wurde die Art in den Alpen beispielsweise in Graubünden in 3000 Meter, in der Silvretta auf 2000 Meter Seehöhe, jeweils auf Gletschern, oder am Timmelsjoch in knapp 2500 Meter Seehöhe nachgewiesen.[16] Raupen und Puppen findet man auf Grund ihres hohen Wärmebedarfs jedoch nur deutlich tiefer, in Baden-Württemberg etwa bis maximal in die untere montane Stufe bis zu 700 Meter Seehöhe,[15] in Bayern bei 730 Metern Seehöhe.[17]
Lebensweise
Wie auch die anderen Arten der Gattung Acherontia können sowohl die Männchen als auch die Weibchen des Totenkopfschwärmers schrille, pfeifende Geräusche erzeugen. Durch Kontraktion von zwei kräftigen mit dem Schlund (Pharynx) verbundenen Muskeln wird die Pharynxdecke gehoben und Luft durch den Saugrüssel eingesogen. Gleichzeitig wird die Speiseröhre (Ösophagus) verschlossen, sodass die Luft nur die Mundhöhle füllt. Ein Teil der Luft streicht dabei auch durch die Mundöffnung, wobei der Epipharynx aufwärts gebogen wird, um durch einen Depressormuskel wieder nach unten gedrückt zu werden. Durch rasches Abwechseln dieses Vorgangs wird die Mundöffnung abwechselnd kurz geöffnet und geschlossen und so werden Schallwellen, ähnlich wie in einer mechanischen Sirene erzeugt. Das Schwingen des Epipharynx erzeugt so einen Ton zwischen 6000 und 8000 Hz, wobei diese Trägerfrequenz mit etwa 280 Impulsen pro Sekunde moduliert wird. Ein so erzeugter Laut dauert etwa eine Sechstelsekunde und hat somit 40 bis 50 Impulse. Durch das Erschlaffen der Muskeln am Pharynx wird die Luft wieder durch den Saugrüssel nach draußen befördert, wobei ein weiterer Laut entsteht, der zwar die gleiche Frequenz hat, jedoch nicht moduliert ist, da der Epipharynx nicht schwingt und so ein gleichmäßiger Luftstrom erzeugt wird. Der so entstehende Ton dauert nur eine Sechzehntelsekunde, ist schwächer und mehr pfeifend; der Hauptton während des Einatmens ist lauter und klingt kratzender.[18]
Die Falter pfeifen meist nur, wenn sie stark beunruhigt werden, etwa bei Berührung. Insbesondere bei Männchen ist die Lauterzeugung auch in der Nähe eines Weibchens oder kurz nach der Ankunft an einer Lichtquelle zu vernehmen.[18][12]
Werden die Falter gestört, entfalten sie ihre Flügel und laufen hüpfend unruhig umher und erzeugen pfeifende Geräusche. Sie fliegen jedoch nicht ab, sondern kriechen unter Pflanzenteile oder in Löcher und Spalten. Bei starker Störung sondern Männchen einen Stoff, der nach modernden Pilzen riecht, aus Drüsen am Sternit des zweiten Hinterleibssegmentes ab. Dieser wird durch pinselartige Haarbüschel, die abgespreizt werden, besser verteilt.[12] Auch wenn künstliche Lichtquellen abgeschaltet werden, suchen dort ruhende Falter Verstecke auf, anstatt wegzufliegen. Sie tun dies dann auf gleiche Weise, wie sie in Bienenstöcke eindringen. Um die Flügelmuskulatur aufzuwärmen, belassen die Falter die Flügel zusammengefaltet über dem Hinterleib und vibrieren mit den Flügeln nur leicht auf und ab. Dies ist eine Anpassung an die Lebensweise in Bienenstöcken; Schwärmer entfalten ihre Flügel dafür ansonsten über dem Rücken.[19]
Nahrung der Falter

Die Imagines sind auf der Suche nach Nahrung oder Geschlechtspartnern ab der Dämmerung bis nach Mitternacht aktiv. Die Falter werden von künstlichen Lichtquellen angelockt, wobei jedoch fast nur Männchen oder selten Weibchen kurz vor der Eiablage anfliegen. Die Tiere fliegen auch gelegentlich die Blüten von verschiedenen Pflanzen, wie etwa Kartoffel, Tabak, Bartnelke, Heckenkirschen, Pfeifensträucher oder Orange an. Sie können sich jedoch auf Grund der Form ihres Saugrüssels nicht von Blütennektar ernähren und berühren aus diesem Grund die Blüten nicht. Schmetterlingsköder werden nicht angeflogen. Anders verhält es sich mit Pflanzensaft, der aus Wunden austritt. Diese Nahrungsquelle wird aber aller Wahrscheinlichkeit nach nur im Notfall genutzt. Ebenso dürfte es sich gegebenenfalls mit dem vermuteten Anstechen von reifem Obst verhalten, das aber anders als bei der nahe verwandten Art Acherontia styx[10] beim Totenkopfschwärmer nicht dokumentiert ist. Die Nahrungsaufnahme ist für die Totenkopfschwärmer nicht nur für ihr Überleben notwendig, sondern auch für die Eireifung bei den Weibchen erforderlich.[12][20]
