Heterodontosaurus
Heterodontosaurus („Echse mit verschiedenartigen Zähnen“) ist eine Gattung der Vogelbeckensaurier (Ornithischia) und der namensgebende Vertreter der Familie Heterodontosauridae. Die Gattung lebte während des Unterjura (Hettangium bis Sinemurium, vor etwa 200 bis 190 Millionen Jahren) im heutigen Südafrika. Fossilfunde stammen aus der Oberen Elliot-Formation und der Clarens-Formation, deren Ablagerungen auf ein wüstenartiges Klima zu Lebzeiten von Heterodontosaurus deuten. Heterodontosaurus wurde 1962 von Alfred W. Crompton und Alan Charig mit der einzigen Art H. tucki erstmals wissenschaftlich beschrieben.
| Heterodontosaurus | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|

Lebendrekonstruktion und Schädel (aus Sereno, 2012) | ||||||||||
| Zeitliches Auftreten | ||||||||||
| Unterjura (Hettangium bis Sinemurium)[1] | ||||||||||
| 201,3 bis 190,8 Mio. Jahre | ||||||||||
| Fundorte | ||||||||||
| ||||||||||
| Systematik | ||||||||||
| ||||||||||
| Wissenschaftlicher Name | ||||||||||
| Heterodontosaurus | ||||||||||
| Crompton & Charig, 1962 | ||||||||||
| Art | ||||||||||
| ||||||||||
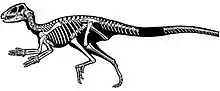
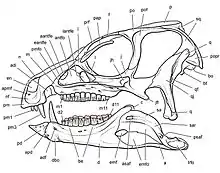
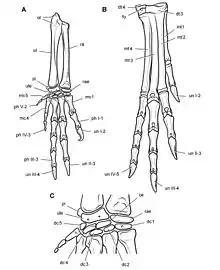
Es handelte sich um einen kleinen, zweibeinig laufenden Pflanzen- oder Allesfresser mit einer geschätzten Länge von 1 bis 1,75 Metern. Namensgebend ist das charakteristische heterodonte Gebiss, das verschiedene Zahntypen umfasst, einschließlich eines Paars hauerartiger „Eckzähne“ und meißelförmiger Backenzähne. Verschiedene Zahntypen, obwohl typisch für Säugetiere, sind unter Reptilien ungewöhnlich. Das Fehlen eines kontinuierlichen Zahnwechsels, ein für Reptilien ebenfalls sehr ungewöhnliches Merkmal, verleitete frühe Studien zu der Annahme, die Tiere hätten eine Sommerruhe gehalten, in welcher sämtliche Zähne auf einmal gewechselt wurden. Weitere Studien befassten sich mit der Frage, ob die charakteristischen Hauer des Gebisses bei beiden Geschlechtern oder nur bei Männchen vorhanden waren (Sexualdimorphismus), ob die Tiere zweibeinig oder vierbeinig liefen, und ob es sich um Pflanzenfresser oder Allesfresser handelte, die auch Fleisch nicht verschmähten.
Merkmale
Heterodontosaurus war ein graziler, zweibeiniger Vogelbeckensaurier aus der Gruppe der Heterodontosauridae.[2] Diese Gruppe umfasste einige der kleinsten bekannten Vogelbeckensaurier;[3] beispielsweise wird Fruitadens auf eine Körperlänge von nur 65 bis 75 cm geschätzt.[2] Heterodontosaurus ist einer der größten Heterodontosauriden:[4] Die Körperlänge wird auf 1 bis 1,75 Meter geschätzt, das Gewicht lag vermutlich zwischen 1,8[5] und 10 Kilogramm.[6] Lediglich Lycorhinus könnte noch größer gewesen sein.[7][4]
Schädel und Gebiss
Der Schädel ist robust gebaut und erscheint in Seitenansicht dreieckig.[8] Die Schnauze endete zu Lebzeiten in einem zahnlosen „Schnabel“ aus Horn. Der obere Teil des „Schnabels“ saß auf dem Prämaxillare, einem dem Hauptknochen des Oberkiefers (Maxillare) üblicherweise vorgelagerten Knochen, während der untere Schnabel dem Prädentale aufsaß, dem vordersten Unterkieferknochen speziell der Vogelbeckendinosaurier. Die Augenhöhlen sind verhältnismäßig groß und annähernd rund, während die externen Nasenöffnungen klein sind. Ein großes Palpebrale, ein für Vogelbeckensaurier typischer spornartiger Knochen, ragt von vorne in die Augenhöhle hinein.[8] Unterhalb der Augenhöhle bildet das Jochbein einen hornartigen, zur Seite gerichteten Fortsatz aus. Zwischen Augenhöhle und externer Nasenöffnung befindet sich ein ausgedehntes Antorbitalfenster, das jedoch eher als Mulde in der Schädelseitenwand erscheint und deshalb auch Antorbitalfossa genannt wird. Diese Mulde beherbergt zwei kleinere Öffnungen, die, je nach Autor, als Foramina oder Fenestrae bezeichnet werden.