Massospondylus
Massospondylus (von altgriechisch μάσσων mássōn „größer“ und σφόνδυλος sphóndylos [auch σπόνδυλος spóndylos] „Rückenwirbel, Halswirbel“, also wörtlich „längerer Wirbel“) ist eine ursprüngliche Gattung der sauropodomorphen Dinosaurier. Die Gattung wurde bereits 1854 von dem englischen Anatom Richard Owen beschrieben und zählt damit zu den ersten Dinosauriern, die benannt wurden, auch wenn sie anfangs nicht als solche erkannt wurden. Bis heute sind über 80 Skelette bekannt, die aus dem Unterjura (Hettangium bis Pliensbachium, vor etwa 201 bis 183 Millionen Jahren) von Südafrika, Lesotho und Simbabwe stammen. Damit gehört Massospondylus zu den am besten bekannten ursprünglichen Sauropodomorpha.
| Massospondylus | ||||||||
|---|---|---|---|---|---|---|---|---|

Diese Rekonstruktion zeigt Massospondylus als ein zweibeiniges Tier. | ||||||||
| Zeitliches Auftreten | ||||||||
| Unterer Jura (Hettangium bis Pliensbachium)[1]:151–152 | ||||||||
| 201,3 bis 182,7 Mio. Jahre | ||||||||
| Fundorte | ||||||||
| Systematik | ||||||||
| ||||||||
| Wissenschaftlicher Name | ||||||||
| Massospondylus | ||||||||
| Owen, 1854 | ||||||||
| Arten | ||||||||
| ||||||||
Die Typusart ist Massospondylus carinatus; sieben weitere Arten wurden in den vergangenen 150 Jahren benannt, von denen derzeit allerdings nur Massospondylus kaalae als gültig anerkannt ist. Die genauen Verwandtschaftsbeziehungen von Massospondylus sind umstritten. Häufig wird diese Gattung in eine als Massospondylidae bezeichneten Familie gestellt – es ist allerdings umstritten, ob und welche anderen Dinosaurier zu dieser Gruppe klassifiziert werden sollen.
Massospondylus erreichte Körperlängen von 4 bis 5 Metern und zeichnete sich durch einen langen Hals und Schwanz, einen kleinen Kopf und einen schlanken Körper aus. Wahrscheinlich war er Pflanzenfresser, obwohl einige Forscher vermuten, dass es sich um einen Allesfresser gehandelt haben könnte. Massospondylus wurde traditionell als vierbeinig dargestellt, eine Studie aus dem Jahr 2007 konnte jedoch zeigen, dass es sich tatsächlich um einen Zweibeiner handelte. Fossile Eier mit Skeletten von Embryonen weisen darauf hin, dass frisch geschlüpfte Exemplare vierbeinig waren, und dass der Übergang in eine zweibeinige Lebensweise erst im Lauf der Individualentwicklung stattfand. Außerdem lassen diese Funde elterliche Fürsorge vermuten.
Merkmale
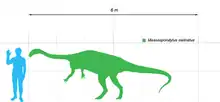
Massospondylus war ein mittelgroßer Vertreter der ursprünglichen Sauropodomorpha. Die Körperlänge wird auf etwa 4[2] bis 5[3]:252[4] Meter, das Gewicht in einer Studie auf 135 Kilogramm,[2] in einer anderen Studie auf bis über 200 Kilogramm[5] geschätzt. Der Bauplan glich dem des Plateosaurus, das Skelett war jedoch insgesamt leichter gebaut. Der Schädel war grazil[6] und proportional kleiner als bei Plateosaurus.[7] Der Hals war im Verhältnis länger als bei den meisten anderen ursprünglichen Sauropodomorpha, die Wirbelkörper der vorderen Halswirbel waren vier Mal so lang wie breit[8]:430 Die Vordergliedmaßen waren nur halb so lang wie die Hintergliedmaßen,[9]:151 aber recht kräftig, worauf das verbreiterte Oberende des Oberarmknochens weist, das Ansatzflächen für eine vergrößerte Armmuskulatur bot[8]:430 Die Hände waren kurz und breit und hatten fünf Finger[8]:430 Vierter und fünfter Finger der Hand waren deutlich kleiner und dünner als die übrigen Finger, was der Hand ein asymmetrisches Aussehen verlieh.[10]:740–750 Der Daumen endete in einer stark vergrößerten, sichelförmigen Kralle und war der längste Finger der Hand.[10]:740–750[8]:430 Der Fuß endete ebenfalls in fünf Zehen, wobei die fünfte Zehe zurückgebildet war und den Boden nicht berührte.[8]:431
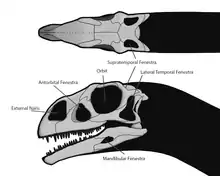
Unterschiede zu verwandten Gattungen finden sich vor allem am Schädel. Dieser weist eine proportional kürzere Schnauze auf[3]:237 und war insgesamt breiter und niedriger als bei anderen ursprünglichen Sauropodomorpha – die größte Breite des Schädels übertrifft die Höhe um 10 %. Die Außenfläche des Schädels wies zahlreiche Öffnungen (Schädelfenster) auf, die das Gewicht reduzierten sowie Ansatzstellen für Muskeln und Platz für Sinnesorgane boten. Die Augenhöhlen waren größer als bei Plateosaurus.[11] Die im vorderen Bereich der Schnauze befindlichen äußeren Nasenöffnungen waren groß, wie es typisch für ursprüngliche Sauropodomorpha war – bei Massospondylus erreichten sie etwa die Hälfte der Größe der Augenhöhlen[3]:237. Das Antorbitalfenster, welches kleiner war als bei verwandten Gattungen wie Plateosaurus, befand sich zwischen Nasenöffnung und Augenhöhle. Hinter den Augen saßen die beiden Schläfenfenster: Das seitliche Schläfenfenster (Infratemporalfenster) ist bei Massospondylus wie ein umgedrehtes „T“ geformt, das in Draufsicht zu sehende obere Schläfenfenster (Supratemporalfenster) spart den hinteren Bereich des Schädeldachs aus. Auch im Unterkiefer befand sich ein kleines Fenster.[12]:242