Wichtigste Nahrungsquelle des Totenkopfschwärmers sind die Nester der Westlichen Honigbiene (Apis mellifera), in die die Falter eindringen, um Honig und Nektar zu saugen. Auffällig ist, dass die Arbeiterinnen der Bienen den Faltern gegenüber keine Aggressivität zeigen. Zunächst dachte man, dass die Totenkopfschwärmer die Bienen durch ihre Pfeifgeräusche besänftigen,[20] tatsächlich geschieht dies jedoch durch die Abgabe von chemischen Substanzen, die den fremden Geruch der Falter tarnen. Der Geruchsstoff der Falter besteht aus einer Mischung von vier Fettsäuren, nämlich Palmitoleinsäure, Palmitinsäure, Stearinsäure und Ölsäure, die auch nahezu in der gleichen Konzentration und im gleichen Verhältnis bei Honigbienen auftreten. Das Mischungsverhältnis ist dabei in allen Körperteilen der Falter und bei Männchen wie Weibchen gleich. Dadurch, dass die Falter den gleichen Geruch wie die Bienen haben, werden sie nicht als Eindringlinge erkannt.[21] Wenn überhaupt, werden die Eindringlinge nur durch die Wächter am Eingang des Nestes attackiert, vor denen sie durch ihre dicke Körperhülle gut geschützt sind. Auch macht ihnen das Bienengift nur wenig aus.[21] In Versuchen überlebten Falter die Injektion einer Giftmenge, die vier Bienenstichen entsprach, unbeschadet.[16] Einmal in das Nest eingedrungen, verharren die Falter zunächst ruhig auf den Waben sitzend und werden von den Bienen ignoriert. Sie klettern unbehelligt mit schwirrenden Flügeln auf den Waben umher, stoßen dabei die Bienen mit den Vorderbeinen beiseite und schütteln jene, die auf sie klettern mit Körper- und Flügelbewegungen ab. Schließlich stechen sie ihren starken Saugrüssel in gedeckelte wie ungedeckelte Zellen und saugen im Schnitt in etwa 15 Minuten ungefähr fünf von ihnen leer, um den Stock danach wieder zu verlassen. Es kommt auch vor, dass die Mittelwand der Zellen durchstochen wird und die Falter die auf der anderen Seite gelegene Zelle so auch aussaugen können. Der Energieverbrauch der Falter ist im Flug enorm. In Ruhe werden in einem Gramm der Flugmuskulatur pro Minute 0,06 µmol Glucose zu Glucose-6-phosphat umgesetzt, während des Fluges sind es 3,9 µmol, was 0,7 Milligramm entspricht.[20]
Wanderflüge, Flug- und Raupenzeiten
Der Totenkopfschwärmer ist ein Wanderfalter, der regelmäßige und weite Wanderungen unternimmt. Jahr für Jahr fliegen die Tiere in stark schwankender Zahl von Afrika nach Süd- und Mitteleuropa, einige sogar nach Nordeuropa ein. Diese Einflüge erfolgen mit stark schwankenden Individuenzahlen. Jahre, in denen die Falter regelmäßig und häufig auftreten, werden durch solche abgelöst, in denen die Falter offenbar gar nicht einfliegen. Raupen und Puppen findet man weit häufiger als Falter;[22] die Raupen treten gelegentlich auch in Massen auf.[12]
Acherontia atropos tritt in Afrika ganzjährig in kontinuierlich aufeinander folgenden Generationen auf. In Europa findet man die ersten Falter meist ab Mai, gelegentlich auch schon ab März/April, gemeinsam mit den vereinzelten Individuen, die eine Überwinterung als Puppe überlebt haben. Dieses gleichzeitige Auftreten bewirkt, dass eine Herkunftsbestimmung der Falter nur sehr schwer möglich ist. Die erste Einflugwelle wird im Laufe des Juni schwächer, die zweite Welle folgt dann im August und September.[14] Die Eier der Weibchen reifen während des Fluges nach Norden. Ist die Reifung abgeschlossen, fliegen die Falter nicht mehr weiter und legen ihre Eier rasch ab. Diese ersten Falter finden meist noch keinen ausreichenden Kartoffelbewuchs, weswegen auf andere Nahrungspflanzen ausgewichen wird. Erst die ab Juli einfliegenden Falter belegen auch Kartoffelpflanzen mit ihren Eiern. Im Mai oder Juni auftretende Kälteeinbrüche, wie etwa die Eisheiligen oder die Schafskälte, beeinträchtigen die Populationen stark, sodass die Eier oder jungen Raupen in großer Zahl zu Grunde gehen. Sie vertragen länger anhaltende Temperaturen unterhalb von 15 °C nicht, können aber ungefähr sechs Tage lang Nachttemperaturen zwischen 5 und 8 °C und Tagestemperaturen um 16 °C überstehen. Die Falter sind temperaturtoleranter und konnten in den Alpen auch bei Schneetreiben im Flug beobachtet werden. Sind die Temperaturen von April bis Juni günstig, können sich die Raupen in Mitteleuropa in vier bis fünf Wochen entwickeln. Demnach treten die ersten adulten Raupen in solchen Jahren ab Mitte bis Ende Juni auf. Im Durchschnitt benötigen die Raupen acht Wochen für die Entwicklung und legen anschließend eine einmonatige Puppenruhe ein. Somit tritt die nächste Faltergeneration ab September auf und verstärkt die zeitgleich einfliegende zweite Einflugwelle.[16] In der Regel findet man Raupen in Europa von Juli bis September und Puppen von September bis Oktober.[22] Auf Gran Canaria treten bereits im Dezember und Januar Eier und Raupen auf.[16]
Nahrung der Raupen

Die Raupen ernähren sich vor allem von Pflanzen aus der Familie der Nachtschattengewächse und insbesondere von der Kartoffel (Solanum tuberosum). Andere wichtige Nahrungspflanzen derselben Familie sind Bittersüßer Nachtschatten (Solanum dulcamara), Lycium europaeum, Gemeiner Bocksdorn (Lycium barbarum), Schwarze Tollkirsche (Atropa belladonna), Virginischer Tabak (Nicotiana tabacum) und Gemeiner Stechapfel (Datura stramonium),[12] sowie Blasenkirschen (Physalis), Tomate (Solanum lycopersicum), Schwarzer Nachtschatten (Solanum nigrum) und Aubergine (Solanum melongena).[12][23]