[8][3] Der untere Rand der Antorbitalgrube wird durch einen markanten Knochengrat gebildet, der zugleich die obere Begrenzung der nach innen (mediad) zurückweichenden unteren, die „Backenzähne“ tragenden Partie des Maxillare (engl. cheek recess) ist.[8] Das Zurückweichen der zahntragenden Partie des Maxillare ist typisch für Vogelbeckendinosaurier und gilt als Hinweis darauf, dass diese Tiere, analog zu Säugetieren, Backentaschen und folglich relativ kleine Mundöffnungen besaßen. Hinter der Augenöffnung liegt das verhältnismäßig große, annähernd eiförmige, schräggestellte Untere Temporalfenster[8] Das elliptisch geformte Obere Temporalfenster ist in Seitenansicht nicht erkennbar. Die Oberen Temporalfenster beider Schädelhälften sind durch einen ausgeprägten Scheitelkamm voneinander getrennt, der Ansatzflächen für die Kiefermuskulatur bot.[8][9]
Das markanteste Merkmal ist die starke Differenzierung des Gebisses die deutlich unterschiedlichen Zahntypen (Heterodontie). Während der vordere Abschnitt des Prämaxillare zahnlos ist und einen hornigen Schnabel trug, sitzen im hinteren Abschnitt des Knochens drei stiftförmige Zähne, von denen der dritte als stark vergrößerter „Hauer“ (englisch tusk) ausgebildet ist. Der Hauer ist durch eine breite Zahnlücke (Diastema) von den sehr engständigen meißelförmigen „Backenzähnen“ des Maxillare getrennt. Das Hauerpaar des Unterkiefers ist deutlich größer als das obere Hauerpaar und griff beim Kieferschluss in eine Einbuchtung im Bereich der Zahnlücke des Maxillare.[3]
Postkraniales Skelett
Der Hals setzte sich aus neun Halswirbeln zusammen und war S-förmig gekrümmt, worauf die Form der Wirbelkörper in Seitenansicht hinweist: So waren die vorderen Wirbelkörper wie ein Parallelogramm geformt, während die der mittleren Halswirbel rechteckig und jene der hinteren Halswirbel trapezförmig waren.[10][11] Der Rumpf war relativ kurz und bestand aus 12 Rückenwirbeln, an denen sich im Beckenbereich 6 miteinander verschmolzene Kreuzbeinwirbel anschlossen.[10] Der proportional lange Schwanz ist nicht vollständig überliefert, bestand aber wahrscheinlich aus 34 bis 37 Schwanzwirbeln.[10] Die Rückenwirbelsäule war ab dem vierten Rückenwirbel durch verknöcherte Sehnen versteift. Dieses Merkmal findet sich bei vielen Vogelbeckensauriern und wirkte vermutlich Biegekräften entgegen, die während der zweibeinigen Fortbewegung die Wirbelsäule belasteten. Anders als bei anderen Vogelbeckensauriern zeigte der Schwanz von Heterodontosaurus keine verknöcherten Sehnen und war somit vermutlich flexibel.[12]
Die Vordergliedmaßen waren kräftig gebaut[13] und mit über 70 % der Länge der Hintergliedmaßen verhältnismäßig lang.[10] Die Speiche des Unterarms maß 70 % der Länge des Oberarmknochens, während die verhältnismäßig große Hand fast ebenso lang wie der Oberarmknochen war.[10] Die Hand endete in fünf, zum Greifen geeigneten Fingern.[13] Der zweite Finger war der längste der Hand, gefolgt vom dritten und ersten Finger (Daumen).[10] Der vierte und fünfte Finger hingegen waren stark zurückgebildet und möglicherweise funktionslos.[14] Die ersten drei Finger endeten in großen und kräftigen Krallen. Die Phalangenformel, welche die Anzahl der Fingerknochen jedes Fingers angibt, betrug 2-3-4-3-2.[10]