Die Zähne waren blattförmig und vergleichsweise lang. Ihre Anzahl nahm mit der Schädelgröße zu: im paarigen Oberkiefer saßen je Seite 14 bis 22, im paarigen Unterkiefer waren beim größten bekannten Schädel je 26 Zähne zu finden. Im Prämaxillare, ein vor dem Oberkiefer befindlicher Knochen, saßen bei allen bekannten Exemplaren auf jeder Seite 4 Zähne. Während die Zahngröße im Oberkiefer nach hinten hin abnahm, zeigten die Zähne im Unterkiefer eine konstante Größe. Das Fehlen von Abnutzungsspuren und die unterschiedliche Höhe der Zähne zeigt, dass die Zähne in relativ kurzen Zeitintervallen ausfielen und durch nachrückende Zähne ersetzt wurden.[11]
Wie andere ursprüngliche Sauropodomorpha besaß Massospondylus vermutlich Backen. Hierauf weisen kleine Öffnungen auf der Oberfläche des Ober- und Unterkiefers hin, die Blutgefäße beinhalteten – so waren bei Massospondylus nur wenige, aber große Öffnungen vorhanden, während bei backenlosen Tieren zahlreiche kleine Öffnungen auftraten. Backen hätten verhindert, dass Nahrung beim Fressen aus dem Mund herausfiel.[3]:242/256 Crompton und Attridge (1986) beschrieben zwei Schädel mit einem ausgeprägten Überbiss und schlussfolgerten, dass dieser Längenunterschied durch einen auf der Front des Unterkiefers sitzenden hornigen Schnabel (Rhamphoteca) ausgeglichen wurde.[13] Umstritten ist allerdings, ob dieser Überbiss eventuell eine Fehlinterpretation ist, die der Zerdrückung der Fossilien geschuldet ist.[12]:254[14] Ein Schnabel wird jedenfalls von aktuellen Studien als unwahrscheinlich bezeichnet.[12]:254[14][8]:430
Forschungsgeschichte
.jpg.webp)
Die ersten Fossilien wurden 1854 von dem englischen Anatom und Paläontologen Richard Owen benannt und beschrieben.[15]:97 Owen wählte den Namen Massospondylus (gr. masson/μάσσων – „länger“; spondylos/σπόνδυλος „Wirbel“), „weil die Wirbel proportional länger sind als die des ausgestorbenen Krokodils Macrospondylus“ (“because the vertebrae are proportionally longer than those of the extinct Crocodile called Macrospondylus”).[15]:97 Er erkannte Massospondylus anfangs nicht als einen Dinosaurier, sondern hielt es für ein „großes ausgestorbenes fleischfressendes Reptil“[10]:797 aus der Verwandtschaft der Eidechsen, Chamäleons und Leguane.[15]:80 Die Fossilien waren Teil einer Sammlung aus insgesamt 56 Reptilienknochen, die 1853 von dem Landvermesser Joseph Millard Orpen auf dem Gelände einer Farm in den Drakensbergen bei Harrismith in Südafrika entdeckt und dem Royal College of Surgeons of England als Schenkung überlassen wurden – seitdem wurden sie im zum College gehörenden Hunterian Museum at the Royal College of Surgeons in London aufbewahrt.[15]:97[16][17]:291 Am 10. Mai 1941 zerstörte ein Bombenangriff das Museum inklusive der Fossilien, weshalb heute nurmehr Abbildungen und Abgüsse des ursprünglichen Materials überliefert sind.[16]
Die Knochen sind außerhalb des ursprünglichen Skelettverbundes vorgefunden worden. Trotzdem konnte Owen drei verschiedene Typen von Schwanzwirbeln beschreiben, die er drei verschiedenen Gattungen zuordnete. Die Gattung Massospondylus zeigte laut Owen bemerkenswert lange Schwanzwirbel, jene der zusätzlich aufgestellten Gattungen Pachyspondylus und Leptospondylus seien dagegen deutlich kürzer. Die Sammlung umfasst neben Wirbeln auch diverse Gürtel- und Gliedmaßenknochen, die Owen jedoch keiner der drei Gattungen zuordnen konnte. Später stellte sich heraus, dass es sich bei den als Massospondylus-Schwanzwirbel angesprochenen Knochen tatsächlich um Halswirbel handelte, und dass alle Fossilien vermutlich zur selben Art gehörten. Womöglich handelte es sich um die Überreste von zwei oder mehr Exemplaren.[18]:1033[15][16]
Bis heute wurden mindestens 80 fragmentarische Skelette und 4 Schädel entdeckt, die sowohl Jungtieren als auch erwachsenen Tieren zugeordnet werden können. Dieses Material stammt aus der Oberen Elliot-Formation, der Clarens-Formation und dem Bushveld-Sandstone von Südafrika und Lesotho sowie dem Forest-Sandstone und dem Oberen Karroo-Sandstone von Simbabwe.[3]:235 Massospondylus wurde außerdem aus der Kayenta-Formation von Arizona (USA) und aus der Canon-del-Colorado-Formation von San Juan (Argentinien) beschrieben – diese Fossilien wurden jedoch kürzlich eigenständigen, nahe verwandten Gattungen zugeordnet. Der Nachweis aus Arizona basiert auf einem gut erhaltenen, 1985 beschriebenen Schädel.[19]:585 Im Jahr 2010 wurde dieser Schädel zusammen mit Resten des Körperskeletts zur neuen Gattung Sarahsaurus überführt.[20] Das argentinische Material, das aus verschiedenen fragmentarischen Skeletten und mindestens einem Schädel besteht, wurde 2009 als Adeopapposaurus beschrieben.[21]
Arten
Die von Richard Owen beschriebene Typusart ist M. carinatus.[15]:97 Bis heute wurden zahlreiche weitere Arten benannt, die heute jedoch, mit Ausnahme von M. kaalae (Barrett, 2009), entweder als Synonyme von M. carinatus oder als Nomina dubia (unbestimmbar) gelten.[16] Diese heute nicht mehr anerkannten Arten schließen M. browni (Seeley, 1895),[22] M. harriesi (Broom, 1911),[17]:299–300 M. hislopi (Lydekker, 1890),[23] M. huenei (Cooper, 1981),[10]:804 M. rawesi (Lydekker, 1890)[23] und M. schwarzi (Haughton, 1924)[24] mit ein.
