Mochlodon
Mochlodon ist eine Gattung von kleinwüchsigen Vogelbeckensauriern aus der Gruppe der Rhabdodontidae innerhalb der Iguanodontia. Die Gattung ist mit zwei Arten aus der Oberkreide von Österreich und Ungarn bekannt.
| Mochlodon | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Mochlodon suessi; Lectotypus PIUW 2349/2 | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberkreide (Santonium – Unteres Campanium) | ||||||||||||
| 86,3 bis 76 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Mochlodon | ||||||||||||
| Seeley, 1881 | ||||||||||||
| Arten | ||||||||||||
Etymologie und Forschungsgeschichte
Der Gattungsname Mochlodon leitet sich ab von den altgriechischen Begriffen Μοχλός („mokhlos“: „Hebel“, „Stange“, „Pfahl“) und ὀδών („odon“: „Zahn“).[1]
Im Jahr 1859 durchsuchten Eduard Suess und sein damaliger Schüler Ferdinand Stoliczka das Haldenmaterial des Kohlebergwerks bei Felbring (Muthmannsdorf) in Niederösterreich nach kreidezeitlichen Süßwassermollusken. Dabei gelang Stoliczka der Fund eines kleinen Zahnes, der in seiner Form stark an jene der bereits 1824 durch Gideon Mantell beschriebenen Dinosauriergattung Iguanodon erinnerte.[2]
Versuche des Bergverwalters Pawlowitsch den Fundhorizont genauer einzugrenzen und weitere Wirbeltierfossilien zu bergen, blieben zunächst erfolglos.[2] Erst beim Vortrieb des neuen „Konstantinstollens“ wurde die fossilführende Schicht erneut aufgefahren und man konnte umfangreiches Belegmaterial sammeln, das an das Paläontologische Institut der Universität Wien übergeben wurde.[2][3][4] Emanuel Bunzel wurde mit einer ersten Sichtung und Beschreibung der Funde beauftragt. Seine Bemühungen wurden 1871 veröffentlicht und resultierten, unter anderem, in der Erstbeschreibung der Gattung Struthiosaurus. Die „Iguanodon-ähnlichen“ Funde aus dieser Sammlung beschrieb Bunzel auf Basis eines Unterkieferfragments (PIUW 2349/2), eines Einzelzahnes und zweier Wirbelkörper, zu Ehren von Suess, zunächst als Iguanodon suessii.[2]
Im Frühjahr 1879 lud Suess den britischen Paläontologen Harry Govier Seeley ein, nach Wien zu kommen und die Funde von Muthmannsdorf genauer zu untersuchen. Seeley verbrachte etwa einen Monat in Wien und gelangte insbesondere in Bezug auf Iguanodon suessii zu wesentlich anderen Schlussfolgerungen als Bunzel. Zum einen sah er im Unterkieferfragment (PIUW 2349/2) ausreichend Unterschiede zur Gattung Iguanodon um die Etablierung einer neuen Gattung Mochlodon zu rechtfertigen, zum anderen interpretierte er weitere Fundstücke, die Bunzel diversen anderen Reptiliengruppen zugeordnet hatte, als vermutlich zu Mochlodon suessii gehörig.[5] Bemerkenswerterweise stellte Seeley in seiner 1881 veröffentlichten Neubeschreibung ausschließlich Vergleiche mit der Gattung Iguanodon an, nicht jedoch mit der bereits 1869 von Philippe Matheron beschriebenen Gattung Rhabdodon aus der Kreide Frankreichs.[4][5]

Zu Beginn des 20. Jahrhunderts beschrieb Franz von Nopcsa in einer Serie von Publikationen ähnliche Funde aus dem Gebiet um Hațeg im heutigen Rumänien, die er zunächst als Mochlodon suessi beziehungsweise als Mochlodon (?) robustum bezeichnete.[6] 1902 revidierte Nopcsa seine Meinung und interpretierte Mochlodon suessi als Jungtiere von Mochlodon robustum, wodurch letzteres Taxon als Juniorsynonym von Mochlodon suessi obsolet wurde.[7] Nach einem direkten Vergleich mit dem Belegmaterial von Rhabdodon priscus, das Matheron bereits 1869 beschrieben, aber nicht akkurat abgebildet hatte, kam Nopcsa 1915 zu der Schlussfolgerung, dass Rhabdodon priscus und Mochlodon suessi ident und die morphologischen Unterschiede auf einen Sexualdimorphismus zurückzuführen seien. Konsequenterweise unterschied Nopcsa in dieser Arbeit nur noch zwischen Rhabdodon priscus und Rhabdodon priscus var. suessi, die er als unterschiedliche Geschlechter ein und derselben Art interpretierte.