Sonoma-Streifenhörnchen
Das Sonoma-Streifenhörnchen (Tamias sonomae, Syn.: Neotamias sonomae) ist eine Hörnchenart aus der Gattung der Streifenhörnchen (Tamias). Es kommt nur im Nordwesten des amerikanischen Bundesstaats Kalifornien nördlich von San Francisco in zwei Unterarten vor. Es handelt sich um ein mittelgroßes Streifenhörnchen, das wie andere Streifenhörnchen primär in Bäumen und Gebüschen lebt, wo es sich vor allem von verschiedenen Pflanzensamen und Früchten ernährt. Die Art wird einem Verwandtschaftskomplex um das Townsend-Streifenhörnchen (Tamias townsendii) zugeordnet und ist von anderen Arten dieses Komplexes nur sehr schwer zu unterscheiden.
| Sonoma-Streifenhörnchen | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Juveniles (links) und adultes Exemplar (rechts) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Tamias sonomae | ||||||||||||
| (Grinnell, 1915) |
Die Art wird von der International Union for Conservation of Nature and Natural Resources (IUCN) als „nicht gefährdet“ eingestuft und die Bestände werden als stabil betrachtet.
Merkmale
Allgemeine Merkmale
Das Sonoma-Streifenhörnchen erreicht eine durchschnittliche Länge von 22,0 bis 27,7 Zentimetern.[1] Die Kopf-Rumpf-Länge wird für einzelne Individuen mit etwa 13,3 bis 13,9 Zentimetern, die Schwanzlänge mit etwa 10,6 bis 11,6 Zentimetern angegeben, und das Gewicht beträgt etwa 70 Gramm.[2][3] Die Ohrlänge beträgt 15 bis 23 Millimeter und die Hinterfußlänge 33 bis 37 Millimeter.[1][4] Die Tiere weisen keinen Sexualdimorphismus auf und entsprechen sich auch in der Größe, Tiere der Unterart Tamias sonomae alleni sind allerdings etwas kleiner als Individuen der Nominatform.[5]

Die Rückenfarbe der Tiere ist im Sommer, von Juli bis September, rotbraun und wie bei anderen Arten der Gattung befinden sich auf dem Rücken mehrere dunkle Rückenstreifen, die durch hellere Streifen getrennt und gegenüber den Körperseiten abgegrenzt sind.[2] Der Kopf ist rotbraun mit rauchgrauer und zimtfarbener Sprenkelung, die dunklen Gesichtsstreifen sind dunkel-rotbraun bis schwärzlich rotbraun, teilweise zimtfarben durchsetzt. Die Schultern, der vordere Rücken und die beiden mittleren hellen Streifen sind sandfarben bis ocker-sandfarben und die Streifen sind im hinteren Teil weiß durchsetzt. Die äußeren beiden hellen Streifen sind matt-weiß und sandfarben durchsetzt. Die dunklen Rückenstreifen sind schwarz, selten dunkel-rotbraun, und die dunklen Seitenstreifen sind dunkel-rotbraun und am Rand häufig gegenüber der dunkel-sandfarbenen Flankenfärbung nur undeutlich abgegrenzt. Der Rumpf ist ocker-sandfarben und mit einem zimtfarbenen Ton durchsetzt und die Hüften sind rotbraun mit zimtfarbener Tönung und teilweise grauweißer Sprenkelung. Die Füße sind sand- bis zimtfarben, die Hinterfüße teilweise rotbraun eingewaschen. Der Schwanz ist oberseits rotbraun bis schwarz und heller sandbraun eingewaschen, unterseits ist er sandfarben und dunkler eingerahmt. Die Bauchseite ist gräulich-weiß und mit einem blassen ocker-sandfarbenen Ton eingewaschen.[5] Der Fellwechsel zum Winterfell findet im Oktober statt. Die Winterfärbung entspricht der Sommerfärbung, ist jedoch insgesamt dunkler, die Bauchseite ist im Winter cremeweiß. Im Vergleich zur Nominatform ist die Unterart Tamias sonomae alleni etwas kleiner und die Rückenfärbung ist generell etwas dunkler über das gesamte Jahr. Die dunklen Rückenstreifen sind schwärzer und deutlicher ausgeprägt und die hellen Streifen an den Flanken sind mehr sandfarben, im Gesicht besitzt es keine olivgrüne Einfärbung.[5]