Die Raupen sind jedoch polyphag und fressen darüber hinaus an einer Vielzahl von Pflanzen aus etlichen Familien, darunter:[12][23]
- Moschuskrautgewächse: Holunder (Sambucus), Gemeiner Schneeball (Viburnum opulus)
- Doldenblütler: Dill (Anethum), Möhren (Daucus)
- Hundsgiftgewächse: Oleander (Nerium oleander),
- Korbblütler: Kanadisches Berufkraut (Conyza canadensis)
- Trompetenbaumgewächse: Catalpa bignonioides, Tecomaria capensis
- Kreuzblütengewächse: Kohl (Brassica),
- Hanfgewächse: Hanf (Cannabis sativa)
- Geißblattgewächse: Heckenkirschen (Lonicera), Schneebeeren (Symphoricarpos)
- Spindelbaumgewächse: Spindelsträucher (Euonymus)
- Fuchsschwanzgewächse: Rübe (Beta vulgaris)
- Hülsenfrüchtler: Ackerbohne (Vicia faba)
- Hortensiengewächse: Europäischer Pfeifenstrauch (Philadelphus coronarius)
- Lippenblütler: Mönchspfeffer (Vitex agnus-castus)
- Ölbaumgewächse: Eschen (Fraxinus), Jasmin (Jasminum), Liguster (Ligustrum), Ölbäume (Olea), Phillyrea, Flieder (Syringa)
- Malvengewächse: Hibiskus (Hibiscus)
- Paulownien: Blauglockenbaum (Paulownia tomentosa)
- Wegerichgewächse: Löwenmäuler (Antirrhinum)
- Rosengewächse: Weißdorne (Crataegus), Wald-Erdbeere (Fragaria vesca), Äpfel (Malus), Pflaume (Prunus domestica), Kultur-Birne (Pyrus communis)
- Rötegewächse: Färber-Meier (Asperula tinctoria)
- Rautengewächse: Orange (Citrus × aurantium), Rauten (Ruta)
- Braunwurzgewächse: Sommerflieder (Buddleja), Königskerzen (Verbascum)
- Brennnesselgewächse: Brennnesseln (Urtica)
Aus Anatolien wurden die Raupen darüber hinaus an Bohnenähnlichem Jochblatt (Zygophyllum fabago) nachgewiesen,[23] auf den Kanaren findet man sie auch an Cussonia, Afrikanischem Tulpenbaum (Spathodea campanulata), Tabebuia, Cordia sebestena, Blaugrünem Tabak (Nicotiana glauca) und Losbäumen (Clerodendrum).[12]
Die Raupen ernähren sich während ihrer gesamten Entwicklung meistens von der Nahrungspflanze, auf der sie geschlüpft sind. Nahrungspflanzenwechsel sind, anders als etwa beim Linienschwärmer (Hyles livornica), möglich, können aber mitunter zum Tod der Raupen führen.[23]
In alter Literatur wird die Art „Stechapfelschwärmer“ genannt oder die Raupen werden, wie etwa bei Rösel von Rosenhof im 18. Jahrhundert, als „Jasmin-Raupen“ bezeichnet. Dies geschah, da die heute wichtigste Nahrungspflanze, die Kartoffel, erst um 1565 erstmals von Amerika nach Europa importiert wurde und erst ab dem 18. Jahrhundert der Anbau in weiten Teilen Europas in großem Stil erfolgte. Der Totenkopfschwärmer war aber davor schon in Europa heimisch, wenn auch nicht in der heute dank des Kartoffelanbaus herrschenden Häufigkeit. Die Raupen fraßen in dieser Zeit an anderen Nachtschattengewächsen. Behauptungen, dass der Totenkopfschwärmer erst durch die in Europa eingeführte Kartoffel auftrat, sind unrichtig.[24]
Aus Europa sind aus jüngerer Zeit keine nennenswerten Schäden durch Raupenfraß dokumentiert. Historisch gab es jedoch immer wieder Jahre, in denen die Tiere so stark auftraten, dass sie Schäden, insbesondere an Kartoffelfeldern verursachten, so etwa 1905 in Norfolk, oder 1950 in großen Teilen Mitteleuropas, wo die Raupen, etwa in Thüringen, sogar mit amtlicher Anordnung gezielt bekämpft wurden.[16] Bei Massenauftreten sind Schäden auch aus Nordafrika und Südeuropa bekannt, so etwa 1955/56 in Orangenplantagen in Tunesien, oder 1973 in Griechenland auf Olivenbäumen.[23]
Paarung und Eiablage
Die männlichen Falter sind schon direkt nach dem Schlupf paarungsbereit und können sich bereits nach dem ersten Flug mit einem ebenso frisch geschlüpften Weibchen paaren. Meistens geschieht dies jedoch erst nach einigen Tagen, zumal frisch geschlüpfte Weibchen häufig versuchen, zu flüchten oder Pfeifgeräusche ausstoßen. Paarungsbereite Weibchen verharren ruhig sitzend und stülpen ihre Duftdrüse am Hinterleibsende aus, um Männchen anzulocken. Die Paarung erfolgt häufig an den Ruheplätzen der Falter, also auf Baumrinde oder am Boden auf trockenem Laub. Sie findet in der Regel am späten Abend ab etwa 22 Uhr statt, kann jedoch bis in die Morgenstunden erfolgen. Das Männchen steuert das Weibchen im Flug oder am Boden laufend direkt an, hält sich an einem Flügelvorderrand fest und vereinigt sich mit ihm, während es leicht schief sitzt. Das Paar ist schließlich Seite an Seite sitzend am Hinterleib aneinander gekoppelt, und ist, wie sonst bei Schwärmern üblich, mit dem Körper in entgegengesetzte Richtungen nur selten verbunden. Die Paarung dauert meist ein bis drei, selten bis zu fünf Stunden. Männchen paaren sich mitunter mit mehreren Weibchen hintereinander. Die befruchteten Eier sind nach ungefähr acht Tagen ablagebereit.[12][19]