[8] 1923 versuchte Nopcsa seine Hypothese zu untermauern, synonymisierte Mochlodon mit Rhabdodon priscum und wies dem französischen Belegmaterial sowie den ursprünglich als Mochlodon robustum bezeichneten Formen Rumäniens ein weibliches Geschlecht zu, während er die zuvor als Mochlodon suessi beschriebenen Formen als männliche Individuen derselben Art interpretierte.[9] 1929 behauptete er hingegen das genaue Gegenteil und bezeichnete Rhabdodon priscum (= Rhabdodon robustum) als Männchen und Rhabdodon suessi als Weibchen derselben Art.[10]
Nach dem Tod Nopcsas im Jahr 1933 kam die Grundlagenforschung zu diesem Themenkreis praktisch vollständig zum Erliegen. Die Bedeutung der Funde war jedoch allgemein anerkannt und wurde in zahlreichen Fachpublikationen und Lehrbüchern rezipiert. Viele dieser Autoren verwendeten jedoch weiterhin die Gattungsbezeichnung Mochlodon, was daran lag, dass der Name Rhabdodon in diesem Zusammenhang streng genommen ungültig war. Bereits 1831 hatte Friedrich Ludwig Fleischmann eine angeblich neue Schlangenart als Rhabdodon fuscus beschrieben. Zwar konnte die von Fleischmann beschriebene und abgebildete Schlange später unschwer als Vertreter der Eidechsennattern (Malpolon) identifiziert werden, ein Taxon, das Leopold Fitzinger schon 1826 eingeführt hatte, die Bezeichnung Rhabdodon war jedoch 1869, als Matheron die französischen Fossilfunde beschrieb, eindeutig bereits belegt und nicht verfügbar.[11]
Um das Problem zu lösen, beantragte Winand Brinkmann 1986 bei der ICZN das Taxon Rhabdodon Fleischmann, 1831 aus der offiziellen Liste der zoologischen Gattungsnamen zu streichen und stattdessen das Taxon Rhabdodon Matheron, 1869 in die Liste aufzunehmen („Case 2536“).[11] Der Antrag kam 1987 bei der ICZN zur Abstimmung und wurde mit großer Mehrheit befürwortet („Opinion 1483“). Das Ergebnis der Abstimmung wurde 1988 veröffentlicht und damit waren alle entsprechenden Funde aus Frankreich, Österreich und Rumänien der Gattung Rhabdodon zuzuordnen.[12]
2003 gliederten David B. Weishampel und Koautoren die rumänischen Funde als eigenständige Gattung Zalmoxes mit zwei Arten (Zalmoxes robustus und Zalmoxes shqiperorum) wieder aus der Gattung Rhabdodon aus und führten für beide Gattungen das übergeordnete Taxon der Rhabdodontidae ein. Die Funde aus Niederösterreich wurden in dieser Publikation erneut unter der Bezeichnung Mochlodon suessi geführt. Die Autoren hielten es für möglich, dass sie eine weitere eigenständige Gattung innerhalb der Rhabdodontidae repräsentierten, ließen die Frage jedoch mit dem Hinweis auf eine gerade in Arbeit befindliche Neubewertung des entsprechenden Belegmaterials vorläufig noch offen.[13]
Die von Weishampel et al. 2003 erwähnte Neubearbeitung wurde erst 2006 veröffentlicht. Die Autoren dieser Studie kamen zu dem Schluss, dass das Belegmaterial nicht für eine genaue Diagnose ausreichte und interpretierten die niederösterreichischen Funde als Jungtier einer nicht näher bestimmbaren Art der Gattung Zalmoxes (Zalmoxes sp.). Gleichzeitig wurde darauf hingewiesen, dass die beiden von Bunzel in seiner Erstbeschreibung erwähnten Wirbelkörper nicht mehr auffindbar waren.[14]