Vom Gelben Fichtenstreifenhörnchen unterscheidet sich das Sonoma-Streifenhörnchen vor allem durch die Größe und die rote Färbung. Farblich ähnelt es stark dem Townsend-Streifenhörnchens (Tamias townsendii) sowie den anderen Arten des townsendii-Verwandtschaftskomplexes und kann mit diesen leicht verwechselt werden. Es ist etwas größer mit längeren Gliedmaßen, einem längeren, breiteren und buschigeren Schwanz und längeren Ohren als das Townsend-Hörnchen. Zudem ist es blasser sowie in der Grundfärbung rötlicher und es besitzt eine arttypische weiße Abgrenzung am Schwanz sowie eine rötliche Fläche an der Schwanzunterseite, die im Vergleich zu anderen Arten blasser und nicht im vorderen Bereich dunkler ist. Im Winterfell sind die Wangen grau statt braun, die Ohren sind im Sommerfell nur spärlich behaart und einfarbig.[5]
Merkmale des Schädels und des Skeletts
Der Schädel der Art ist lang und schmal mit einer maximalen Breite, die etwa 54 % der Gesamtlänge ausmacht. Der Schädel der Nominatform hat eine Gesamtlänge von 38,0 bis 39,7, durchschnittlich 38,7 Millimetern, im Bereich der Jochbögen beträgt die Breite 19,4 bis 21,3, durchschnittlich 20,3 Millimeter. Der Schädel von Tamias sonomae alleni ist etwas kleiner. Der Schädel ist im Vergleich zu anderen Arten länger und schmaler mit enger anliegenden Jochbögen. Der Hirnschädel ist lang und abgeflacht,[5] die Schnauzenregion (Rostrum) ist tief und die Nasenbeine sind an der Spitze oberhalb der Schneidezähne durch einen kleinen Knoten getrennt.[5][4] Die Foramina incisiva sind kürzer als bei anderen Arten, das hintere Ende des Gaumenbeins ist verdickt und läuft in einem kurzen statt einem langen Dorn bei anderen Arten aus. Weitere arttypische Merkmale betreffen vor allem den Aufbau und die Ausprägung des Zungenbeins (Os hyoideum).[5]
| 1 | · | 0 | · | 2 | · | 3 | = 22 |
| 1 | · | 0 | · | 1 | · | 3 |
Die Tiere besitzen wie alle der Untergattung Neotamias zugeordneten Arten im Oberkiefer und im Unterkiefer pro Hälfte einen zu einem Nagezahn ausgebildeten Schneidezahn (Incisivus), dem eine Zahnlücke (Diastema) folgt. Hierauf folgen im Oberkiefer je zwei Prämolaren und im Unterkiefer je ein Prämolar sowie drei Molaren. Insgesamt verfügen die Tiere damit über ein Gebiss aus 22 Zähnen.[5] Die oberen Schneidezähne sind im Vergleich zu anderen Arten stark zurückgebogen, die Prämolaren und Molaren sind nur klein ausgebildet.[5]
Wie andere Streifenhörnchen besitzen die männlichen Tiere einen Penisknochen (Bacculum), der eine Länge von etwa 3,0 bis 3,3 Millimetern hat. Er hat einen sehr dünnen Schaft und einen flachen Kiel, der etwa 10 % der Spitze ausmacht. Der Winkel zwischen dem Schaft und der Spitze beträgt 130° und das distale Ende des Schaftes ist leicht seitlich zusammengedrückt.[4][5] Das weibliche Gegenstück, der Klitorisknochen (Baubellum), ist lang und dünn und bildet aus Basis und Schaft eine weite U-Form. Er hat eine Basallänge von 0,5 bis 1,1 Millimetern, die der Länge des Schaftes entspricht. Der Winkel zwischen dem Schaft und der Spitze beträgt 150,4° und die Spitze ist 0,4 bis 0,7 Millimeter lang mit einem sehr kurzen, aber sichtbaren Kiel.[6][5]
Genetische Merkmale