Bereits einige Tage davor fliegen die Weibchen potentielle Nahrungspflanzen der Raupen an und zeigen Ablageverhalten. Sie legen schließlich in ein bis fünf Wochen jeweils etwa 10 bis 30, insgesamt im Durchschnitt 150 Eier ab. Maximal können Weibchen etwa 200 Eier legen, davon bis zu 75 pro Tag. Die Ablage erfolgt hauptsächlich einzeln auf der Unterseite älterer Blätter.[5]
Entwicklung

Wie auch bei anderen wandernden Schwärmerarten, etwa dem Oleanderschwärmer (Daphnis nerii) oder dem Linienschwärmer (Hyles livornica), liegt die kritische Photoperiode für die Entwicklung auch beim Totenkopfschwärmer bei Kurztagen mit ungefähr 14 Stunden Licht. Bei Raupen, die sich unter längeren Tagen entwickeln, steigt die Wahrscheinlichkeit, dass die sich aus ihnen entwickelnden Falter unfruchtbar sind, keine oder wenige Eier bilden, oder diese nach der Ablage zu Grunde gehen. Raupen, die sich im Sommer in Mitteleuropa oder weiter nördlich entwickeln, sind 15 und mehr Stunden Licht pro Tag ausgesetzt, aus ihnen entwickeln sich daher nur selten fortpflanzungsfähige Falter.[25]
Die Raupen schlüpfen je nach Temperatur nach fünf bis 12 Tagen. Bei 20 °C benötigen sie ungefähr acht Tage. Die frisch geschlüpften Raupen fressen zunächst ihre Eischalen, um sich anschließend meist auf der Unterseite der Blätter an einer Blattader zum Fressen zu positionieren. Sie leben anfänglich in Gruppen und fressen kleine Löcher in die Blätter (Lochfraß). In den Fraßpausen ruhen die Raupen auf Unterseite der Blätter. Ab dem dritten Raupenstadium sitzen sie beim Ruhen charakteristisch mit aufgerichtetem Vorderkörper und klammern sich dabei mit dem Nachschieber und den letzten beiden Bauchbeinpaaren an der Mittelrippe fest. Nach einiger Zeit der Entwicklung fressen die Raupen als Einzelgänger die Blätter vom Rand her ab. Sie sind nicht nur nachts aktiv, sondern fressen auch tagsüber und sind während ihrer gesamten Entwicklung extrem träge. Sie bewegen sich nur, wenn sie ein neues Blatt zum Fressen benötigen, wodurch nach einiger Zeit auf dem Bereich, wo die Raupe auf der Pflanze sitzt, eine Kahlstelle verursacht wird. Durch ihre Färbung sind sie ausgezeichnet getarnt, sodass selbst große, ausgewachsene Raupen meist nur schwer auf den Pflanzen entdeckt werden können. Werden sie gestört, erzeugen sie mit ihren Mandibeln klickende Geräusche und können ihre Angreifer mit den Mundwerkzeugen auch verletzen. Vor jeder Häutung verharren die Raupen einige Stunden bewegungslos auf der Blattunterseite, wobei sie als junge Tiere besonders auf der Mittelrippe, als ältere, größere Tiere an den Stängeln sitzen. Häufig wird die abgelöste Raupenhaut nach der Häutung mitsamt dem Analhorn aufgefressen.[25]
Sind die Raupen verpuppungsbereit, nehmen sie keine Nahrung mehr zu sich und verfärben sich innerhalb von wenigen Stunden von vorne beginnend in ein dunkles und trübes Gelb-Orange. Sie befeuchten während dieser Zeit ihren Körper mit Speichel, welcher den Verfärbungsprozess offenbar beschleunigt.[12] Sie machen sich dann auf die Suche nach einem geeigneten Platz zur Verpuppung und sondern während ihres Marsches sowohl aus dem Mund, als auch aus dem After eine alkalische Flüssigkeit ab, die auch beim Bau der Puppenkammer abgesondert wird. Die Verpuppung erfolgt im Erdboden eingegraben ab einer Tiefe von etwa 3 cm, meist zwischen 15 und 40 cm. Abhängig von der Temperatur ruht die Raupe im Erdboden einige Zeit, bevor sie sich endgültig verpuppt. Diese Ruhezeit dauert bei 20 °C sechs bis sieben Tage, bei 10 bis 14 °C bis zu 16 Tage. Die Verpuppung erfolgt in einer eiförmigen Höhle, die durch Bewegungen der Raupe auf der Innenseite relativ glattwandig wird. Sie hat etwa die Ausmaße von acht mal sechs Zentimetern. Die Puppe ist sehr aktiv und beweglich. Wenn sie gestört wird, vollführt sie schlagende und drehende Bewegungen. Kurz vor dem Schlupf können auch die Puppen bereits kurze, einfache „Pieps“-Töne ohne Wiederholungen erzeugen. Die Überwinterung findet für gewöhnliche im Puppenstadium statt, wobei die Tiere jedoch europäische Winter nur in Ausnahmefällen überleben. Die Dormanz der Überwinterung wird durch tiefe Temperaturen unter 5 bis 7 °C kurz nach der Verpuppung induziert. Ist dieser Zustand einmal ausgelöst, schlüpfen die Falter erst nach auf mehrmonatige kühle Temperaturen folgende Temperaturen über 20 °C und reagieren nicht auf einen kurz darauf folgenden Temperaturanstieg. Findet die Puppenruhe bei Temperaturen statt, die nicht unter 15 bis 17 °C fallen, tritt die Dormanz als Quieszenz auf, das heißt, die Entwicklung wird zwar verzögert, fertig entwickelte Falter schlüpfen aber unabhängig von der Dauer der tiefen Temperaturen, sobald die Temperaturen wieder über 20 °C steigen. Dies tritt in Mitteleuropa unter Berücksichtigung der Bodentiefe, in der die Verpuppung stattfindet nicht vor Mai ein, was bei der anschließend notwendigen vier bis sechswöchigen Entwicklungsdauer bis zum adulten Falter einen Schlupf erst ab Juni/Juli bedeutet. Dieser späte Schlupf hat zur Folge, dass die Eier dieser Falter erst im Juli/August ablagebereit sind, was wiederum eine Raupenentwicklung bis Ende September bewirkt. Diese Raupen sind Kurztagen mit weniger als 14 Stunden Licht ausgesetzt und damit wiederum fortpflanzungsfähig.[16] Die Eiablage oder Rückwanderung dieser Falter ist nach Traub in Ebert jedoch nicht dokumentiert, sie ergänzen durch ihr Auftreten lediglich die eingewanderten Populationen, die ansonsten mit ihrem Wanderflug allein für das kontinuierliche Auftreten der Art nördlich von Südeuropa verantwortlich sind.[22] Reinhardt und Harz sprechen jedoch von mehreren dokumentierten „Rückfliegern“,[16] was auch Weidemann und Köhler bestätigen.[26]