2012 ergab sich eine drastische Wendung in der Angelegenheit. Attila Ősi und mehrere Koautoren, darunter auch Weishampel, berichteten von umfangreichem neuem Belegmaterial aus der Csehbánya-Formation des Iharkút-Bauxit-Tagebaus im Komitat Veszprém in Ungarn, das in Bezug auf anatomische Details und Größe weitgehende Übereinstimmung mit den Funden von Muthmannsdorf aufwies. Die Gattung Mochlodon wurde mit Mochlodon suessi als Typusart als gültiges eigenständiges Taxon wieder eingeführt und um eine zweite Art Mochlodon vorosi von der ungarischen Fundstelle ergänzt. Der Artzusatz „vorosi“ ehrt den ungarischen Paläontologen Attila Vörös.[4]
Fossilbeleg und Alterseinstufung der Funde
_(16578115769).jpg.webp)
Mochlodon suessi (Typusart)
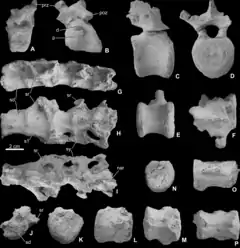
Fossilfunde dieser Art sind ausschließlich von der historischen Fundstelle bei Muthmannsdorf bekannt. Nach Ősi et al., 2012[4] umfasst das Belegmaterial:
- Das Fragment eines rechten Unterkiefers mit mehreren Zähnen (PIUW 2349/2; Lectotypus für Typusart und Gattung)
- Einen einzelnen, isolierten Zahn des Unterkiefers (PIUW 2349/3)
- Einen einzelnen, isolierten Zahn des Oberkiefers (PIUW 2349/4)
- Das Fragment eines Scheitelbeins (PIUW 2349/54; von Bunzel ursprünglich als Scheitelbein einer Eidechse identifiziert)
- Ein unvollständiges, linkes Schulterblatt (PIUW 3518)
- Ein fragliches Speichenfragment (PIUW 3517)
- Ein einzelnes, isoliertes Krallenbein, vermutlich von einer der vorderen Gliedmaßen (PIUW 2349/38)
- Das Fragment eines linken Oberschenkelknochens (PIUW 2349/III)
- Das Fragment eines vermutlich rechten Schienbeins (PIUW 2348/35)
Alle Belegstücke stammen aus der Grünbach-Formation als Teil der Gosau-Gruppe und werden am Paläontologischen Institut der Universität Wien (PIUW) aufbewahrt.[4] Die kohleführende Grünbach-Formation lässt sich auf Basis von biostratigraphisch verwertbaren Fossilfunden aus der unterlagernden Maiersdorf-Formation und der überlagernden Piesting-Formation dem unteren Campanium zuordnen, was einem Alter von etwa 83,5–76 Ma entspricht.[15]
Mochlodon vorosi
Von der zweiten Art liegt wesentlich mehr Belegmaterial vor:[4]
- Ein vollständiger linker Unterkiefer mit vier abgebrochenen Zähnen (MTM V 2010.105.1; Holotypus der Art)
- Vier weitere, fast vollständige Unterkiefer, zwei linke (MTM V 2010.105.1, MTM 2012.15.1) und zwei rechte (MTM V 2010.107.1, MTM V 2010.109.1), sowie insgesamt sechs Unterkieferfragmente
- Ein linkes Postorbitale (MTM 2012.14.1)
- Zwei rechte Quadrata (MTM V 2010.110.1, MTM V 2010.111.1)
- Insgesamt 15 einzelne Zähne des Oberkiefers und 23 einzelne Zähne des Unterkiefers
- Vier einzelne Wirbel aus unterschiedlichen Abschnitten der Wirbelsäule
- Ein fast vollständiges, aber flachgedrücktes Kreuzbein (MTM V 2010.121.1)
- Insgesamt drei Rabenbeine (MTM V 01.53, MTM V 2010.122.1, MTM V 2010.123.1) und ein fragmentarisches Schulterblatt (MTM 2012.22.1)
- Ein vollständiger Oberarmknochen (MTM V 2010.128.1), sowie das Fragment eines weiteren (MTM 2012.23.1)
- Eine vollständige Ulna (MTM 2012.24.1)
- Zwei fast vollständige (MTM V 01.225, MTM V 2010.126.1) und ein fragmentarisch erhaltener Oberschenkelknochen (MTM 2012.25.1)
- Ein vollständiger Schienbeinknochen (MTM V 2010.127.1) und zwei Schienbeinfragmente (MTM V 01.101, MTM 2012.26.1)
- Zwei Phalangen (MTM 2012.27.1, MTM 2012.28.1)
Alle Fossilbelege stammen aus der Csehbánya-Formation und werden am Ungarischen Naturwissenschaftlichen Museum (Magyar Természettudományi Múzeum – MTM) in Budapest aufbewahrt.[4] Die Csehbánya-Formation lässt sich auf Basis palynologischer Befunde in das Santonium (etwa 86,3–83,5 mya) stellen.[16] Mochlodon vorosi ist damit nicht nur älter als Mochlodon suessi, sondern gleichzeitig auch der älteste bekannte Vertreter der sich gesichert den Rhabdodontidae zuordnen lässt.[4]