Die Hintergliedmaßen waren lang und grazil und endeten in vier Zehen, von denen jedoch nur die zweite, dritte und vierte den Boden berührte.[10] Ein für Vogelbeckensaurier einzigartiges Merkmal war die Verschmelzung verschiedener Bein- und Fußknochen: Schienbein und Wadenbein waren mit den oberen Fußwurzelknochen (Sprungbein und Fersenbein) zu einem Tibiotarsus verschmolzen, während die unteren Fußwurzelknochen mit den Mittelfußknochen zu einem Tarsometatarsus verschmolzen waren.[15] Diese Konstellation findet sich auch bei heutigen Vögeln und hat sich bei diesen und Heterodontosaurus unabhängig voneinander entwickelt.[16] Der Tibiotarsus war um 30 % länger als der Oberschenkelknochen.[10]
Systematik
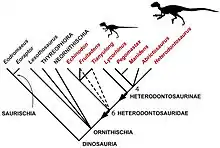
Heterodontosaurus ist die namensgebende und am besten bekannte Gattung der Heterodontosauridae, einer Gruppe sehr kleiner, zweibeiniger Vogelbeckensaurier (Ornithischia). Ein Großteil der Funde stammt aus dem Unterjura Südafrikas; jüngere Funde belegen aber eine deutlich breitere geographische und zeitliche Verbreitung. So sind Heterodontosauriden mittlerweile aus Nordamerika, Südamerika, Europa und Asien nachgewiesen; die Gruppe existierte vermutlich von der Obertrias bis zur Unterkreide über einen Zeitraum von annähernd 100 Millionen Jahren.[6][17] Die systematische Position der Gruppe innerhalb der Vogelbeckensaurier ist umstritten. So weisen sie beispielsweise zahlreiche ursprüngliche Merkmale auf, sodass einige Studien in der Vergangenheit die Heterodontosauriden an die Basis der Vogelbeckensaurier stellten, was auch besser zum vergleichsweise hohen geologischen Alter ihrer Vertreter passt[18] (stärker abgeleitete Ornithischier sind eher typisch für die Kreidezeit als für den Jura).Die meisten Studien legen eine enge Verwandtschaft mit den Ornithopoda (Hadrosauridae und Verwandte) oder den Marginocephalia (gehörnte und Dickschädel-Dinosaurier[19]) nahe. Nach neuesten Studien werden die Heterodontosauridae als Vertreter der Cerapoda und Vorfahren der Pachycephalosauria betrachtet, was sie zu den Ältesten bekannten Vertretern der Cerapoda macht. In derselben Studie wurden die Heterodontosauridae, die Familie, zu der Heterodontosaurus gehört, als paraphyletisch bezeichnet, was bedeutet, dass die Vertreter der Heterodontosauridae zwar einen gemeinsamen Vorfahren haben, aber nicht alle Nachkommen dieses gemeinsamen Vorfahren einschließen.[19]
Über die Verwandtschaftsbeziehungen zwischen Heterodontosaurus und anderen Heterodontosauriden ist wenig bekannt, da die meisten bekannten Vertreter nur sehr fragmentarisch überliefert sind.[18] Beispielsweise können Feng-Lu Han und Kollegen in ihrer phylogenetischen Analyse die innere Systematik der Gruppe nicht auflösen;[20] einige andere Studien erreichen eine Auflösung nur durch das Entfernen einiger der nur fragmentarisch überlieferten Vertreter.[18] Einige Analysen betrachten Heterodontosaurus als nächstverwandt mit dem ebenfalls aus Südafrika stammenden Lycorhinus.[18] Außerdem kommen verschiedene Analysen zu dem Ergebnis, dass Fruitadens und Tianyulong näher miteinander verwandt waren als mit anderen Gattungen.[18][21] Paul Sereno (2012) fasst die südafrikanischen Vertreter (Heterodontosaurus, Lycorhinus, Pegomastax, Abrictosaurus) sowie den südamerikanischen Manidens als Heterodontosaurinae zusammen, während er die aus den nördlichen Landmassen (Laurasia) stammenden Heterodontosauriden Echinodon, Fruitadens und Tianyulong außerhalb dieser Gruppe klassifiziert, wobei er nicht auflösen kann, ob diese Gattungen ihrerseits eine Klade bilden.[18]
Entdeckungsgeschichte
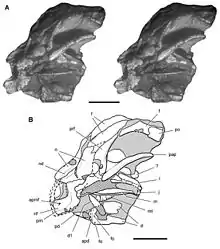