Der Schlupf der Falter erfolgt normalerweise am Abend zwischen 18 und 21 Uhr, nur selten kann er auch später oder sogar tagsüber erfolgen. Der Zeitpunkt ist abhängig vom Temperaturzyklus der die Puppe umgebenden Erde. Ist es tagsüber wärmer als in der Nacht, schlüpfen die Falter am Abend, ist es umgekehrt, am Morgen. Die Falter kriechen nach dem Schlupf durch die Erde ins Freie und suchen laufend nach einer geeigneten Stelle, an der sie nach oben klettern können. Dort lassen sie ihre Flügel nach unten hängen, diese sind nach etwa 40 Minuten voll entfaltet und nach insgesamt zwei Stunden auch ausgehärtet. Normalerweise verlassen die Falter diesen Ruheplatz frühestens am nächsten Abend, lediglich um ein besseres Versteck aufzusuchen, verlassen sie den Platz früher.[19]
Spezialisierte Feinde
Am Totenkopfschwärmer sind verschiedene Arten von Schlupfwespen (Ichneumonidae) und Raupenfliegen (Tachinidae) als Parasitoide nachgewiesen:[12]
- Ichneumonidae
- Amblyjoppa fuscipennis
- Amblyjoppa proteus
- Callajoppa cirrogaster
- Callajoppa exaltatoria
- Diphyus longigena
- Diphyus palliatorius
- Ichneumon cerinthius
- Netelia vinulae
- Tachinidae
- Compsilura concinnata
- Drino atropivora
- Masicera pavoniae
- Winthemia rufiventris
Taxonomie und Systematik

Der Totenkopfschwärmer wurde 1758 von Carl von Linné in der 10. Auflage seines Werks Systema Naturae als Sphinx atropos erstbeschrieben, Jacob Heinrich Laspeyres stellte die Art 1809 in die von ihm neu aufgestellte Gattung Acherontia, der die Art heute noch zugerechnet wird.[27] Der Gattungsname Acherontia ist von Acheron, einem der fünf Flüsse der Unterwelt aus der griechischen Mythologie abgeleitet. Das Artepitheton leitet sich von der Schicksalsgöttin Atropos ab, einer der drei Moiren aus der griechischen Mythologie, deren Aufgabe es ist, den Lebensfaden zu zerschneiden. Linné nahm mit der Vergabe des wissenschaftlichen Namens auf den damals verbreiteten Aberglauben Bezug. Auch die Namen der zwei weiteren Arten der Gattung haben Bezug zur griechischen Unterwelt: Styx ist ein Fluss der Unterwelt, Lachesis ist eine weitere Moire.[28]
Die Gattung Acherontia wird gemeinsam mit vier anderen Gattungen in die Tribus Acherontiini gestellt, deren Monophylie gut begründet ist. Die Verwandtschaftsverhältnisse zwischen den Gattungen innerhalb der Tribus sind nicht vollends erforscht, sicher scheint jedoch, dass Acherontia am nächsten mit der Gattung Coelonia verwandt ist, die demnach die Schwestergruppe bildet. Die drei Arten der Acherontia sind dabei von Coelonia gut abgegrenzt. Anhand von morphologischen Untersuchungen von Imagines, Raupen, Puppen und Raupennahrungspflanzen konnte gezeigt werden, dass Acherontia atropos mit Acherontia styx nächstverwandt ist, das Schwestertaxon der beiden Arten ist Acherontia lachesis.[28]
Es ergeben sich in der Gattung Acherontia also folgende Verwandtschaftsverhältnisse:
| |||||||||||||
Gefährdung und Schutz
Aufgrund seiner weiten Verbreitung und Häufigkeit ist der Totenkopfschwärmer nicht gefährdet. Als Wanderfalter wird er in Mitteleuropa in den Roten Listen gefährdeter Arten meist nicht geführt, in Deutschland ist er als Wanderfalter gelistet und als ungefährdet eingestuft. Die Art ist in der Bundesartenschutzverordnung (BArtSchV) nicht erfasst.[29]
Die Art hat auf Grund ihrer Vorliebe für Kartoffelpflanzen in Europa hohe Verluste durch die Ernte sowohl im Raupen-, als auch im Puppenstadium zu verzeichnen. Auch die in der Landwirtschaft verwendeten Insektizide tun dazu ihr Übriges.[30] Da das Vorkommen in Europa jedoch allein durch aus dem Süden einwandernde Tiere gewährleistet wird, haben diese Verluste keine Auswirkung auf die heimischen Populationen und ist die Art auch deshalb nicht in den Roten Listen erfasst.[29]
Nach den Erfahrungen früherer Schmetterlingssammler ließen sich Totenkopfschwärmer in den seinerzeit üblichen „Tötungsgläsern“ mit im Glasboden eingegipstem Zyankali praktisch nicht einschläfern, auch nicht mit dem sonst üblichen Gemisch von Äther und Chloroform. Dagegen bewährte sich immer wieder die Injektion eines in eine Spritze aufgezogenen Suds aus Zigarettentabak.