Merkmale
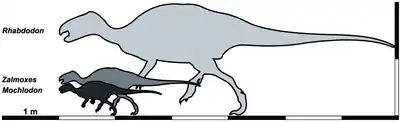
Körpergröße und Körpermasse
Mochlodon ist eine Gattung relativ kleinwüchsiger Vertreter der Iguanodontia. Auf Basis der vorhandenen Fossilbelege lässt sich die Gesamtkörperlänge der adulten Tiere auf etwa 1,6–1,8 m bestimmen. Mochlodon war damit deutlich kleiner als die nahe verwandte Gattung Zalmoxes, für die eine maximale Gesamtkörperlänge von bis zu 2,5 m wahrscheinlich ist. Die Gattungen Mochlodon und Zalmoxes aus Österreich, Ungarn und Rumänien stehen damit größenmäßig in deutlichem Gegensatz zur Gattung Rhabdodon aus Frankreich und Spanien, die eine Gesamtkörperlänge von bis zu 6 m erreichen konnte.[4]
Für Mochlodon vorosi wird eine Körpermasse von 31 kg geschätzt.[17]
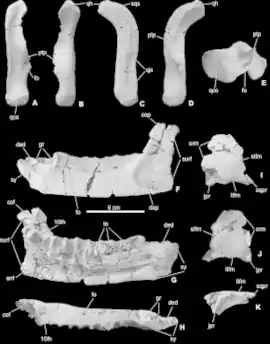
Gattungs- und arttypische Merkmale
(In Klammer gesetzte Abkürzungen beziehen sich auf die nebenstehende Abbildung. Die Beschreibung der gattungstypischen Merkmale erfolgt in Anlehnung an die Beschreibung durch Ősi et al., 2012.[4])
Die Gattung Mochlodon unterscheidet sich von anderen Vertretern der Rhabdodontidae insbesondere durch einige Merkmale des bezahnten Unterkiefers (Dentale):
Die Außenseite des Unterkiefers weist caudal, unmittelbar unterhalb des Processus coronoideus („cop“) eine flache Vertiefung („dep“) auf. Dieses Merkmal ist bei Mochlodon suessi schwächer ausgeprägt als bei Mochlodon vorosi. Das umfangreichere Belegmaterial für Mochlodon vorosi deutet zudem darauf hin, dass das Merkmal bei größeren Individuen stärker ausgeprägt war, als bei kleineren. Die Oberfläche dieser Vertiefung ist mit feinen, rostrocaudal orientierten Knochenleisten skulpturiert. Die physiologische Funktion dieser Vertiefung ist noch nicht abschließend geklärt, vermutet wird jedoch, dass es sich um eine verstärkte Ansatzstelle für die externen Kieferadduktoren handelt.
Die Symphyse („sy“), die Kontaktfläche der knorpeligen Verbindung zwischen den beiden Unterkieferästen, ist nur leicht nach innen gekrümmt. Die dorsale Kante („ded“) der Symphyse verläuft in seitlicher Ansicht gerade, parallel zur Längsachse des Unterkiefers (Mochlodon vorosi) oder ist nur leicht rostroventral geneigt (Mochlodon suessi). In diesem Bereich befindet sich zudem an der Oberkante des Unterkiefers eine tiefe Grube („gr“), die sich caudal erweitert.
Mochlodon vorosi unterscheidet sich zudem von Rhabdodon und Zalmoxes in der Form des Quadratbeins, dessen Kopf („qh“) stark nach hinten gekrümmt ist. Bei Mochlodon suessi liegt kein entsprechender Fossilbeleg für dieses Element des primären Kiefergelenks vor.
Bezahnung

(In Klammer gesetzte Abkürzungen beziehen sich auf die nebenstehende Abbildung. Die Beschreibung der Bezahnung erfolgt in Anlehnung an die Beschreibungen durch Ősi et al., 2012[4] und Virág & Ősi, 2017.[18])
Wie bei anderen Vertretern der Rhabdodontidae unterscheiden sich die Zähne des Ober- und des Unterkiefers in ihrer Form:
Die Krone der Oberkieferzähne weist in labialer und lingualer Ansicht einen annähernd abgerundet rechteckigen Umriss auf. Die Abnutzungsfläche („wf“) liegt lingual. Der Zahnschmelz („en“) der Labialseite ist mit 8–13 annähernd parallelen und gleich starken Längsrippen („lr“) ornamentiert, die an der Basis der Krone sowie mesial und distal von einem leicht erhöhten Rand („cr“) umrahmt werden. Der mesiale und der distale Rand der Krone sind leicht gezähnt. Bei weniger stark abgenutzten Oberkieferzähnen zeigt der Schmelz der Lingualseite ebenfalls eine Ornamentierung mit schwach ausgeprägten, parallelen Längsrippen, die jedoch bei zunehmender Abnutzung verschwindet.