Das erste Heterodontosaurus-Fossil wurde bereits Anfang des 20. Jahrhunderts von Robert Broom entdeckt, allerdings erst kürzlich als solches erkannt. Broom verkaufte das Fossil 1913 zusammen mit zahlreichen anderen südafrikanischen Fossilien an das American Museum of Natural History in New York, wo es seitdem zusammen mit den Synapsiden-Fossilien der Broom-Sammlung archiviert wurde (Exemplarnummer AMNH 24000). Erst ein knappes Jahrhundert später wurde Paul Sereno bei der Durchsicht der Sammlung auf das Fossil aufmerksam, das noch immer größtenteils in einem Gesteinsblock eingeschlossen war. Die anschließende Präparation brachte das Schädelfragment eines noch nicht erwachsenen Heterodontosaurus zum Vorschein. Hinter dem Schädel fanden sich zudem Teile mehrerer Halswirbel, was darauf hindeutet, dass möglicherweise auch Teile des restlichen Skeletts erhalten waren, die jedoch an der Fundstelle im Gestein verblieben. Wie Sereno angibt, stammt das Fossil wahrscheinlich aus der Clarens-Formation. 2012 wurde der Fund erstmals publiziert.[22]
Anfang der 1960er Jahre tauchte schließlich ein teilweises Skelett mit einem nahezu vollständigen Schädel in der Transkei unmittelbar südlich von Lesotho auf. Der Fund (Holotyp, SAM-PK-K337) stammt aus der Clarens-Formation und wurde in den Jahren 1961–1962 von Alfred W. Crompton während einer gemeinsamen paläontologischen Expedition des Iziko South African Museum und des British Museum gemacht; der Fundort ist heute als Tyinindini-Lokalität bekannt. Die von Arthur E. Rixon durchgeführte Präparation des Schädels gestaltete sich schwierig, da ein dünner aber sehr harter Überzug aus Hämatit die Anwendung einer Diamantsäge erforderte; beim Sägen wurde der Schädel leicht beschädigt.[23][24][25] Bereits 1962 veröffentlichten Crompton und Alan Charig eine vorläufige Beschreibung des Schädels in der Fachzeitschrift Nature, und benannten Heterodontosaurus tucki als neue Art und Gattung.[24] Dabei weist der Name Heterodontosaurus (gr. hetero – „anders“; odous – „Zahn“; sauros – „Echse“)[26] auf die für einen Vogelbeckensaurier ungewöhnliche heterodonte Bezahnung, während der Name tucki G. C. Tuck ehrt, den Manager des südafrikanischen Ablegers der Austin Motor Company, welcher die Expedition unterstützte.[24] 2011 veröffentlichten David Norman und Kollegen eine umfangreichere Beschreibung des Schädels;[27] das Restskelett wurde jedoch nie beschrieben und scheint heute verloren zu sein.[23][25] Der Schädel wird heute im Iziko South African Museum in Kapstadt aufbewahrt.[25]

Dasselbe Fundgebiet wurde im Sommer 1966–1967 von einer weiteren gemeinsamen Expedition untersucht; dieses Mal waren neben dem Iziko South African Museum und dem British Museum auch die Yale University und das University College London beteiligt. Aus einer einzigen, als Voyizane bezeichneten Fundlokalität in der Oberen Elliot-Formation kamen fünf Heterodontosaurus-Exemplare zum Vorschein, darunter der bislang vollständigste Fund, ein nahezu komplettes Skelett mit Schädel (SAM-PK-K1332). Dieses Skelett wurde ebenfalls von Crompton entdeckt und fand sich eingebettet in roten Sandsteinen. Der Schädel, der durch Ione Rudner im Iziko South African Museum präpariert wurde, ist besser erhalten als der Holotyp-Schädel und war nicht wie letzterer mit Hämatit bedeckt. Zahlreiche kleine Risse wurden allerdings mit Klebstoff ausgebessert, und die Oberflächen von Knochen und Zähne wurden mit einer Schicht aus Klebstoff stabilisiert, die teilweise feinere anatomische Details unkenntlich macht.[25] Die übrigen Funde aus Voyizane umfassen den vorderen Abschnitt des Schädels eines Jungtiers (SAM-PK-K10487); zwei Oberkiefer-Fragmente (SAM-PK-K1326 und SAM-PK-K1334); sowie ein fragmentarisches, schädelloses Skelett inklusive Teilen der Wirbelsäule, des Beckengürtels und der Vorder- und Hinterextremitäten (SAM-PK-K1328).[23]