Kulturelle Rezeption

Der Totenkopfschwärmer galt durch seine markante Zeichnung, insbesondere den namensgebenden „Totenkopf“ am Thorax, bei dem gelegentlich sogar Andeutungen von gekreuzten Beinknochen erkennbar sind, als „Totenvogel“. Unter diesem Namen fand er bereits 1719 in der „Breslauischen Kunst- und Naturgeschichte“ seinen Eintrag. Auch sein Auftreten in Bienenstöcken und die nachtaktive Lebensweise verstärkten seinen Ruf als unheilbringend. Dort wo er sich in Häuser verirrte, befürchtete man Unheil.[31]

Der Totenkopfschwärmer fand aber auch in die moderne Mythologie Einzug.
- In Luigi Bertellis Kinderbuch Ciondolino (zuerst italienisch 1895, deutsch: Max Butziwackel der Ameisenkaiser, 1920) hilft der in eine Ameise verwandelte Max einem Bienenschwarm dabei, den Angriff eines Totenkopfschwärmers auf das Bienennest abzuwehren.
- Im Roman Dracula (1897) des irischen Autors Bram Stoker sendet Graf Dracula diese Schmetterlingsart zu Renfield, dem verwirrten Insassen einer Anstalt.
- Der Totenkopfschwärmer kommt auch im Film Ein andalusischer Hund (original französisch: Un chien andalou) von Luis Buñuel und Salvador Dalí vor,[32] der zum ersten Mal 1929 in Paris aufgeführt wurde.
- Auch in Thomas Harris' Buch Das Schweigen der Lämmer (USA 1989) und in dem darauf basierenden Film (USA 1991, Regie: Jonathan Demme, Drehbuch: Ted Tally) tritt der Totenkopfschwärmer in Erscheinung. Der Serienmörder „Buffalo Bill“ platziert die Puppe der Falter im Mund seiner Opfer;[28] die Titelseite des Buchs und das Filmplakat zeigen den Totenkopfschwärmer.
Aber auch der Schüler von Carl von Linné, James Barbut, philosophierte anhand des Umgangs von Arbeitskollegen und Entomologen mit dem Totenkopfschwärmer über Tierethik und wissenschaftliche Neugierde wie folgt: "The ſphinx atropos ſqueakes when hurt, nearly as loud as a mouſe, which, when uttered in the moſt plaintive tone, naturally ſhocks the human heart, and makes it ſhudder at the thought of deſtroying inoffenſive animals merely for the ſake of curioſity. I cannot help reflecting on this tyranny, this wanton cruelty, exerciſed by thoughtleſs man, on many animals, but eſpecially in inſects: 'tis certain, that every animal poſſeſſing life, has feeling; and, therefore, is as capable of ſuffering pain, as of enjoying pleaſure; and, as Shakeſpeare humanely expreſſes “The poor beetle cruſhed beneath the foot, feels the pangs of death as great as when a monarch falls.” Gentle reader, pardon this digreſſion, my feelings commanded my pen."[33]
Quellen
Literatur
- Arno Bergmann: Die Großschmetterlinge Mitteldeutschlands. Band 3: Spinner und Schwärmer. Verbreitung, Formen und Lebensgemeinschaften. Urania-Verlag, Jena 1953, DNB 450378365.
- Günter Ebert: Die Schmetterlinge Baden-Württembergs. Band 4, Nachtfalter II (Bombycidae, Endromidae, Lemoniidae, Saturniidae, Sphingidae, Drepanidae, Notodontidae, Dilobidae, Lymantriidae, Ctenuchidae, Nolidae). Ulmer, Stuttgart 1994, ISBN 3-8001-3474-8.
- J. J. de Freina, Th. Witt: Die Bombyces und Sphinges der Westpalaearktis. Band 1. Noctuoidea, Sphingoidea, Geometroidea, Bombycoidea. EFW Edition Forschung & Wissenschaft Verlag, München 1987, ISBN 3-926285-00-1.
- Ian J. Kitching, Jean-Marie Cadiou: Hawkmoths of the World. An Annotated and Illustrated Revisionary Checklist (Lepidoptera: Sphingidae). Cornell University Press, New York 2000, ISBN 0-8014-3734-2.
- Manfred Koch: Wir bestimmen Schmetterlinge. Band 2: Bären, Spinner, Schwärmer und Bohrer Deutschlands. 2., erweiterte Auflage. Neumann, Radebeul/Berlin 1964, DNB 452481929.
- A. R. Pittaway: The Hawkmoths of the western Palaearctic. Harley Books, 1993, ISBN 0-946589-21-6.