Die Krone der Unterkieferzähne weist in labialer und lingualer Ansicht einen annähernd rautenförmigen Umriss auf. Die Abnutzungsfläche liegt labial. Der Zahnschmelz der lingualen Seite wird, im Gegensatz zu den Zähnen des Oberkiefers, durch eine besonders stark ausgeprägte, mittig liegende, primäre Längsrippe („pr“) in zwei U-förmige Felder geteilt. Die primäre Längsrippe wird auf jedem dieser Felder von 5–7 schwächer ausgeprägten Nebenrippen („sr“) begleitet, die in der Regel nicht bis zur Basis der Krone reichen. Der mesiale und der distale Rand der Krone sind leicht gezähnt („dm“). Ein leicht erhöhter Rand an der Kronenbasis fehlt. Die Labialseite weniger stark abgenutzter Unterkieferzähne zeigt, analog zur Lingualseite der Oberkieferzähne, ebenfalls eine Ornamentierung mit schwach ausgeprägten, parallelen Längsrippen, die bei zunehmender Abnutzung verschwindet.
Die einfachen Wurzeln der Oberkieferzähne sind robust und erreichen eine Länge, die etwa dem Dreifachen der Kronenlänge entspricht, während die Wurzeln der Unterkieferzähne maximal nur etwa 1,5-fache Kronenlänge erreichen. In beiden Fällen finden sich auf der Lingualseite der Wurzeln flache, elliptische Einbuchtungen („rgr“), die der Position der Krone des jeweils nachdrängenden Ersatzzahnes entsprechen.
Die Abnutzungsflächen der Zähne zeigen gerade, annähernd parallele Kratzer, die auf eine weitgehend orthale Kaubewegung der Kiefer schließen lassen. Das bedeutet, dass sich Ober- und Unterkiefer nur vertikal, nicht jedoch palinal (vor und zurück) oder transversal (seitlich) zueinander bewegten. Einige wenige Einzelzähne, die ursprünglich als Unterkieferzähne von Mochlodon vorosi bestimmt worden waren, zeigen ein davon abweichendes Abnutzungsmuster mit annähernd konzentrisch angeordneten, bogenförmigen Kratzern, die für eine gleichzeitige Kombination aus orthaler und palinaler Kieferbewegung beim Kauvorgang sprechen. Eine solche circumpalinale Kaubewegung ist von Vertretern der Ornithopoda unbekannt, wurde jedoch bei einigen Vertretern der Neoceratopsia festgestellt, deren Zähne große Ähnlichkeit mit denen von Ornithopoden haben können.[19] Es wird daher vermutet, dass die Einzelzähne mit abweichenden Abnutzungsmustern nicht Mochlodon vorosi zuzuordnen sind, sondern Ajkaceratops kozmai, dem einzigen bekannten Vertreter der Neoceratopsida aus der Csehbánya-Formation.[18]
Systematik
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Systematische Stellung der Gattung Mochlodon innerhalb der Ornithopoda, vereinfacht nach Herne et al., 2019.[20] |
Das nebenstehende Kladogramm zeigt die systematische Stellung von Mochlodon innerhalb der Ornithopoda, vereinfacht nach Matthew C. Herne et al., 2019.[20]
Mochlodon zeigt sich in dieser phylogenetischen Analyse als Schwestertaxon von Zalmoxes. Die beiden kleinwüchsigen Gattungen Osteuropas bilden innerhalb der Rhabdodontidae eine eigene Teilklade, der die größere, westeuropäische Gattung Rhabdodon gegenübersteht. Dem Alter von Mochlodon vorosi entsprechend muss die Trennung in kleinwüchsige, osteuropäische und große, westeuropäische Formen bereits vor dem Santonium erfolgt sein.[4]
Die Rhabdodontidae selbst bilden eine eigenständige Teilklade innerhalb der Gruppe der Iguanodontia, die allen anderen Vertretern dieser Gruppe gegenübersteht.[20] Sie entsprechen rangmäßig in etwa einer Familie und sind für Europa endemisch.[4]
Weishampel et al. hatten die Rhabdodontidae ursprünglich definiert als „der jüngste gemeinsame Vorfahr von Zalmoxes robustus und Rhabdodon priscus und alle seine Nachfahren“ (knotenbasiertes Taxon).[13] Diese Definition wurde später allerdings im Sinne der allgemeinen Vorschläge von Paul Sereno[21] umformuliert zu „alle Vertreter der Iguanodontia die mit Rhabdodon priscus näher verwandt sind als mit Parasaurolophus walkeri“[22] oder, gleichbedeutend, aber anders ausgedrückt: „die umfassendste Klade, die Rhabdodon priscus enthält, nicht jedoch Parasaurolophus walkeri“[23] (stammbasiertes Taxon).