M. browni, M. harriesi und M. schwarzi stammen aus der Oberen Elliot-Formation aus der Kapprovinz in Südafrika. Alle drei Arten basieren auf fragmentarischen Resten und werden in einem Review-Artikel von 2004 als Nomina dubia deklariert.[3]:236 M. browni basiert auf zwei Hals-, zwei Rücken- und drei Schwanzwirbeln sowie verschiedenen Resten des Hinterbeins.[22] M. harriesi wurde anhand einer gut erhaltenen Vordergliedmaße und Teilen der Hintergliedmaße beschrieben.[17]:299–300 M. schwarzi wurde anhand eines unvollständigen Hinterbeins und eines Kreuzbeins beschrieben.[24] Als M. hislopi und M. rawesi wurden Fossilien aus Indien bezeichnet.[23] M. hislopi basiert auf Wirbeln aus der obertriassischen Maleri-Formation von Andhra Pradesh, M. rawesi auf einem einzelnen Zahn aus der oberkreidezeitlichen Takli-Formation von Maharashtra.[25] Der Name M. hislopi wurde in dem letzten Review-Artikel vorläufig als ein zu den Sauropodomorpha zu stellendes Nomen dubium beibehalten,[3]:236 bei M. rawesi jedoch könnte es sich um einen Theropoden[26] oder um einen Nicht-Dinosaurier[19]:586 gehandelt haben. M. huenei ist eine neue Kombination, die Michael Cooper von Lufengosaurus huenei ableitete, da er Lufengosaurus und Massospondylus als Synonyme betrachtete.[10]:804 Diese Synonymität wird jedoch nicht länger akzeptiert.[3]:235
M. kaalae, neben M. carinatus die einzige derzeit anerkannte Art, wurde 2009 anhand eines fragmentarischen Schädels beschrieben, der in den Schichten der Oberen Elliot-Formation nahe dem Dorf Voyizane in Ostkap gefunden wurde. Aus dieser Region stammen außerdem verschiedene Funde des zeitgenössischen Massospondylus carinatus. M. kaalae unterscheidet sich von der Typusart durch die Morphologie des Hirnschädels sowie verschiedenen anderen Schädelmerkmalen, beispielsweise durch die Proportionen der Prämaxillare.[18]
Synonyme
Wie alle Tiere zeigte auch Massospondylus innerartliche (individuelle) und ontogenetische (altersbedingte) Variation, was die Zuordnung bestimmter Funde erschwert. In der Vergangenheit wurden eine Reihe von Gattungen beschrieben, deren Unterschiede zu anderen Massospondylus-Fossilien heute auf ebendiese Variationen zurückgeführt werden und die deshalb meist als Synonyme von Massospondylus gelten. Zu den Synonymen zählen die bereits oben erwähnten Gattungen Leptospondylus und Pachyspondylus (Owen, 1854) sowie Aristosaurus, Dromicosaurus, Gryponyx taylori und Hortalotarsus.[10]:691
Hortalotarsus skirtopodus basiert auf Resten eines Hinterbeins, die 1888 bei Barkly East in Ostkap entdeckt und 1894 von Harry Seeley beschrieben wurden.[27] Robert Broom (1911) bemerkt:[17]:293
“Originally most of the skeleton was in the rock, and it was regarded by the farmers as the skeleton of a Bushman, but it is said to have been destroyed through fear that a Bushman skeleton in the rock might tend to weaken the religious belief of the rising generation.”
„Ursprünglich war der Großteil des Skelettes im Felsen vorhanden, und es wurde von den Bauern für das Skelett eines Buschmanns gehalten; es wird jedoch gesagt, dass es zerstört wurde aus Angst, ein Buschmann-Skelett im Felsen könnte den religiösen Glauben der aufstrebenden Generation schwächen.“
Aristosaurus erectus wurde von E.C.N. van Hoepen (1920) beschrieben und gründet auf einem nahezu vollständigen, wenn auch schädellosen Skelett, das beim Abbau in einem Naturstein-Bruch nahe Roosendal entdeckt und 1915 vom Transvaal-Museum aufgekauft wurde.[28] Hoepen beschrieb außerdem Dromicosaurus gracilis anhand eines von ihm entdeckten, fragmentarischen Skeletts.[19]:584 Gryponyx taylori wurde von Sidney H. Haughton (1924) benannt, es basiert auf 1915 bei Fouriesburg entdeckte Beckengürtelknochen und Kreuzbeinwirbel.[19]:584 Alle diese Arten stammen aus der Oberen Elliot-Formation von Südafrika, aus welcher auch Massospondylus stammt. Nach den Internationalen Regeln für die Zoologische Nomenklatur (ICZN) handelt es sich um jüngere Synonyme: Sie wurden benannt, als Massospondylus bereits wissenschaftlich beschrieben war; der Name Massospondylus hat daher Priorität.