- Rolf Reinhardt, Kurt Harz: Wandernde Schwärmerarten. Totenkopf-, Winden-, Oleander- und Linienschwärmer. (= Die Neue Brehm-Bücherei. Band. 596.) Westarp & Spektrum, Magdeburg/Heidelberg/Berlin/Oxford 1996, ISBN 3-89432-859-2.
- Hans-Josef Weidemann, Jochen Köhler: Nachtfalter. Spinner und Schwärmer. Naturbuch-Verlag, Augsburg 1996, ISBN 3-89440-128-1.
Weblinks
- Entomologenportal Fotos
- www.lepiforum.de Fotos und Taxonomie
- Sphingidae of the Western Palaearctic (englisch)
- www.schmetterling-raupe.de
- Moths and Butterflies of Europe and North Africa (englisch)
- Markku Savela: Lepidoptera and some other life forms (englisch)
Einzelnachweise
- Josef J. de Freina, Thomas J. Witt: Noctuoidea, Sphingoidea, Geometroidea, Bombycoidea. In: Die Bombyces und Sphinges der Westpalaearktis. 1. Auflage. Band 1. EFW Edition Forschung & Wissenschaft, München 1987, ISBN 3-926285-00-1, S. 405.
- Rolf Reinhardt, Kurt Harz: Wandernde Schwärmerarten. Totenkopf-, Winden-, Oleander- und Linienschwärmer. In: Die Neue Brehm-Bücherei Bd. 596. 2., unveränderte Auflage. Westarp Wissenschaften, Magdeburg/Heidelberg/Berlin/Oxford 1996, ISBN 3-89432-859-2, S. 30 f.
- A. R. Pittaway: The Hawkmoths of the western Palaearctic. Harley Books, 1993, ISBN 0-946589-21-6, S. 82.
- Rolf Reinhardt, Kurt Harz: Wandernde Schwärmerarten. Totenkopf-, Winden-, Oleander- und Linienschwärmer. In: Die Neue Brehm-Bücherei Bd. 596. 2., unveränderte Auflage. Westarp Wissenschaften, Magdeburg/Heidelberg/Berlin/Oxford 1996, ISBN 3-89432-859-2, S. 32.
- Rolf Reinhardt, Kurt Harz: Wandernde Schwärmerarten. Totenkopf-, Winden-, Oleander- und Linienschwärmer. In: Die Neue Brehm-Bücherei Bd. 596. 2., unveränderte Auflage. Westarp Wissenschaften, Magdeburg/Heidelberg/Berlin/Oxford 1996, ISBN 3-89432-859-2, S. 34.
- Rolf Reinhardt, Kurt Harz: Wandernde Schwärmerarten. Totenkopf-, Winden-, Oleander- und Linienschwärmer. In: Die Neue Brehm-Bücherei Bd. 596. 2., unveränderte Auflage. Westarp Wissenschaften, Magdeburg/Heidelberg/Berlin/Oxford 1996, ISBN 3-89432-859-2, S. 35.
- Rolf Reinhardt, Kurt Harz: Wandernde Schwärmerarten. Totenkopf-, Winden-, Oleander- und Linienschwärmer. In: Die Neue Brehm-Bücherei Bd. 596. 2., unveränderte Auflage. Westarp Wissenschaften, Magdeburg/Heidelberg/Berlin/Oxford 1996, ISBN 3-89432-859-2, S. 36 f.
- Rolf Reinhardt, Kurt Harz: Wandernde Schwärmerarten. Totenkopf-, Winden-, Oleander- und Linienschwärmer. In: Die Neue Brehm-Bücherei Bd. 596. 2., unveränderte Auflage. Westarp Wissenschaften, Magdeburg/Heidelberg/Berlin/Oxford 1996, ISBN 3-89432-859-2, S. 37 f.
- Rolf Reinhardt, Kurt Harz: Wandernde Schwärmerarten. Totenkopf-, Winden-, Oleander- und Linienschwärmer. In: Die Neue Brehm-Bücherei Bd. 596. 2., unveränderte Auflage. Westarp Wissenschaften, Magdeburg/Heidelberg/Berlin/Oxford 1996, ISBN 3-89432-859-2, S. 42 f.
- Sphingidae of the Eastern Palaearctic: Acherontia styx. A.R. Pittaway, abgerufen am 24. September 2009.
- Dubatolov; V. V. (2011): Discovery of Acherontia atropos L. (Lepidoptera, Sphingidae) in North-East Kazakhstan. Amurian zoological journal III(1), S. 58–59
- Sphingidae of the Western Palaearctic: Acherontia atropos. A.R. Pittaway, abgerufen am 4. September 2009.
- Günter Ebert: Die Schmetterlinge Baden Württembergs. 1. Auflage. Band 4. Nachtfalter II Bombycidae, Endromidae, Lasiocampidae, Lemoniidae, Saturniidae, Sphingidae, Drepanidae, Notodontidae, Dilobidae, Lymantriidae, Ctenuchidae, Nolidae. Ulmer, Stuttgart (Hohenheim) 1994, ISBN 3-8001-3474-8, S. 127.
- Josef J. de Freina, Thomas J. Witt: Noctuoidea, Sphingoidea, Geometroidea, Bombycoidea. In: Die Bombyces und Sphinges der Westpalaearktis. 1. Auflage. Band 1. EFW Edition Forschung & Wissenschaft, München 1987, ISBN 3-926285-00-1, S. 406.
- Günter Ebert: Die Schmetterlinge Baden Württembergs. 1. Auflage. Band 4. Nachtfalter II Bombycidae, Endromidae, Lasiocampidae, Lemoniidae, Saturniidae, Sphingidae, Drepanidae, Notodontidae, Dilobidae, Lymantriidae, Ctenuchidae, Nolidae. Ulmer, Stuttgart (Hohenheim) 1994, ISBN 3-8001-3474-8, S. 123.