2016 identifizierten Paul-Emile Dieudonné und Koautoren die australische Gattung Muttaburrasaurus als Schwestertaxon der Rhabdodontidae und führten das übergeordnete Taxon der Rhabdodontomorpha (Rhabdodontidae + Muttaburrasaurus; knotenbasiertes Taxon) ein.[23] Dieser Interpretation widersprachen 2019 jedoch Herne et al., die bei ihrer Analyse Muttaburrasaurus in einer gemeinsamen Klade mit den Dryomorpha, also abseits der Rhabdodontidae, fanden.[20]
Palökologie
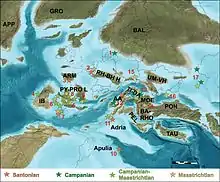
Mitteleuropa bestand während der oberen Kreidezeit aus einer Ansammlung von größeren und kleineren, weitgehend voneinander isolierten Inseln („Europäischer Kreide-Archipel“).[24]
Die Sedimente der kohleführenden Grünbach-Formation wurden am Küstensaum und den angrenzenden, weiter im Landesinneren liegenden, Gebieten einer dieser Inseln abgelagert. Zahlreiche Pflanzenfossilien bieten Einblick in das komplexe Ökosystem. Teiche und Altgewässer waren zumindest teilweise mit Wasserpflanzen der Gattung Quereuxia und Brasenites überwuchert. Die Flora der angrenzenden Sumpfgebiete wurde von Farnen und Schraubenbaumgewächsen (Pandanites) dominiert. Sumpf-, Bruch- und Auwälder wurden von Vertretern der Walnussgewächse, Palmen (Sabalites) und immergrünen Nadelbäumen der Gattung Geinitzia aufgebaut. In den Uferregionen von Bächen und Flüssen wuchsen Farne der Gattung Sphenopteris und Bäume und Sträucher aus der Familie der Platanengewächse, wobei letztere mit Ettingshausenia das dominierende Florenelement dieses Standorts bildeten. Mesophytische Wälder an etwas höher gelegenen und trockeneren Standorten wurden von Koniferen der Gattung Pagiophyllum dominiert. Die Paläoflora der Grünbach-Formation lässt auf ein humides, subtropisches Klima mit heißen Sommern und relativ kurzen, nicht ariden Trockenzeiten und frostfreien Wintern schließen.[25]
Die alluvialen Sedimente der etwas älteren Csehbánya-Formation wurden im Tiefland-Saum einer der größeren dieser Inseln in einer Schwemmebene abgelagert.[26] Weiter zur Küste hin gingen die Schwemmebenen in die Süßwasser-Sümpfe und Seen der Ajka Kohle-Formation über.[27] Das Pollenspektrum der Csehbánya-Formation wird von Vertretern der Normapolles-Gruppe dominiert (40 %), die vermutlich von Pflanzen aus der Verwandtschaft der Buchenartigen stammen. Pollen von anderen Angiospermen tragen weitere 15 % bei und Sporen von Moosen und Farnen sind gemeinsam mit 37 % vertreten, während Pollen von Gymnospermen mit 8 % nur eine untergeordnete Rolle spielen.[16] Die Schwemmebene der Csehbánya-Formation war vermutlich mit Normapolles-dominierten Laubmischwäldern bewachsen. Häufig treten Nussfrüchte der Gattung Sphaeracostata auf, die dieser Gruppe zugeordnet werden. Magnoliengewächse der Gattung Padragkutia und Nadelbäume aus der Familie der Araukariengewächse stellten untergeordnete Elemente dieser Wälder, wobei bei Letzteren auch die Möglichkeit besteht, dass ihre Überreste aus höher gelegenen Bereichen des Hinterlandes in den Ablagerungsraum gespült wurden. Das Unterholz wurde von Farnen und krautigen Bedecktsamigen Pflanzen gebildet. Lokale Anhäufungen von Brennnesselgewächsen werden als Anzeichen für einen mäßig durchfeuchteten Untergrund und häufige Störungen des Ökosystems gewertet. Die Uferbereiche von seichten Seen und Teichen wurden durch Bestände von Pandanites geprägt. In Summe lässt die Paläoflora der Csehbánya-Formation ebenfalls auf ein humides, subtropisches Klima mit jahreszeitlich bedingten Trockenperioden schließen.[26]
Mochlodon war, wie alle Vertreter der Rhabdodontidae, vermutlich ein reiner Pflanzenfresser. Die scherenartig schließende Bezahnung war gut geeignet um auch relativ hartfaserige Pflanzenteile zu zerschneiden und einige Autoren vermuten, dass die Rhabdodontidae allgemein auf entsprechende Einkeimblättrige Pflanzen wie Sabalites oder Pandanites spezialisiert waren.[22]
Bereits Franz von Nopcsa hatte zu Beginn des 20. Jahrhunderts Überlegungen dazu angestellt, ob es sich bei den kleinwüchsigen, oberkretazischen Ornithopoden aus Österreich und Rumänien nicht um einen Fall von Inselverzwergung handeln könnte. Auf Basis der Funde von Mochlodon vorosi stellen Ősi et al., 2012 allerdings die alternative Hypothese in den Raum, dass die Gattungen Mochlodon und Zalmoxes keine Fälle von Inselverzwergung darstellen, sondern, ganz im Gegenteil die großwüchsige Gattung Rhabdodon einen Fall von Inselgigantismus darstellt.[4]
Einzelnachweise
- A. Gaudry: Les enchaînements du monde animal dans les temps géologiques: Fossiles secondaires. Band 3, Paris, 1890, S. 222 (Digitalisat).