Systematik
| Kladogramm, vereinfacht nach Yates (2007)[29] | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| In diesem Review wurde Massospondylus als naher Verwandter von Lufengosaurus und Coloradisaurus klassifiziert. Das Kladogramm unterstützt die Hypothese der „extremen Prosauropoden-Paraphylie“. Dies ist nur eines von vielen vorgeschlagenen Kladogrammen zur Systematik der ursprünglichen Sauropodomorpha. |
Die Sauropodomorpha werden traditionell in zwei Gruppen eingeteilt: die Prosauropoden und die Sauropoden. Diese Klassifikation gilt heute als widerlegt, da sich die Sauropoden von Vertretern der klassischen Prosauropoden ableiten – somit schließen die „Prosauropoden“ die Sauropoden mit ein und sind damit paraphyletisch. Aktuelle Diskussionen drehen sich um die Frage, wie stark diese Paraphylie ausgeprägt ist: Anhänger der „Kern-Prosauropoden-Monophylie“ sind der Auffassung, dass einige Kerngruppen der klassischen Prosauropoden – die Massospondylidae, die Plateosauridae und die Riojasauridae – eine monophyletische Gruppe bilden, während Anhänger der „extremen Prosauropoden-Paraphylie“ zu dem Ergebnis gelangen, dass auch diese Gruppen sich unabhängig voneinander von der zu den Sauropoden führenden Entwicklungslinie abspalteten.[8]:426–428
Massospondylus wird häufig innerhalb der Familie Massospondylidae klassifiziert. Diese Gruppe enthält per Definition alle Gattungen, die näher mit Massospondylus verwandt sind als mit Plateosaurus. Die Gattungen, die nach dieser Definition innerhalb der Massospondylidae zu klassifizieren und damit die nächsten Verwandten von Massospondylus sind, variieren je nach Studie. Häufig werden Coloradisaurus, Yunannosaurus und Lufengosaurus als Massospondyliden erkannt.[29][30][31] Andere Forscher kamen allerdings zu dem Ergebnis, dass Massospondylus der einzige Massospondylide ist und betrachten die Familie als obsolet.[32]
Vorkommen und Paläohabitat
Massospondylus-Fossilien stammen aus der Karoo-Supergruppe in Südafrika, Lesotho und Simbabwe.[3]:235 Diese gewaltige Schichtfolge kam vom Oberperm bis zum Unterjura zur Ablagerung und ist die weltweit bedeutendste kontinentale Fossillagerstätte in diesem Zeitintervall.[1]:135 Massospondylus tritt in unterjurassischen Schichten der Karoo-Supergruppe auf – das Vorkommen reicht vom frühesten Unterjura unmittelbar oberhalb der Trias-Jura-Grenze bis vor etwa 183 Millionen Jahren,[1]:151–152 als gewaltige Lavaflüsse (Flutbasalte) die Ablagerung der Karoo-Supergruppe zum Abschluss brachten.[33] Massospondylus ist das am häufigsten gefundene Fossil dieses bisher noch wenig erforschten Schichtintervalls,[34]:84 das auch als Massospondylus range zone bezeichnet wird.[1]:151–152
Massospondylus lebte in einem recht trockenen, wüstenartigen Klima.[10]:807–812 Den Lebensraum teilte es mit Temnospondyli, Schildkröten, dem Sphenodontier Clevosaurus, Rauisuchier, Krokodilverwandten wie Protosuchus und Ornithosuchus, theropode Dinosaurier wie Megapnosaurus, ornithopode Dinosaurier wie Lesothosaurus, Abrictosaurus, Heterodontosaurus, Lycorhinus und Pegomastax[35] sowie mit verschiedenen Cynodontiern.[34]:81–85 In der Oberen Elliot-Formation und der Clarens-Formation, aus denen ein Großteil der Massospondylus-Fossilien stammt, galt Massospondylus carinatus lange als der einzige bekannte Sauropodomorphe.[34]:84 Paul Barrett (2009) bemerkte jedoch, dass mindestens vier weitere Sauropodomorphen, darunter zwei Sauropoden, vorliegen, wenn auch nur mit fragmentarischen Resten.[18]:1032 Jüngst wurden mit Ignavusaurus[30] und Arcusaurus[36] zwei weitere Sauropodomorphen aus der Oberen Elliot-Formation beschrieben.
Paläobiologie
Fortbewegung und Beweglichkeit

Lange wurde angenommen, dass sich diese Tiere vor allem quadruped (auf vier Beinen laufend) fortbewegten. Eine 2007 publizierte biomechanische Studie von Matthew Bonnan und Phil Senter kam jedoch zu dem Schluss, dass Massospondylus und der verwandte Plateosaurus obligat biped waren, sich also ausschließlich zweibeinig fortbewegten. So war es diesen Tieren unmöglich, die Hand zu pronieren (die Handfläche nach unten zu drehen). Hierauf weisen auch in-situ-Funde von Armen, die noch im ursprünglichen Skelettverbund erhalten sind und die stets einander gegenüberstehende Handflächen zeigen. Auch der sehr eingeschränkte Bewegungsraum der Arme deutet auf eine bipede Fortbewegungsweise.[9]:147–148[38]
Robert Bakker (1987) vermutet, dass das Schulterblatt bei quadrupeden Dinosauriern mit den Vorderbeinen pendelte und somit als eine Art verlängertes Bein die Schrittweite vergrößerte. Dieses Merkmal findet sich bei vielen heutigen, quadrupeden Säugetieren und könnte bei einem quadrupeden Massospondylus den eingeschränkten Bewegungsraum der Arme ausgeglichen haben[9]:147–148. Voraussetzung dabei ist jedoch, dass die Schlüsselbeine zurückgebildet waren – funktionelle Schlüsselbeine wirken wie eine Spange zwischen dem rechten und linken Schulterblatt und verhindern für die Fortbewegung relevante Rotationen dieses Knochens. Tatsächlich zeigen sich bei der Gruppe der Ceratopsia stark reduzierte Schlüsselbeine, und die damals dominierende Ansicht weitete diesen Befund auf alle Dinosauriergruppen abseits der zu den Vögeln führenden Entwicklungslinie aus.[39]
Eine jüngere Entdeckung zeigt allerdings, dass Massospondylus gut ausgebildete Schlüsselbeine besaß, die an ihren Enden miteinander V-förmig verbunden waren und somit dem Gabelbein der Vögel glichen. Dieser Fund beweist, dass der Schultergürtel tatsächlich relativ statisch war und keinen relevanten aktiven Beitrag zur Fortbewegung leisten konnte. Die Reduktion der Schlüsselbeine war folglich eine Innovation, die sich auf die zu den Ceratopsia führende Entwicklungslinie beschränkte. Der Nachweis von Schlüsselbeinen bei Massospondylus ist außerdem ein weiteres Indiz dafür, dass das Gabelbein der heutigen Vögel aus der Verschmelzung der beiden Schlüsselbeine hervorgegangen ist.[40]