Ein relativ großes Schnauzenfragment (NM QR 1788) wurde 1975 auf der Tushielaw-Farm entdeckt, etwa 60 km östlich von Voyizane. Dieses Fossil wurde lange dem Prosauropoden Massospondylus zugeschrieben, bis Porro und Kollegen 2011 zeigten, dass es sich bei dem im National Museum von Südafrika aufbewahrten Exemplar tatsächlich um ein weiteres Heterodontosaurus-Fossil handelt.[23]
Paläohabitat
Heterodontosaurus-Fossilien stammen aus der Oberen Elliot-Formation und der darüberliegenden Clarens-Formation.[1] Dieses Schichtintervall wird auch als Massospondylus range zone bezeichnet, benannt nach dem prosauropoden Dinosaurier Massospondylus, welcher das häufigste Fossil dieser Schichten darstellt.[28] Die Obere Elliot-Formation, welche den Großteil der Fossilien barg, wird aus rötlichen, fluviatil (durch Flüsse) bis äolisch (durch Wind) abgelagerten Sandsteinen aufgebaut. Die etwas jüngere Clarens-Formation enthielt unter anderem das Holotyp-Exemplar von Heterodontosaurus.[29] Die durch creme-farbene Sedimente aufgebaute Formation lagerte teils äolisch, teils innerhalb eines Playasees ab. Die Clarens-Formation ist weniger reich an Fossilien als die Obere Elliot-Formation; zudem bildet sie bevorzugt Kliffs aus, was ihre Zugänglichkeit erschwert.[30]
Heterodontosaurus lebte vermutlich in einem trockenen, wüstenartigen Klima.[31] Aus der Oberen Elliot-Formation sind drei weitere Heterodontosauriden bekannt – Abrictosaurus, Lycorhinus und Pegomastax – damit stellt diese Formation die weltweit größte bekannte Vielfalt dieser Gruppe. In der Clarens-Formation kommt Heterodontosaurus zusammen mit dem Heterodontosauriden Geranosaurus vor.[30] Neben Heterodontosauriden wurden andere Dinosaurier (unter anderem Lesothosaurus, Megapnosaurus und Massospondylus), temnospondyle Amphibien, Schildkröten, Sphenodontier, Rauisuchier, Krokodilverwandte, sowie verschiedene Cynodontier (Säugetier-Vorläufer) gefunden.[28]
Paläobiologie
Ontogenese
Über die individuelle Entwicklungsgeschichte (Ontogenese) von Heterodontosaurus ist wenig bekannt. Der 2006 beschriebene Schädel eines Jungtiers (SAM-PK-K10487) erlaubt erstmals Einblicke in anatomische Veränderungen, die in der Entwicklung vom Jungtier zum ausgewachsenen Tier stattfanden: So werden mit zunehmendem Alter die Augenhöhlen proportional kleiner, während der Abschnitt des Schädels vor den Augenhöhlen länglicher wird. Außerdem verschmelzen Teile des Nasenbeins und des Zwischenkieferbeins. Da es, abgesehen von der Zunahme der Anzahl der Zähne im Oberkiefer, keine Änderungen im Gebiss gibt, wird vermutet, dass sich die Ernährung des Tieres während des Wachstums nicht änderte.[32] Die Gesamtlänge des Schädels wird auf 45 mm rekonstruiert. Hätte das Jungtier dieselben Körperproportionen gezeigt wie ein erwachsenes Tier, würde die Körperlänge 45 cm betragen haben – tatsächlich war das Tier vermutlich kleiner, da Schädel bei Jungtieren generell proportional größer sind als bei erwachsenen Tieren (Kindchenschema).[33]
Sexualdimorphismus
Richard Thulborn vermutete 1974, dass es sich bei den vergrößerten Hauern der Heterodontosauride um ein sekundäres Geschlechtsmerkmal handelte (Sexualdimorphismus). So hätten nur männliche Exemplare Hauer besessen, während es sich bei hauerlosen Exemplaren, wie dem Typexemplar von Abrictosaurus, um Weibchen gehandelt hätte.[34] Richard Butler und Kollegen (2006) zogen die Hypothese jüngst in Zweifel: So zeigt der von ihnen beschriebene, von einem Jungtier stammende Schädel, dass Hauer bereits in einem frühen Entwicklungsstadium ausgeprägt waren – geschlechtsspezifische Unterschiede seien in einem derart frühen Stadium noch nicht zu erwarten. Außerdem seien Hauer in nahezu allen bekannten Schädeln vorhanden; bei einem echten Sexualdimorphismus dagegen sei ein 50:50-Verhältnis zwischen hauertragenden und hauerlosen Exemplaren zu erwarten. Eine Ausnahme bilde lediglich das Typexemplar von Abrictosaurus – das Fehlen der Hauer stellt jedoch vermutlich eine Spezialisierung dieser Gattung dar.[32]