- E. Bunzel: Die Reptilfauna der Gosauformation in der Neuen Welt bei Wiener-Neustadt. In: Abhandlungen der k. & k. geologischen Reichsanstalt, Band 5, Nummer 1, 1871, 18 S. + 8 Tafeln (zobodat.at [PDF]).
- B. Plöchinger, G. Bardossy, R. Oberhauser & A. Papp: Die Gosaumulde von Grünbach und der Neuen Welt (Niederösterreich). In: Jahrbuch der Geologischen Bundesanstalt, Band 104, 1961, S. 359–441 (Digitalisat).
- A. Ősi, E. Prondvai, R. Butler & D. B. Weishampel: Phylogeny, Histology and Inferred Body Size Evolution in a New Rhabdodontid Dinosaur from the Late Cretaceous of Hungary. In: PLoS ONE, Band 7, Nummer 9, 2012, e44318, doi:10.1371/journal.pone.0044318.
- H. G. Seeley: The Reptile Fauna of the Gosau Formation preserved in the Geological Museum of the University of Vienna. With a Note on the Geological Horizon of the Fossils at Neue Welt, west of Wiener Neustadt. In: The Quarterly Journal of the Geological Society of London, Band 37, 1881, S. 620–707 (Digitalisat).
- F. Nopcsa: Dinosaurierreste aus Siebenbürgen (Schädel von Limnosaurus transsylvanicus nov. gen. et spec.). In: Denkschriften der kaiserlichen Akademie der Wissenschaften Wien, Mathematisch-Naturwissenschaftliche Classe, Band 68, 1900, S. 555–591 (Digitalisat).
- F. Nopcsa: Dinosaurierreste aus Siebenbürgen II (Schädelreste von Mochlodon) mit einem Anhange: zur Phylogenie der Ornithopodiden. In: Denkschriften der kaiserlichen Akademie der Wissenschaften Wien, Mathematisch-Naturwissenschaftliche Classe, Band 72, 1902, S. 149–175 (zobodat.at [PDF]).
- F. Nopcsa: Die Dinosaurier der siebenbürgischen Landesteile Ungarns. In: Mitteilungen aus dem Jahrbuche der königlich ungarischen Geologischen Reichsanstalt, Band 23, Nummer 1, 1915, S. 1–24 (Digitalisat).
- F. Nopcsa: On the Geological Importance of the Primitive Reptilian Fauna of the Uppermost Cretaceous of Hungary; with a Description of a New Tortoise (Kallokibotium). In: Quarterly Journal of the Geological Society of London, Band 79, 1923, S. 100–116 (Digitalisat).
- F. Nopcsa: Sexual Differences in Ornithopodous Dinosaurs. In: Palaeobiologica, Band 2, 1929, S. 187–200 (zobodat.at [PDF]).
- W. Brinkmann: Rhabdodon Matheron, 1869 (Reptilia, Ornithischia): proposed conservation by suppression of Rhabdodon Fleishmann, 1831 (Reptilia, Serpentes) - Z.N.(S.) 2536. In: The Bulletin of Zoological Nomenclature, Band 43, Nummer 3, 1986, S. 269–272 (Digitalisat).
- ICZN: Opinion 1483 − Rhabdodon Matheron, 1869 (Reptilia, Ornithischia): conserved. In: The Bulletin of Zoological Nomenclature, Band 45, Nummer 1, 1988, S. 85–86 (Digitalisat).
- D. B. Weishampel, C.-M. Jianu, Z. Csiki & D. B. Norman: Osteology and phylogeny of Zalmoxes (n. g.), an unusual Euornithopod dinosaur from the latest Cretaceous of Romania. In: Journal of Systematic Palaentology, Band 1, Nummer 2, 2003, S. 65–123 (Digitalisat).
- S. Sachs & J. J. Hornung: Juvenile ornithopod (Dinosauria: Rhabdodontidae) remains from the Upper Cretaceous (Lower Campanian, Gosau Group) of Muthmannsdorf (Lower Austria). In: Geobios, Band 39, Nummer 3, 2006, S. 415–425 (Zusammenfassung).