Michael Cooper (1981) stellte fest, dass die Gelenkfortsätze der Halswirbel gegenüber der Horizontalen um 45° gekippt waren. Dies führte laut diesem Forscher dazu, dass der Hals zwar vertikale, aber keine seitlichen Bewegungen erlaubte. Auch die restliche Wirbelsäule sei in ihrer horizontalen Beweglichkeit sehr eingeschränkt gewesen, weshalb das Tier bei einer Drehung des Halses stets den gesamten Körper mitgedreht haben müsse.[10]:720 Paul Barrett und Paul Upchurch (2007) widersprechen und geben an, dass die Gelenkfortsätze lediglich an der Halsbasis gekippt waren, in den vorderen Halswirbeln aber fast waagerecht standen, was eine ausreichende seitliche Beweglichkeit des Halses erlaubt habe.[14]
Ernährung
Massospondylus und verwandte Gattungen waren wahrscheinlich Pflanzen- oder Allesfresser. Die Zähne waren gerade, blattförmig und grob gesägt und damit gut zum Zerkleinern von pflanzlicher Nahrung geeignet. Die Zahnkronen waren breiter als die Zahnwurzeln, was die Lücken zwischen den Zähnen reduzierte, sodass die Zahnreihe eine durchgehende Schneidekante zum Abschneiden von Pflanzenmaterial bildete.[8]:434 Paul Barrett (2000) bemerkt jedoch, dass die Zähne des Prämaxillare sowie die vordersten Zähne des Oberkiefers von den restlichen Zähnen abweichen: Diese Zähne waren leicht nach hinten gekrümmt und fein gesägt – damit zeigten sie Gemeinsamkeiten mit den Zähnen fleischfressender Dinosaurier. Barrett folgerte, dass Massospondylus als Allesfresser seinen Speiseplan mit kleinen Beutetieren wie kleinen Reptilien, Säugetieren und Wirbellosen aufgebessert haben könnte.[41] Michael Cooper (1981) vermutete, dass ein Anteil der Nahrung aus Aas bestanden hat, und spekulierte, dass die sichelförmige Handkralle zum Aufbrechen von Kadavern gedient haben könnte.[10]:814–815
Raath (1974) berichtete über die Entdeckung von Gastrolithen (Magensteinen) in drei Massospondylus-Skeletten aus dem Forest-Sandstone von Zimbabwe.[42] Lange wurde angenommen, dass diese von den Tieren verschluckten Steine zum Zermahlen der Nahrung im Magen gedient haben, ähnlich wie es bei heutigen Vögeln der Fall ist. Oliver Wings und Martin Sander (2007) zeigten jedoch, dass diese Steine keinen relevanten Beitrag zur Verdauung leisten konnten: So waren die Steine poliert und nicht rau wie bei Vögeln.[43] Außerdem zeigte nur eines der Massospondylus-Skelette genug Gastrolithe, um eine effektive Magenmühle zu bilden.[8]:435
Wachstum

Aussagen über das Wachstum und über das individuelle Lebensalter fossiler Wirbeltiere können aus Wachstumsringen und anderen Strukturen der äußeren Knochenschicht (Kortikalis) abgeleitet werden. Massospondylus zeigte eine Phase schnellen Wachstums bis zum 15. Lebensjahr, in der bis zu 34,6 kg pro Jahr zugelegt wurden – diese Rate ist vergleichbar mit der rezenter Säugetiere. Anders als bei Säugetieren gab es allerdings keinen Wachstumsstopp; auch nach dem 15. Lebensjahr nahm die Körpergröße, wenn auch deutlich verlangsamt, weiterhin zu.[5][44]
Martin Sander und Nicole Klein (2005) untersuchten das Wachstum der nahe verwandten Gattung Plateosaurus und zeigten, dass diese Tiere ihr Wachstum an die jeweiligen Umweltbedingungen anpassten. Bei reichem Nahrungsangebot oder guten klimatischen Bedingungen war das Wachstum beschleunigt und die Phase schnellen Wachstums verlängert. Dieses als Entwicklunsplastizität bekannte Phänomen tritt heute bei ektothermen Tieren wie Reptilien auf. Bei den endothermen (warmblütigen) Säugetieren und Vögeln folgt das Wachstum dagegen einer definierten Wachstumskurve – die Wachstumsrate und die endgültige Körpergröße variieren zwischen Individuen nur wenig. Plateosaurus ist der einzige Dinosaurier und damit das vermutlich einzige bekannte endotherme Tier, bei dem Entwicklungsplastizität nachgewiesen wurde – alle anderen entsprechend untersuchten Dinosaurier einschließlich Massospondylus entsprechen Säugetieren und Vögeln. Die Forscher folgern: “… [Plateosaurus] possibly represents the initial stage in the evolution of metabolic thermoregulation (endothermy) in dinosaurs, in which endothermy was decoupled from developmental plasticity” („… Plateosaurus repräsentiert möglicherweise ein Anfangsstadium in der Evolution der metabolischen Thermoregulation (Endothermie) bei Dinosauriern, in dem Endothermie und Entwicklungsplastizität noch voneinander entkoppelt waren“).[45]:1802[45]
Reproduktion und Entwicklung
%252C_Golden_Gate_National_Park%252C_South_Africa%252C_Early_Jurassic_-_Royal_Ontario_Museum_-_DSC00145.JPG.webp)

Ein bedeutender Fund gelang 1976, als der Paläontologe James Kitching einen Block mit sechs fossilen Eiern an einem Straßenaufschluss im südafrikanischen Golden Gate Highlands National Park fand.[46]:1653 Diese rundlichen und im Durchmesser bis zu 6 cm messenden Eier enthielten Überreste von Embryonen. Eine solche in-ovo-Erhaltung (Erhaltung innerhalb von Eiern) ist bei Dinosauriern äußerst selten, tatsächlich ist dieser Fund bis heute der älteste bekannte. Erst seit 2006 wird die heute als Rooidraai-Lokalität bekannte Fundstelle von Forschern um Robert Reisz genauer untersucht, wobei insgesamt 10 Gelege mit Eiern entdeckt wurden, einige ebenfalls mit Embryonenresten. Diese Entdeckung bietet einzigartige Einblicke in die Reproduktions- und Entwicklungsbiologie dieser Tiere.[47][48]