Sommerruhe und Zahnwechsel
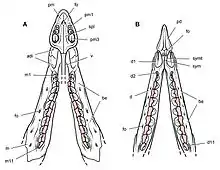
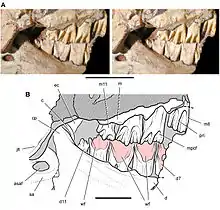
Thulborn (1974, 1978) prägte die Hypothese, dass Heterodontosauriden die trockene und nahrungsarme Jahreszeit in einer Sommerruhe verbrachten. Die Hypothese fußte auf Beobachtungen des Gebisses von Heterodontosaurus: So finden sich keine Hinweise auf einen kontinuierlichen Zahnwechsel, wie er für Dinosaurier und andere diapside Reptilien[27] typisch ist. Die Backenzähne waren einheitlich abgenutzt, ohne dass frisch durchgebrochene Zähne vorhanden waren. Die mutmaßlich widerstandsfähige Pflanzennahrung hätte laut Thulborn jedoch zu einem schnellen Abnutzen der Zähne geführt, was einen regelmäßigen Wechsel der Bezahnung unerlässlich macht. Ein Wechsel der gesamten Bezahnung könne nur in den Phasen der Sommerruhe stattgefunden haben, wo die Tiere keine Nahrung aufnahmen. Als weiteres Argument für die Sommerruhen-Hypothese führt Thulborn die Abnutzungs-Fazetten der Zähne auf, die durch Kontakt der oberen mit der unteren Bezahnung entstanden. Bei dem zeitgenössischen Ornithischia Fabrosaurus fänden sich an jedem Zahn eine vordere und eine hintere Abnutzungsfazette, die durch das Ineinandergreifen der oberen und unteren Zähne entstanden. Die Kieferbewegung bei Fabrosaurus sei damit ausschließlich vertikal gewesen. Bei Heterodontosauriden dagegen gingen die Abnutzungs-Fazetten ineinander über und bilden eine Bahn über die gesamte Zahnreihe. Dies weise darauf hin, dass die Tiere ihre Nahrung nicht mit Vertikalbewegungen, sondern mit Vor-Zurück-Bewegungen der Kiefer zerkleinerten. Diese Vor-Zurück-Bewegungen seien nur möglich, wenn die Zähne hinsichtlich ihres Abnutzungsgrads und ihrer Größe einheitlich sind; diese Voraussetzung sei bei einem kontinuierlichen Zahnwechsel nicht gegeben. Dies sei ein Hinweis darauf, dass die gesamte Bezahnung periodisch innerhalb der Sommerruhe als eine Einheit gewechselt wurde.[34][35][36]
Eine ausführliche Analyse von James Hopson (1980) stellte Thulborns Ideen in Frage. Hopson zeigte, dass das Muster der Abnutzungs-Fazetten tatsächlich für eine vertikale und seitliche Kieferbewegung spricht, nicht aber für eine Vor-Zurück-Kieferbewegung. Außerdem würde der Abnutzungsgrad der einzelnen Zähne variieren, was einen kontinuierlichen Zahnwechsel anzeigt. Allerdings zeigten Röntgenaufnahmen, dass in den Kiefern des vollständigsten Heterodontosaurus-Skeletts tatsächlich keine Zähne neu gebildet wurden. Hopson folgerte, dass Jungtiere einen kontinuierlichen Zahnwechsel zeigten, welcher bei ausgewachsenen Tieren jedoch zum Stillstand kam. Hopson resümiert, dass für Thulborns Sommerruhe-Hypothese keine Nachweise existieren.[36]
Butler und Kollegen (2008) führten schließlich Computertomographie-Scans an einem juvenilen Schädel durch, die, zur Überraschung dieser Forscher, keine sich in Bildung befindlichen Zähne aufweisen. Die Forscher führen aus, dass es in der Individualentwicklung der Tiere dennoch Zahnwechsel gegeben haben muss: So zeigen die Zähne des Jungtier-Schädels dieselbe Morphologie wie jene ausgewachsener Tiere – diese Merkmale wären nicht erhalten, wäre der Zahn schlicht kontinuierlich gewachsen. Der Zahnwechsel bei Heterodontosaurus sei sporadischer gewesen als bei verwandten Gattungen.[32] Noch nicht durchgebrochene Ersatzzähne konnten erst in einem von Norman und Kollegen (2011) beschriebenen Oberkiefer (SAM-PK-K1334) sowie in einem von Paul Sereno (2012) beschriebenen juvenilen Schädel (AMNH 24000) nachgewiesen werden. Wie diese Funde zeigen, war der Zahnwechsel episodisch und nicht kontinuierlich wie bei anderen Heterodontosauriden.[37][38] Die Ersatzzähne zeigten die für ursprüngliche Ornithischia typischen in Seitenansicht dreieckigen Zahnkronen. Die durchgebrochenen Backenzähne haben ihre charakteristische meißelartige Form folglich erst durch gegenseitige Abnutzung der oberen und unteren Bezahnung erhalten.[37]