Das Genom des Sonoma-Streifenhörnchens besteht wie das anderer Streifenhörnchen aus 38 Chromosomen (2n=38). Dabei handelt es sich um vier Paare großer metazentrischer, sechs Paare großer submetazentrischer, vier Paare großer akrozentrischer, ein Paar kleiner metazentrischer und drei Paare kleiner akrozentrischer Chromosomen. Das X-Chromosom ist submetazentrisch, das Y-Chromosom akrozentrisch. Innerhalb der Gattung wird es dem Typ-B-Karyotyp zugeordnet.[5][3]
Verbreitung und Lebensraum
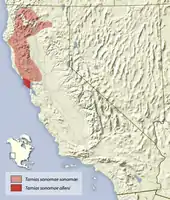
Das Sonoma-Streifenhörnchen kommt nur im Nordwesten des amerikanischen Bundesstaats Kalifornien nördlich von San Francisco vor. Das Verbreitungsgebiet reicht dabei vom Siskiyou County bis an die San Francisco Bay, wobei die Tiere in Höhen unterhalb von 1800 Metern vorkommen.[2][7]
Die Vegetationszonen des Habitats reichen von der oberen Sonora-Zone („upper sonoran zone“), die durch steppenartige Vegetation geprägt ist, bis in die waldreiche Übergangsvegetation („transition zone“).[3] Die Tiere leben im Küstenbereich vor allem in offenen Waldbeständen des Küstenmammutbaums (Sequoia sempervirens) sowie in trockenen Beständen der Gelb-Kiefer (Pinus ponderosa), der Weiß-Kiefer (Pinus sabiniana), der Weihrauchzeder (Calocedrus decurrens), des Amerikanischen Erdbeerbaumes (Arbutus menziesii) und gebüschigen Eichenbeständen (Quercus), etwa im Chaparral. Zudem kommt die Art entlang von Flüssen und offenen Gebüschflächen sowie in Beständen der Kolorado-Tanne (Abies concolor) und der Pracht-Tanne (Abies magnifica) vor.[5][2][3] Jungwälder und Einschlagsgebiete werden von den Tieren in der Regel gemieden.[5]
Das Sonoma-Streifenhörnchen kommt in Teilen seines Verbreitungsgebietes in Nordkalifornien sympatrisch mit dem Gelben Fichtenstreifenhörnchen (Tamias amoenus) sowie dem Gelbwangen-Streifenhörnchen (Tamias ochrogenys), dem Allen-Streifenhörnchen (Tamias senex) und dem Siskiyou-Streifenhörnchen (Tamias siskiyou), die gemeinsam mit ihm der Verwandtschaftsgruppe des townsendii-Komplexes zugeordnet werden, vor. Diese Vorkommen sind vor allem durch das Ineinandergreifen der Höhenzüge und damit der spezifischen Vegetation in den Übergangsgebieten bedingt; während die drei genannten Arten die borealen Fichtenwälder mit Douglasien und Kolorado-Tanne (Abies concolor) bevorzugen, lebt das Sonoma-Streifenhörnchen in von Kolorado-Tanne, Gelb-Kiefer (Pinus ponderosa) und Kalifornischer Schwarzeiche (Quercus kelloggii) geprägten Mischwaldgebieten.[5] Gegenüber dem Merriam-Streifenhörnchen (Tamias merriami) ist das Verbreitungsgebiet über die San Francisco Bay und die San Pablo Bay sowie das dazwischenliegende Grasland abgegrenzt. Vom Verbreitungsgebiet des Langohr-Streifenhörnchens (Tamias quadrimaculatus) in der Sierra Nevada wird es durch die etwa 60 Kilometer breite Region des Lassen Peak getrennt, in der beide Arten nicht vorkommen.[5]
Lebensweise
Die Art ist tagaktiv und lebt sowohl am Boden wie auch kletternd in Bäumen und Gebüschen. Die Waldgebiete, in denen die Art anzutreffen ist, beinhalten Bäume, Sträucher, Holzstapel und eine Auflage aus Pflanzenmaterial am Waldboden. Die Tiere ernähren sich wie andere Streifenhörnchen vor allem herbivor von Samen der Nadelbäume, Eicheln sowie Samen, Früchten und anderen Pflanzenteilen verschiedener Pflanzenarten.[2] Eine wissenschaftliche Untersuchung der Nahrungszusammensetzung liegt nicht vor, es ist jedoch anzunehmen, dass die Tiere verschiedene Pflanzen ihres Lebensraums als Nahrungsquelle nutzen, darunter etwa die Virginische Traubenkirsche (Prunus virginiana, bekannt als chokecherry), Felsenbirnen (Amelanchier) und Becherkätzchen (Garrya). Andere Arten der Gattung ernähren sich zudem von Blättern, Blüten und Kräutern sowie Pilzen, Insekten und Vogeleiern.[4] Die Nahrungssuche erfolgt vor allem im Geäst kleinerer Gebüsche. Die Tiere suchen häufig exponierte Positionen auf Baumstämmen, größeren Ästen der unteren Baumregionen und auf Steinen, als Ruhe- und Beobachtungsplätze und um die gesammelte Nahrung zu verzehren.[5]