Alle entdeckten Gelege waren unvollständig; das vollständigste entdeckte Gelege enthielt 34 Eier. Die Gelege wurden in mindestens drei verschiedenen stratigraphischen Schichten gefunden. Hieraus lässt sich schließen, dass an diesem Ort mindestens zwei Massospondylus-Exemplare zu mindestens vier verschiedenen Zeitpunkten genistet haben. Wie viele Nester tatsächlich vorhanden waren, lässt sich wegen der Lage des Fundorts an einem Steilhang nicht ermitteln. Vermutlich handelte es sich um eine Nistkolonie, die sich periodisch an diesem Ort versammelte. Die Gelege wurden in unmittelbarer Wassernähe in weichem Sediment angelegt, worauf Bioturbation (durch wirbellose Tiere angelegte Grabgänge) hinweist. Vermutlich wurden die Eier zumindest teilweise im Sediment begraben, worauf die sehr dünnen, nur 0,1 mm dicken Eierschalen hinweisen. Die Eierschale ermöglicht den Gasaustausch zwischen Ei und Umgebung – eine dünne Eierschale stellt den Gasaustausch auch in sauerstoffarmer und kohlendioxidreicher Umgebung sicher. Hinweise auf Nestbau fehlen, die Eier waren jedoch eng gepackt in Reihen angeordnet. Die Körpergröße der erwachsenen Tiere lässt vermuten, dass die Eier nach dem Legen vom Muttertier aktiv in diese Position geschoben wurden.[47]
Die gefundenen Embryonen-Skelette stammen wahrscheinlich von Tieren, die kurz vor dem Schlupf standen.[46]:1664 Das embryonale Skelett unterscheidet sich von dem adulter Tiere nur in den deutlich anderen Proportionen. Der Schädel ist auffällig groß mit einer kurzen Schnauze und sehr große Augenöffnungen, deren Durchmesser 39 % der Schädellänge beträgt. Die Halswirbel sind kurz und stehen damit im Kontrast zu den stark verlängerten Halswirbeln erwachsener Tiere. Schambein und Sitzbein sowie die Schwanzwirbel sind ebenfalls proportional kleiner als bei Skeletten erwachsener Tiere. Auffällig sind außerdem die deutlich längeren Vordergliedmaßen. Der große Kopf und die langen Vordergliedmaßen zeigen, dass sich diese Tiere nach dem Schlüpfen quadruped fortbewegt haben müssen, und dass es in einem späteren Entwicklungsstadium einen Übergang zur Bipedie gegeben hat, wie sie erwachsene Tiere zeigen.[48] Skelette von frisch geschlüpften Tieren sind von einem anderen Sauropodomorphen bekannt, Mussaurus.[9]:152[46]:1664 Diese Embryonenreste gleichen denen von Massospondylus, weshalb davon ausgegangen werden kann, dass auch andere ursprüngliche Sauropodomorphen zuerst quadruped waren und erst in der weiteren Entwicklung biped wurden.[9]:152
Die Gruppe Dinosauria leitet sich von bipeden Vorfahren ab.[6] Verschiedene Entwicklungslinien innerhalb der Dinosaurier, unter anderem die zu den Sauropoden führende, gingen jedoch von der bipeden in eine quadrupede Fortbewegung über. Die Quadrupedie im Jugendstadium von Massospondylus erlaubt einzigartige Rückschlüsse auf den Evolutionsmechanismus, der zur Quadrupedie der Sauropoden führte: Dieser Übergang von einer bipeden in eine quadrupede Lebensweise ist demnach auf Pädomorphose zurückzuführen, das Beibehalten ursprünglich nur in Jugendstadien vorhandener Merkmale auch bei erwachsenen Tieren.[48] Reisz und Kollegen diskutierten außerdem die Frage, warum Jungtiere ursprünglicher sauropodomorpher Dinosaurier abweichend vom ursprünglichen bipeden Zustand quadruped waren: So sei der Hals im ursprünglichen Zustand S-förmig gebogen gewesen, wurde von Sauropodomorphen jedoch als Anpassung auf eine pflanzliche Ernährung gerade gehalten. Diese Modifikation, die den Körperschwerpunkt nach vorne verlagert, könnte laut diesen Forschern im Zusammenspiel mit den großen Köpfen für die Evolution der Quadrupedie bei frisch geschlüpften Tieren verantwortlich gewesen sein.[48]
Obwohl die Jungtiere nach dem Schlüpfen quadruped waren, zeigten sie keine Anpassungen an eine Pronation der Hand, sie konnten also die Handinnenfläche bei vertikal stehendem Arm nicht nach unten drehen. Zusammen mit den kleinen Beckenknochen und dem großen Kopf weist dies darauf hin, dass eine effektive Fortbewegung nicht möglich war.[9]:152 Die Tiere könnten somit Nesthocker gewesen sein, was einen gewissen Grad an elterlicher Fürsorge voraussetzt. Diese Interpretation wird durch das Fehlen von Zähnen bei den Embryonen sowie durch Fußspuren unterstützt, die bei den Gelegen gefunden wurden und frisch geschlüpften Massospondylus zugeordnet werden können. Diese Fußspuren zeigen sowohl Hand- als auch Fußabdrücke, was die aus den Skelettproportionen geschlussfolgerte Quadrupedie bestätigt. Die Handabdrücke lassen erkennen, dass die Handflächen nach innen und der Daumen nach vorne zeigte, was bestätigt, dass die Hände nicht proniert waren. Zudem maßen die größten Abdrücke etwa 15 mm, während ein frisch geschlüpftes Tier einen nur 7 mm großen Abdruck hinterlassen hätte, was zeigt, dass die Tiere nach dem Schlupf für einige Zeit am Nistplatz geblieben waren.[48][47]
Weblinks
Einzelnachweise
- Bruce S. Rubidge: 27th Du Toit Memorial Lecture. Re-uniting lost continents – Fossil reptiles from the ancient Karoo and their wanderlust. In: South African Journal of Geology. Band 108, Nr. 1, 2005, ISSN 0371-7208, S. 135–172, doi:10.2113/108.1.135.
- Frank Seebacher: A new method to calculate allometric length-mass relationships of dinosaurs. In: Journal of Vertebrate Paleontology. Band 21, Nr. 1, 2001, ISSN 0272-4634, S. 51–60, doi:10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2.
- Peter Galton, Paul Upchurch: Prosauropoda. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 232–258.
- Cécilio C. Vasconcelos, Adam M. Yates: Sauropodomorph biodiversity of the upper Elliot Formation (Lower Jurassic) of southern Africa. In: Geoscience Africa 2004. Abstract Volume 2. University of the Witwatersrand, Johannesburg 2004, ISBN 0-620-32470-8, S. 670.
- Gregory M. Erickson, Kristina Curry Rogers, Scott A. Yerby: Dinosaurian growth patterns and rapid avian growth rates. In: Nature. Band 412, Nr. 6845, 2001, S. 429–433, doi:10.1038/35086558.
- Paul C. Sereno: The origin and evolution of dinosaurs. In: Annual Review of Earth and Planetary Sciences. Band 25, Nr. 1, 1997, ISSN 0084-6597, S. 435–489, hier S. 436–437, 451, doi:10.1146/annurev.earth.25.1.435.
- Heinrich Mallison: It’s time for even more Plateosaurus! (prosauropod proportions). In: dinosaurpalaeo. Abgerufen am 8. August 2014.
- Adam M. Yates: Basal Sauropodomorpha: The „Prosauropods“. In: Michael K. Brett-Surman, Thomas R. Holtz Jr., James O. Farlow (Hrsg.): The Complete Dinosaur. 2nd edition. Indiana University Press, Bloomington IN 2012, ISBN 978-0-253-35701-4, S. 425–443.