- Rolf Reinhardt, Kurt Harz: Wandernde Schwärmerarten. Totenkopf-, Winden-, Oleander- und Linienschwärmer. In: Die Neue Brehm-Bücherei Bd. 596. 2., unveränderte Auflage. Westarp Wissenschaften, Magdeburg/Heidelberg/Berlin/Oxford 1996, ISBN 3-89432-859-2, S. 58 ff.
- Toni Schuberl: Exotische Schmetterlinge am Brotjacklriegel. In: Toni Schuberl (Hrsg.): Eginger Jahrbuch 2016. ISBN 978-3-946910-00-8.
- Rolf Reinhardt, Kurt Harz: Wandernde Schwärmerarten. Totenkopf-, Winden-, Oleander- und Linienschwärmer. In: Die Neue Brehm-Bücherei Bd. 596. 2., unveränderte Auflage. Westarp Wissenschaften, Magdeburg/Heidelberg/Berlin/Oxford 1996, ISBN 3-89432-859-2, S. 53 ff.
- Rolf Reinhardt, Kurt Harz: Wandernde Schwärmerarten. Totenkopf-, Winden-, Oleander- und Linienschwärmer. In: Die Neue Brehm-Bücherei Bd. 596. 2., unveränderte Auflage. Westarp Wissenschaften, Magdeburg/Heidelberg/Berlin/Oxford 1996, ISBN 3-89432-859-2, S. 50 ff.
- Rolf Reinhardt, Kurt Harz: Wandernde Schwärmerarten. Totenkopf-, Winden-, Oleander- und Linienschwärmer. In: Die Neue Brehm-Bücherei Bd. 596. 2., unveränderte Auflage. Westarp Wissenschaften, Magdeburg/Heidelberg/Berlin/Oxford 1996, ISBN 3-89432-859-2, S. 56 f.
- R. F. A. Moritz, W. H. Kirchner, R. M. Crewe: Chemical Camouflage of the Death’s Head Hawkmoth (Acherontia atropos L.) in Honeybee Colonies, Naturwissenschaften 78 (1991) 179–182
- Günter Ebert: Die Schmetterlinge Baden Württembergs. 1. Auflage. Band 4. Nachtfalter II Bombycidae, Endromidae, Lasiocampidae, Lemoniidae, Saturniidae, Sphingidae, Drepanidae, Notodontidae, Dilobidae, Lymantriidae, Ctenuchidae, Nolidae. Ulmer, Stuttgart (Hohenheim) 1994, ISBN 3-8001-3474-8, S. 124.
- Rolf Reinhardt, Kurt Harz: Wandernde Schwärmerarten. Totenkopf-, Winden-, Oleander- und Linienschwärmer. In: Die Neue Brehm-Bücherei Bd. 596. 2., unveränderte Auflage. Westarp Wissenschaften, Magdeburg/Heidelberg/Berlin/Oxford 1996, ISBN 3-89432-859-2, S. 39 ff.
- Rolf Reinhardt, Kurt Harz: Wandernde Schwärmerarten. Totenkopf-, Winden-, Oleander- und Linienschwärmer. In: Die Neue Brehm-Bücherei Bd. 596. 2., unveränderte Auflage. Westarp Wissenschaften, Magdeburg/Heidelberg/Berlin/Oxford 1996, ISBN 3-89432-859-2, S. 33.
- Rolf Reinhardt, Kurt Harz: Wandernde Schwärmerarten. Totenkopf-, Winden-, Oleander- und Linienschwärmer. In: Die Neue Brehm-Bücherei Bd. 596. 2., unveränderte Auflage. Westarp Wissenschaften, Magdeburg/Heidelberg/Berlin/Oxford 1996, ISBN 3-89432-859-2, S. 85 ff.
- Hans-Josef Weidemann, Jochen Köhler: Nachtfalter, Spinner und Schwärmer. Naturbuch-Verlag, Augsburg 1996, ISBN 3-89440-128-1, S. 24.
- Acherontia atropos (Linnaeus 1758). Fauna Europaea, Version 1.3, 19.04.2007, abgerufen am 4. September 2009.
- Ian J. Kitching: Phylogeny of the death’s head hawkmoth, Acherontia [Laspeyres], and related genera (Lepidoptera: Sphingidae, Acherontiini), Systematic Entomology 28 (2003) 71-88
- Günter Ebert: Die Schmetterlinge Baden Württembergs. 1. Auflage. Band 4. Nachtfalter II Bombycidae, Endromidae, Lasiocampidae, Lemoniidae, Saturniidae, Sphingidae, Drepanidae, Notodontidae, Dilobidae, Lymantriidae, Ctenuchidae, Nolidae. Ulmer, Stuttgart (Hohenheim) 1994, ISBN 3-8001-3474-8, S. 128.
- Hans-Josef Weidemann, Jochen Köhler: Nachtfalter, Spinner und Schwärmer. Naturbuch-Verlag, Augsburg 1996, ISBN 3-89440-128-1, S. 26.
- Rolf Reinhardt, Kurt Harz: Wandernde Schwärmerarten. Totenkopf-, Winden-, Oleander- und Linienschwärmer. In: Die Neue Brehm-Bücherei Bd. 596. 2., unveränderte Auflage. Westarp Wissenschaften, Magdeburg/Heidelberg/Berlin/Oxford 1996, ISBN 3-89432-859-2, S. 61 f.
- Un Chien Andalou: Frame Enlargements: Part 4 of 4. The University of Alabama, College of Communication and Information Sciences, abgerufen am 29. September 2009.
- James Barbut (1781): The Genera Insectorum of Linnæus, Exemplified by Various Specimens English Insects drawn by Nature