Fortbewegung und Stoffwechsel
Obwohl die meisten Studien Heterodontosaurus als zweibeinigen (bipeden) Läufer darstellen, vermuteten einige Autoren eine teilweise oder vollständige vierbeinige (quadrupede) Fortbewegung. Santa Luca (1980) beschrieb verschiedene Merkmale des Armskeletts, die sich auch bei quadrupeden Tieren finden und eine kräftige Armmuskulatur anzeigen: So war das Olekranon der Elle relativ groß, was den Hebelarm des Unterarms vergrößert. Der große mediale Epikondylus des Oberarmknochens bot eine Ansatzstelle für kräftige Beugermuskeln des Unterarms, während Fortsätze an den Krallen zeigen, dass die Hand zu einem kräftigen nach vorne gerichteten Schub beim Gehen fähig war. Heterodontosaurus hätte sich beim langsamen Gehen vierbeinig fortbewegt, wäre zum Laufen jedoch in eine zweibeinige Fortbewegung gewechselt.[12] Teresa Maryańska und Halszka Osmólska (1985) vermuteten ebenfalls eine quadrupede Fortbewegung; als weiteres Argument führten sie die stark nach unten gekrümmte Rückenwirbelsäule in dem vollständigsten bekannten Exemplar auf.[39] Gregory Paul (1987) vermutete dagegen, dass sich diese Tiere grundsätzlich quadruped fortbewegten – höhere Geschwindigkeiten seien durch Galoppieren erreicht worden.[40] Allerdings gibt Paul keine anatomischen Merkmale zur Unterstützung dieser Hypothese an. David Weishampel und Lawrence Witmer (1990) sowie Norman und Kollegen (2004) argumentierten für eine ausschließlich bipede Fortbewegung, basierend auf der Form der Krallen und der Anatomie des Schultergürtels.[41][42] Die von Santa Luca beschriebenen Merkmale seien Anpassungen zur Nahrungssuche – so könnten die kräftigen Arme zum Ausgraben von Wurzeln oder zum Aufbrechen von Insektennestern gedient haben.[41]
Die meisten jüngeren Studien betrachten Dinosaurier als endotherme (warmblütige) Tiere mit einem Stoffwechsel, der mit dem heutiger Säugetiere und Vögel vergleichbar ist. Pontzer und Kollegen (2009) berechneten die zur Fortbewegung nötige aerobe Ausdauer bei verschiedenen Dinosauriern und konnten daraus auf Endothermie bei den meisten untersuchten Gattungen schließen. Heterodontosaurus hätte bereits bei moderaten Laufgeschwindigkeiten die maximale aerobe Ausdauer überschritten, die einem gleich großen ektothermen Tier zur Verfügung stünde.[43]
Ernährung und Funktion der Hauer
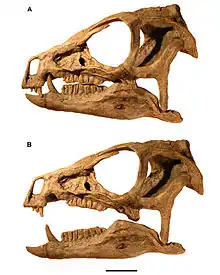
Ein Großteil der veröffentlichten Studien betrachtet Heterodontosaurus als einen Pflanzenfresser. Wie Molnar (1977) anmerkte, hätten die Hauer keine Funktion in der Ernährung der Tiere gehabt, sondern seien als geschlechtsspezifisches Merkmal möglicherweise in innerartlichen Kämpfen und zur Zurschaustellung verwendet worden. Eine ähnliche Funktion fände sich bei den verlängerten Eckzähnen der heutigen Muntjaks.[13]