- Matthew F. Bonnan, Phil Senter: Were the basal sauropodomorph dinosaurs Plateosaurus and Massospondylus habitual quadrupeds? In: Paul M. Barrett, David J. Batten (Hrsg.): Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs (= Special Papers in Palaeontology. Band 77). The Palaeontological Association, London 2007, ISBN 978-1-4051-6933-2, S. 139–155.
- Michael R. Cooper: The Prosauropod Dinosaur Massospondylus carinatus Owen From Zimbabwe: Its Biology, Mode of Life and Phylogenetic Significance (= Occasional Papers of the National Museums and Monuments of Rhodesia. Series B: Natural Sciences. Band 6, Nr. 10, ZDB-ID 405377-1). National Museums and Monuments, Salisbury 1981.
- Christoper E. Gow, James W. Kitching, Michael A. Raath: Skulls of the prosauropod dinosaur Massospondylus carinatus Owen in the collections of the Bernard Price Institute for Palaeontological Research. In: Palaeontologia Africana. Band 27, 1990, ISSN 0078-8554, S. 45–58, hier 54–56.
- Hans-Dieter Sues, Robert R. Reisz, Sanja Hinic, Michael A. Raath: On the skull of Massospondylus carinatus Owen, 1854 (Dinosauria: Sauropodomorpha) from the Elliot and Clarens formations (Lower Jurassic) of South Africa. In: Annals of Carnegie Museum. Band 73, Nr. 4, 2004, ISSN 0097-4463, S. 239–257.
- Alfred W. Crompton, John Attridge: Masticatory apparatus of the larger herbivores during Late Triassic and Early Jurassic times. In: Kevin Padian: The Beginning of the Age of Dinosaurs. Faunal Change Across the Triassic–Jurassic Boundary. Cambridge University Press, Cambridge u. a. 1986, ISBN 0-521-30328-1, S 223–236, hier S. 227–228.
- Paul M. Barrett, Paul Upchurch: The evolution of feeding mechanisms in early sauropodomorph dinosaurs. In: Paul M. Barrett, David J. Batten (Hrsg.): Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs (= Special Papers in Palaeontology. Band 77). The Palaeontological Association, London 2007, ISBN 978-1-4051-6933-2, S. 98–99.
- Descriptive catalogue of the fossil organic remains of Reptilia and Pisces contained in the museum of the Royal College of Surgeons of England. Taylor and Francis, London 1854.
- Adam M. Yates, Paul M. Barrett: Massospondylus carinatus Owen 1854 (Dinosauria: Sauropodomorpha) from the Lower Jurassic of South Africa: Proposed conservation of usage by designation of a neotype. In: Palaeontologia Africana. Band 45, 2010, S. 7–10, Digitalisat (PDF; 540 kB)
- Robert Broom: On the dinosaurs of the Stormberg, South Africa. In: Annals of the South African Museum. Band 7, Nr. 4, Article 13, 1911, ISSN 0303-2515, S. 291–308, Digitalisat.
- Paul M. Barrett: A new basal sauropodomorph dinosaur from the upper Elliot Formation (Lower Jurassic) of South Africa. In: Journal of Vertebrate Paleontology. Band 29, Nr. 4, 2009, S. 1032–1045, doi:10.1671/039.029.0401.
- Donald F. Glut: Dinosaurs. The Encyclopedia. McFarland & Company, Jefferson NC u. a. 1997, ISBN 0-89950-917-7.
- Timothy B. Rowe, Hans-Dieter Sues, Robert R. Reisz: Dispersal and diversity in the earliest North American sauropodomorph dinosaurs, with a description of a new taxon. In: Proceedings of the Royal Society. Series B: Biological Sciences. Band 278, Nr. 1708, 2011, ISSN 0950-1193, S. 1044–1053, doi:10.1098/rspb.2010.1867.
- Ricardo N. Martínez: Adeopapposaurus mognai, gen. et sp. nov (Dinosauria: Sauropodomorpha), with comments on adaptations of basal sauropodomorpha. In: Journal of Vertebrate Paleontology. Band 29, Nr. 1, 2009, S. 142–164, doi:10.1671/039.029.0102.
- Harry G. Seeley: On the type of the genus Massospondylus and on some Vertebrae and limb-bone of M. (?) browni. In: The Annals and Magazine of Natural History. Series 6, Band 15, Nr. 85, Article 12, 1895, ZDB-ID 280102-4, S. 102–125, doi:10.1080/00222939508677852, Digitalisat.
- Richard Lydekker: Note on certain vertebrate remains from the Nagpur district. In: Records of the Geological Survey of India. Band 23, Nr. 1, 1890, ISSN 0370-5226, S. 21–24.
- Sydney H. Haughton: The fauna and stratigraphy of the Stormberg Series. In: Annals of the South African Museum. Band 12, Nr. 8, Article 17, 1924, S. 323–497, Digitalisat.
- David B. Weishampel, Paul M. Barret, Rodolfo A. Coria, Jean Le Loeuff, Xu Xing, Zhao Xijin, Ashok Sahni, Elizabeth M. Gomani, Christopher R. Noto: Dinosaur Distribution. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 517–683, hier S. 525 und 595.
- Ralph E. Molnar, David B. Weishampel, Halszka Osmólska, Peter Dodson: Problematic Theropoda: „Carnosaurs“. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. University of California Press, Berkeley CA u. a. 1990, ISBN 0-520-06726-6, S. 306–317.
- Harry G. Seeley: On Hortalotarsus skirtopodus, a new Saurischian fossil from Barkly East, Cape Colony. In: The Annals and Magazine of Natural History. Series 6, Band 14, Nr. 84, Article 53, 1894, S. 411–419, doi:10.1080/00222939408677828, Digitalisat.
- Egbert C. N. van Hoepen: Contributions to the knowledge of the reptiles of the Karroo Formation. 5. A New Dinosaur from the Stormberg Beds. In: Annals of the Transvaal Museum. Band 7, Nr. 2, 1920, ISSN 0041-1752, S. 77–92.
- Adam M. Yates: The first complete skull of the Triassic dinosaur Melanorosaurus Haughton (Sauropodomorpha: Anchisauria). In: Paul M. Barrett, David J. Batten (Hrsg.): Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs (= Special Papers in Palaeontology. Band 77). The Palaeontological Association, London 2007, ISBN 978-1-4051-6933-2, S. 9–55, hier S. 31.