Sie kommunizieren über sehr hohe, vogelrufähnliche und arttypische Rufe,[8] die sie bei potenzieller Bedrohung ausstoßen. Die weiblichen Tiere rufen häufiger als die männlichen und es gibt signifikante Unterschiede in der Ruffrequenz von jüngeren und älteren Tieren, tragenden und säugenden Weibchen, paarungsbereiten Männchen und anderen Teilgruppen innerhalb der Populationen.[3] Die Tiere rufen in Form von Rufclustern („bursts“) mit mehreren Einzelrufen („chips“) aus dem Geäst von Bäumen und Gebüschen und auch vom Boden aus, sie können dabei 10 bis über 80 bursts mit 50 bis mehr als 100 chips pro Minute ausstoßen. Die Rufe bilden im Sonagramm eine V-Form, bei dem die Tonhöhe erst abnimmt und später wieder zunimmt; darin unterscheiden sie sich von allen verwandten Arten.[8] Als Reaktion auf die Rufe ziehen sich die Tiere in der Regel rasch tiefer in das Geäst von Bäumen oder Büschen oder in Holzstapel und andere Verstecke zurück, wo sie sich dann ruhig verhalten.[9][5]
Fortpflanzung und Entwicklung

Die Paarungszeit liegt bei dieser Art im Frühjahr und in der Regel haben die Weibchen nur einen Wurf pro Jahr. In tieferen Lagen beginnt die Paarungszeit deutlich früher als in den Hochlagen und die Weibchen der Tieflagen können ihren Eisprung bereits mehrere Wochen vor denen der Hochlagen haben. Bei den Männchen schwellen während der Paarungszeit die Hoden an und wandern vom Dezember bis Juni aus den Lenden in einen äußeren Hodensack; im späten März erreichen die Hoden ihre maximale Länge von 13 bis 16,5 Millimetern. In der Zeit, in der die Männchen fortpflanzungsfähig sind, wandern sie aktiv in die Territorien mit paarungsbereiten Weibchen ein und konkurrieren mit anderen paarungswilligen Männchen.[5]
Auch die Jungtiere werden im Tiefland deutlich früher geboren als in den Hochlagen. Die Tragzeit beträgt etwa 30 bis 31 Tage und die Jungtiere werden im Frühjahr geboren. Der Wurf besteht durchschnittlich aus drei bis fünf Jungtieren und wird 39 bis 45 Tage von der Mutter gesäugt, auch wenn die Jungtiere tagsüber bereits das Nest verlassen. Die Weibchen betreuen den Nachwuchs allein und die weiblichen Jungtiere bleiben nach der Entwöhnung häufig bei der Mutter, während sich die männlichen Tiere verteilen.[2] Man geht davon aus, dass sich durch dieses Verhalten Verwandtschaftsgruppen weiblicher Tiere bilden, die auch die häufigeren Alarmrufe bei den weiblichen Tieren als Ergebnis einer Verwandtenselektion zum Schutz nahe verwandter Individuen bedingt.[3] Das Geschlechterverhältnis der Tiere nach der Geburt und beim Verlassen des Nestes liegt bei etwa 1:1, verändert sich jedoch zu Gunsten der Weibchen bis zum ersten Herbst. Dies wird auf die unterschiedlichen Verteilungsstrategien der männlichen und weiblichen Jungtiere nach dem Verlassen des Nestes zurückgeführt. Auch im ersten Frühjahr nach der Überwinterung überwiegen die jungen Weibchen gegenüber den jungen Männchen, nach der ersten Fortpflanzung liegt das Geschlechterverhältnis im folgenden Herbst jedoch aufgrund der erhöhten Mortalität der Weibchen während ihrer ersten Geburten und Aufzuchten wieder etwa bei 1:1. Im höheren Alter überwiegen erneut die Weibchen, während die Mortalität der Männchen vor allem im Alter von drei bis fünf Jahren deutlich höher liegt als die der Weibchen.[9][5]