- Gerald Hofer, Erich Draganits, Michael Wagreich, Christa-Charlotte Hofmann, Doris Reischenbacher, Marie-Louise Grundtner & Magda Bottig: Stratigraphy and Geochemical Characterisation of Upper Cretaceous Nonmarine – Marine Cycles (Grünbach Formation, Gosau Group, Austria). In: Austrian Journal of Earth Sciences, Band 104, Nummer 2, 2011, S. 90–107 (Digitalisat)
- E. R. Bodor & V. Baranyi: Palynomorphs of the Normapolles group and related plant mesofossils from the Iharkút vertebrate site, Bakony Mountains (Hungary). In: Central European Geology, Band 55, Nummer 3, 2012, S. 259–292 (Digitalisat).
- R. B. J. Benson, N. E. Campione, M. T. Carrano, Ph. D. Mannion, C. Sullivan, P. Upchurch & D. C. Evans: Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage. In: PLoS Biology, Band 12, Nummer 5, 2014, e1001853, doi:10.1371/journal.pbio.1001853.
- A. Virág & A. Ősi: Morphometry, Microstructure, and Wear Pattern of Neornithischian Dinosaur Teeth From the Upper Cretaceous Iharkút Locality (Hungary). In: The Anatomical Record - Advances in Integrative Anatomy and Evolutionary Biology, Band 300, Nummer 8, 2017, S. 1439–1463, doi:10.1002/ar.23592.
- F. J. Varriale: Dental microwear reveals mammal-like chewing in the neoceratopsian dinosaur Leptoceratops gracilis. In: PeerJ, Band 4, 2016, e2132, doi:10.7717/peerj.2132.
- Matthew C. Herne, J. P. Nair, A. R. Evans & A. M. Tait: New small-bodied ornithopods (Dinosauria, Neornithischia) from the Early Cretaceous Wonthaggi Formation (Strzelecki Group) of the Australian-Antarctic rift system, with revision of Qantassaurus intrepidus Rich and Vickers-Rich, 1999. In: Journal of Paleontology, Band 93, Nummer 3, 2019, S. 543–584 (Digitalisat).
- P. C. Sereno: The Logical Basis of Phylogenetic Taxonomy. In: Systematic Biology, Band 54, Nummer 4, 2005, S. 595–619 (Digitalisat).
- P. Godefroit, G. Garcia, B. Gomez, K. Stein, A. Cincotta, U. Lefèvre & X. Valentin: Extreme tooth enlargement in a new Late Cretaceous rhabdodontid dinosaur from Southern France. In: Nature - Scientific Reports, Band 7, 2017, Artikel Nummer 13098, doi:10.1038/s41598-017-13160-2.
- P.-E. Dieudonné, Th. Tortosa, F. T. Fernández-Baldor, J. I. Canudo & I. Díaz-Martínez: An Unexpected Early Rhabdodontid from Europe (Lower Cretaceous of Salas de los Infantes, Burgos Province, Spain) and a Re-Examination of Basal Iguanodontian Relationships. In: PLoS ONE, Band 11, Nummer 6, 2016, e0156251, doi:10.1371/journal.pone.0156251.
- Z. Csiki-Sava, E. Buffetaut, A. Ősi, X. Pereda-Suberbiola & St. L. Brusatte: Island life in the Cretaceous - faunal composition, biogeography, evolution, and extinction of land-living vertebrates on the Late Cretaceous European archipelago. In: ZooKeys, Band 469, 2015, S. 1–161, doi:10.3897/zookeys.469.8439.
- A. B. Herman & J. Kvaček: Late Cretaceous Grünbach Flora of Austria. Naturhistorisches Museum Wien, Wien, 2010, ISBN 978-3-902421-43-2, 224 S. (Digitalisat).
- G. Botfalvai, J. Haas, E. R. Bodor, A. Mindszenty & A. Ősi: Facies architecture and palaeoenvironmental implications of the upper Cretaceous (Santonian) Csehbánya formation at the Iharkút vertebrate locality (Bakony Mountains, Northwestern Hungary) In: Palaeogeography, Palaeoclimatology, Palaeoecology, Band 441, Nummer 4, 2016, S. 659–678 (Abstract); (Manuskriptversion).
- J. Kocsis, A. Ősi, T. Vennemann, C. N. Trueman & M. R. Palmer: Geochemical study of vertebrate fossils from the Upper Cretaceous (Santonian) Csehbánya Formation (Hungary): Evidence for a freshwater habitat of mosasaurs and pycnodont fish. In: Palaeogeography, Palaeoclimatology, Palaeoecology, Band 280, 2009, S. 532–542,(Abstract).