- Fabien Knoll: A primitive sauropodomorph from the upper Elliot Formation of Lesotho. In: Geological Magazine. Band 147, Nr. 6, 2010, ISSN 0016-7568, S. 814–829, hier S. 814 und 826, doi:10.1017/S001675681000018X.
- Diego Pol, Alberto Garrido, Ignacio A. Cerda: A new sauropodomorph dinosaur from the Early Jurassic of Patagonia and the origin and evolution of the sauropod-type sacrum. In: PLoS ONE. Band 6, Nr. 1, 2011, e14572, doi:10.1371/journal.pone.0014572.
- Paul Upchurch, Paul M. Barrett, Peter M. Galton: A phylogenetic analysis of basal sauropodomorph relationships: Implications for the origin of sauropod dinosaurs. In: Paul M. Barrett, David J. Batten (Hrsg.): Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs (= Special Papers in Palaeontology. Band 77). The Palaeontological Association, London 2007, ISBN 978-1-4051-6933-2, S. 57–90, hier S. 64, Abstract.
- Roger M. H. Smith, Patrick G. Eriksson, Willem J. Botha: A review of the stratigraphy and sedimentary environments of the Karoo-aged basins of Southern Africa. In: Journal of African Earth Sciences. Band 16, Nr. 1/2, 1993, ISSN 1464-343X, S. 143–169, doi:10.1016/0899-5362(93)90164-L.
- Fabien Knoll: The tetrapod fauna of the Upper Elliot and Clarens formations in the main Karoo Basin (South Africa and Lesotho). In: Bulletin de la Société géologique de France. Band 176, Nr. 1, 2005, ISSN 0037-9409, S. 81–91, doi:10.2113/176.1.81.
- Paul C. Sereno: Taxonomy, morphology masticatory function and phylogeny of heterodontosaurid dinosaurs (= ZooKeys. Band 226, Special Issue). Pensoft, Sofia 2012, ISBN 978-954-642-652-9, S. 149.
- Adam M. Yates, Matthew F. Bonnan, Johann Neveling: A new basal sauropodomorph dinosaur from the Early Jurassic of South Africa. In: Journal of Vertebrate Paleontology. Band 31, Nr. 3, 2011, S. 610–625, doi:10.1080/02724634.2011.560626.
- William Lindsay, Nigel Larkin, Neil Smith: Displaying Dinosaurs at The Natural History Museum, London. In: Curator. The Museum Journal. Band 39, Nr. 4, 1996, ISSN 0011-3069, S. 270–272, doi:10.1111/j.2151-6952.1996.tb01102.x.
- Heinrich Mallison; The digital Plateosaurus II: An assessment of the range of motion of the limbs and vertebral column and of previous reconstructions using a digital skeletal mount. In: Acta Palaeontologica Polonica. Band 55, Nr. 3, 2010, ISSN 0567-7920, S. 433–458, doi:10.4202/app.2009.0075.
- Robert Bakker: The return of the dancing dinosaurs. In: Sylvia J. Czerkas, Everett C. Olson (Hrsg.): Dinosaurs past and present. Band 1. University of Washington Press, Seattle WA 1987, ISBN 0-938644-24-6, S. 38–69.
- Adam M. Yates, Celcilio C. Vasconcelos: Furcula-like clavicles in the prosauropod dinosaur Massospondylus. In: Journal of Vertebrate Paleontology. Band 25, Nr. 2, 2005, S. 466–468, doi:10.1671/0272-4634(2005)025[0466:FCITPD]2.0.CO;2.
- Paul M. Barrett: Prosauropod dinosaurs and iguanas: speculations on the diets of extinct reptiles. In: Hans-Dieter Sues (Hrsg.): Evolution of herbivory in terrestrial vertebrates. Perspectives from the fossil record. Cambridge University Press, Cambridge u. a. 2000, ISBN 0-521-59449-9, S. 42–78, hier S. 53, doi:10.1017/CBO9780511549717.004.
- Michael A. Raath: Fossil vertebrate studies in Rhodesia. Further evidence of gastroliths in prosauropod dinosaurs (= Arnoldia. Band 7, Nr. 5, ISSN 0066-7781). National Museums and Monuments of Rhodesia, Salisbury 1974.
- Oliver Wings, P. Martin Sander: No gastric mill in sauropod dinosaurs: new evidence from analysis of gastrolith mass and function in ostriches. In: Proceedings of the Royal Society. Series B: Biological Sciences. Band 274, Nr. 1610, 2007, S. 635–640, doi:10.1098/rspb.2006.3763.
- Anusuya Chinsamy: Bone histology and growth trajectory of the prosauropod dinosaur Massospondylus carinatus Owen. In: William A. S. Sarjeant (Hrsg.): Vertebrate fossils and the evolution of scientific concepts. Writings in tribute to Beverly Halstead, by some of his many friends. Gordon and Breach, Singapore u. a. 1995, ISBN 2-88124-996-5, S. 229–339.
- P. Martin Sander, Nicole Klein: Developmental Plasticity in the Life History of a Prosauropod Dinosaur. In: Science. Band 310, Nr. 5755, 2005, S. 1800–1802, doi:10.1126/science.1120125.
- Robert R. Reisz, David C. Evans, Hans-Dieter Sues, Diane Scott: Embryonic Skeletal Anatomy of the Sauropodomorph Dinosaur Massospondylus from the Lower Jurassic of South Africa. In: Journal of Vertebrate Paleontology. Band 30, Nr. 6, 2010, S. 1653–1665, doi:10.1080/02724634.2010.521604.
- Robert R. Reisz, David C. Evans, Eric M. Roberts, Hans-Dieter Sues, Adam M. Yates: Oldest known dinosaurian nesting site and reproductive biology of the Early Jurassic sauropodomorph Massospondylus. In: Proceedings of the National Academy of Sciences of the United States of America. Band 109, Nr. 7, 2012, S. 2428–2433, doi:10.1073/pnas.1109385109.
- Robert R. Reisz, Diane Scott, Hans-Dieter Sues, David C. Evans, Michael A. Raath: Embryos of an Early Jurassic prosauropod dinosaur and their evolutionary significance. In: Science. Band 309, Nr. 5735, 2005, S. 761–764, doi:10.1126/science.1114942.