Ökologische Bedeutung, Fressfeinde und Parasiten
Der einzige nachgewiesene Prädator des Sonoma-Streifenhörnchens ist der Rotschwanzbussard (Buteo jamaicensis), darüber hinaus stellen jedoch auch andere Greifvögel und Raubtiere potenzielle Fressfeinde dar. Als Parasiten sind zwei Arten von Tierläusen (Hoplopleura arboricola und Neohaematopinus pacificus)[5] sowie die Nymphen verschiedener Zecken (Dermacentor occidentalis, Ixodes pacificus und Ixodes spinipalpis) nachgewiesen.[2] Infektionen mit dem Erreger der Pest, Yersinia pestis, sowie dem Erreger der Lyme-Borreliose, Borrelia burgdorferi, für die Nagetiere der Region als potenzielle Reservoirwirte relevant sind, sind beim Sonoma-Streifenhörnchen nicht nachgewiesen.[10][5] Dagegen konnten Anaplasma phagocytophilum, der ebenfalls über Zecken übertragene Erreger der Humanen Granulozytären Anaplasmose und der Caninen Anaplasmose in mehreren Sonoma-Streifenhörnchen identifiziert werden.[11]
Durch seine Ernährung sowie durch die Anlage von Vorräten spielt das Sonoma-Streifenhörnchen wie andere Arten der Gattung eine wesentliche Rolle bei der Aussaat und Verteilung von Pflanzensamen sowie bei der Verbreitung von Mykorrhiza-Sporen.[4]
Systematik
|
Das Sonoma-Streifenhörnchen wird als eigenständige Art innerhalb der Gattung der Streifenhörnchen (Tamias) eingeordnet, die aus 25 Arten besteht.[14] Die wissenschaftliche Erstbeschreibung stammt von dem amerikanischen Zoologen Joseph Grinnell aus dem Jahr 1915, der es als Eutamias sonomae anhand von Individuen aus der Region westlich von Guerneville, Sonoma County, in Kalifornien beschrieb und nach dem County als Fundort benannte.[15][5][14] Die Tiere stammten dabei aus der Sammlung des California Museum of Vertebrate Zoology in Berkeley,[15] dessen erster Direktor Grinnell war.
1985 wurde die Art im Rahmen einer Revision durch Levenson et al. mit allen anderen Streifenhörnchen der Gattung Tamias zugeschlagen.[16] Innerhalb dieser wird das Sonoma-Streifenhörnchen gemeinsam mit den meisten anderen Arten der Untergattung Neotamias zugeordnet, die auch als eigenständige Gattung diskutiert wird.[17] Teilweise wurde es als Unterart des Townsend-Streifenhörnchens (Tamias townsendii) betrachtet,[14][5] wird heute jedoch einer als townsendii-Gruppe bezeichneten Artengruppe zugeordnet, die neben dem Sonoma- und dem Townsend-Streifenhörnchen sieben weitere Arten enthält: T. senex, T. ochrogenys, T. siskiyou, T. merriami, T. quadrimaculatus, T. obscurus und T. dorsalis. Innerhalb dieser Verwandtschaftsgruppe werden auf der Basis molekularbiologischer Daten das Sonoma-Streifenhörnchen als Schwesterart des Allen-Streifenhörnchens (Tamias senex) und beide gemeinsam als Schwestergruppe des Townsend-Streifenhörnchens betrachtet.[12][13] Alternativ wird das Sonoma-Streifenhörnchen als Schwesterart einer Verwandtschaftsgruppe aus Townsend-Streifenhörnchen, Allen-Streifenhörnchen und Siskiyou-Streifenhörnchen (Tamias siskiyou) angesehen.[18]

Innerhalb der Art werden gemeinsam mit der Nominatform zwei Unterarten unterschieden:[2]
- Tamias sonomae sonomae (Grinnell, 1915): Nominatform; kommt im gesamten Verbreitungsgebiet vor und ist etwas größer und heller als Tamias sonomae alleni.
- Tamias sonomae alleni Howell, 1922: lebt nur im äußersten Südwesten des Verbreitungsgebietes nahe der San Francisco Bay. Die Unterart ist etwas kleiner und dunkler als die Nominatform. Die Form wurde 1922 von Arthur H. Howell als Eutamias townsendii alleni und damit als Unterart des Townsend-Streifenhörnchens anhand eines männlichen Individuums aus Inverness im Marin County, Kalifornien, beschrieben; benannt ist sie nach Joel Asaph Allen. Ursprünglich wurde der Typus von Allen der Unterart Eutamias townsendii hindsii zugeordnet, nachdem diese jedoch mit Eutamias townsendii townsendii synonymisiert wurde, beschrieb Howell auf der Basis des Exemplars die neue Unterart.[19]
Status, Bedrohung und Schutz

Das Sonoma-Streifenhörnchen wird von der International Union for Conservation of Nature and Natural Resources (IUCN) als „nicht gefährdet“ (Least Concern, LC) eingestuft und die Bestände werden als stabil eingeschätzt. Begründet wird dies durch das relativ große Verbreitungsgebiet von mehr als 20.000 km2 und das regelmäßige Vorkommen, bestandsgefährdende Risiken sind nicht bekannt.[7]
Über das Sonoma-Streifenhörnchen und das sehr kleine Verbreitungsareal mit verschiedenen Schlüsselhabitaten liegen nur sehr wenige spezifische Studien vor; daher ist unbekannt, inwieweit anthropogene Einflüsse die Art negativ beeinflussen.[4] Ein potenzielles Risiko besteht aufgrund des sehr kleinen Verbreitungsgebietes, in dem größere Lebensraumveränderungen einen starken Einfluss auf den Gesamtbestand der Art haben könnten.[3]
Belege
- Lloyd G. Ingles: Mammals of the Pacific States: California, Oregon, Washington. Stanford University Press 1965, ISBN 0-8047-1843-1, S. 181.
- Richard W. Thorington Jr., John L. Koprowski, Michael A. Steele: Squirrels of the World. Johns Hopkins University Press, Baltimore MD 2012, ISBN 978-1-4214-0469-1, S. 340–341.
- J.L. Koprowski, E.A. Goldstein, K.R. Bennett, C. Pereira Mendes: Sonoma Chipmunk. In: Don E. Wilson, T.E. Lacher, Jr., Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World: Lagomorphs and Rodents 1. (HMW, Band 6) Lynx Edicions, Barcelona 2016, ISBN 978-84-941892-3-4, S. 789–790.
- Samir Hasmukh Shah: Tamias sonomae (Sonoma chipmunk) im Animal Diversity Web, 1999, abgerufen am 7. Mai 2017.
- Troy L. Best: Tamias sonomae. (Memento vom 15. März 2016 im Internet Archive) Mammalian Species 444, 1993.
- Dallas A. Sutton: The Female Genital Bone of Chipmunks, Genus Eutamias. The Southwestern Naturalist 27 (4), November 1982, S. 393–402. (JSTOR 3670714)
- Neotamias sonomae in der Roten Liste gefährdeter Arten der IUCN 2015.4. Eingestellt von: A.V. Linzey, NatureServe (G. Hammerson), 2008. Abgerufen am 5. August 2016.
- Leonard R. Brand: The vocal repertoire of chipmunks (Genus Eutamias) in California. Animal Behaviour 24 (2), 1976, S. 319–335, doi:10.1016/S0003-3472(76)80040-1.
- Stephen F. Smith: Alarm Calls, Their Origin and Use in Eutamias sonomae. Journal of Mammalogy 59 (4), 1978, S. 888–893, doi:10.2307/1380172. JSTOR 1380172
- Robert S. Lane, Richard N. Brown: Wood rats and kangaroo rats: potential reservoirs of the Lyme disease spirochete in California. Journal of Medical Entomology 28 (3), 1991, S. 299–302, PMID 1875357. (Volltext)
- Daniel Rejmanek, Gideon Bradburd, Janet Foley: Molecular characterization reveals distinct genospecies of Anaplasma phagocytophilum from diverse North American hosts. Journal of Medical Microbiology 61 (2), Februar 2012, S. 204–212, doi:10.1099/jmm.0.034702-0, PMID 1875357. (Volltext)
- Antoinette J. Piaggio, Greg S. Spicer: Molecular Phylogeny of the Chipmunk Genus Tamias Based on the Mitochondrial Cytochrome Oxidase Subunit II Gene. Journal of Mammalian Evolution 7 (3), September 2000, S. 147–166, doi:10.1023/A:1009484302799. (Volltext)
- Antoinette J. Piaggio, Greg S. Spicer: Molecular Phylogeny of the Chipmunks Inferred from Mitochondrial Cytochrome b and Cytochrome Oxidase II Gene Sequences. Molecular Phylogenetics and Evolution 20 (3), September 2001, S. 335–350, doi:10.1006/mpev.2001.097. (Volltext)
- Tamias (Neotamias) sonomae. In: Don E. Wilson, DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4.
- Joseph Grinnell: Eutamias sonomae, a new chipmunk from the inner northern coast belt of California. Contribution from the Museum of vertebrate zoology of the University of California, University of California Press, Berkeley 1915, S. [321]–325. (Volltext)
- Howard Levenson, Robert S. Hoffmann, Charles F. Nadler, Ljerka Deutsch, Scott D. Freeman: Systematics of the Holarctic Chipmunks (Tamias). Journal of Mammalogy 66 (2), Mai 1985, S. 219–242, doi:10.2307/1381236.
- Bruce D. Patterson, Ryan W. Norris: Towards a uniform nomenclature for ground squirrels: the status of the Holarctic chipmunks. Mammalia 80 (3), Mai 2016, S. 241–251, doi:10.1515/mammalia-2015-0004.
- Noah Reid, John R. Demboski, Jack Sullivan: Phylogeny Estimation of the Radiation of Western North American Chipmunks (Tamias) in the Face of Introgression Using Reproductive Protein Genes. Systematic Biology 61 (1), 2012, S. 44–62, doi:10.1093/sysbio/syr094. PMC 3243737 (freier Volltext)
- Arthur H. Howell: Diagnoses of Seven New Chipmunks of the Genus Eutamias, with a List of the American Species. Journal of Mammalogy 3, 1922, S. 178–185. (Volltext)
Literatur
- Richard W. Thorington Jr., John L. Koprowski, Michael A. Steele: Squirrels of the World. Johns Hopkins University Press, Baltimore MD 2012, ISBN 978-1-4214-0469-1, S. 340–341.
- Troy L. Best: Tamias sonomae. Mammalian Species 444, 1993.
- J.L. Koprowski, E.A. Goldstein, K.R. Bennett, C. Pereira Mendes: Sonoma Chipmunk. In: Don E. Wilson, T.E. Lacher, Jr., Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World: Lagomorphs and Rodents 1. (HMW, Band 6) Lynx Edicions, Barcelona 2016, S. 789–790, ISBN 978-84-941892-3-4.
Weblinks
- Neotamias sonomae in der Roten Liste gefährdeter Arten der IUCN 2015.4. Eingestellt von: A.V. Linzey, NatureServe (G. Hammerson), 2008. Abgerufen am 5. August 2016.
- Samir Hasmukh Shah: Tamias sonomae (Sonoma chipmunk) im Animal Diversity Web, 1999, abgerufen am 7. Mai 2017.