Verschiedene jüngere Studien warfen allerdings die Möglichkeit auf, dass die Hauer von Heterodontosaurus tatsächlich zum gelegentlichen Reißen von Beutetieren eingesetzt worden sein könnten.[44][32][45][2] So zeigt Paul Barrett (2000), dass die Schneidekanten der Hauer mit einer feinen Zähnelung (Serration) versehen seien, ähnlich jener, wie sie sich bei Theropoden (fleischfressende Dinosaurier) findet. Bei Muntjaks dagegen sei keine Zähnelung vorhanden. Ein weiterer Hinweis auf eine fakultativ allesfressende Ernährungsweise sei die Form der beiden vor dem oberen Hauer sitzenden Zähne.[44] Richard Butler und Kollegen (2008) argumentieren, dass die Hauer kein geschlechtsspezifisches Merkmal seien und sich schon früh in der Entwicklung der Tiere herausbildeten – somit erschiene es wahrscheinlicher, dass die Hauer in einer allesfressenden Ernährungsweise oder zur Verteidigung eingesetzt wurden als in innerartlichem Konkurrenzverhalten.[32] Forscher um David Norman (2011) betonen, dass die Arme und Hände verhältnismäßig lang und mit großen, gebogenen Krallen ausgestattet gewesen seien, was das Packen kleiner Beutetiere ermöglichte. Dagegen seien die Hinterbeine lang gewesen und hätten ein schnelles Laufen ermöglicht. Als Allesfresser hätte Heterodontosaurus während der vegetationsarmen Trockenzeiten einen deutlichen Selektionsvorteil gehabt.[45][13]
Paul Sereno (2012) dagegen führt verschiedene Schädel- und Gebissmerkmale auf, die laut diesem Forscher für eine reine oder überwiegend pflanzenfressende Ernährungsweise bei Heterodontosauriden sprechen. So besäßen diese Tiere einen zum Abschneiden geeigneten Schnabel, spezialisierte Backenzähne mit Schneidefunktion, sowie Wangen, welche der oralen Verarbeitung der Nahrung dienten. Die Schließmuskulatur der Kiefer sei vergrößert gewesen, während das Kiefergelenk relativ zur Zahnreihe tiefer gelegt war, was die Effektivität der Kiefermuskeln bei der Verarbeitung von Pflanzenmaterial vergrößerte. Schließlich seien die Unterschiede in Größe und Position der Hauer bei verschiedenen Heterodontosauriden zu groß, als dass sie eine spezifische Funktion in der Ernährung gehabt haben könnten. Sereno vermutet, dass Heterodontosauriden am ehesten mit heutigen Nabelschweinen zu vergleichen seien, welche ähnliche Hauer zeigen und sich von einer Vielzahl verschiedenen Pflanzenmaterials ernähren, darunter Wurzeln, Knollen, Früchte und bodennahe Vegetation.[13]
Literatur
- Richard J. Butler, Laura B. Porro, David B. Norman: A Juvenile Skull of the Primitive Ornithischian Dinosaur Heterodontosaurus tucki from the ‚Stormberg‘ of Southern Africa. In: Journal of Vertebrate Paleontology. Bd. 28, Nr. 3, 2008, ISSN 0272-4634, S. 702–711, doi:10.1671/0272-4634(2008)28[702:AJSOTP]2.0.CO;2, Digitalisat (PDF; 890,38 kB).
- Richard J. Butler, Jin Liyong, Chen Jun, Pascal Godefroit: The postcranial osteology and phylogenetic position of the small ornithischian dinosaur Changchunsaurus parvus from the Quantou Formation (Cretaceous: Aptian–Cenomanian) of Jilin Province, north-eastern China. In: Palaeontology. Bd. 54, Nr. 3, 2011, ISSN 0031-0239, S. 667–683, doi:10.1111/j.1475-4983.2011.01046.x.
- Alfred W. Crompton, Alan J. Charig: A new ornithischian from the Upper Triassic of South Africa. In: Nature. Bd. 196, Nr. 4859, 1962, S. 1074–1077, doi:10.1038/1961074a0.
- James A. Hopson: Tooth function and replacement in early Mesozoic ornithischian dinosaurs: implications for aestivation. In: Lethaia. Bd. 13, Nr. 1, 1980, ISSN 0024-1164, S. 93–105, doi:10.1111/j.1502-3931.1980.tb01035.x.
- Albert P. Santa Luca: The postcranial skeleton of Heterodontosaurus tucki (Reptilia, Ornithischia) from the Stormberg of South Africa. In: Annals of the South African Museum. Bd. 79, Nr. 7, 1980, ISSN 0303-2515, S. 159–211, Digitalisat.
- David B. Norman, Hans-Dieter Sues, Lawrence M. Witmer, Rodolfo A. Coria: Basal Ornithopoda. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2. Ausgabe. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 393–412, Digitalisat (PDF; 2,66 MB).
- David B. Norman, Alfred W. Crompton, Richard J. Butler, Laura B. Porro, Alan J. Charig: The Lower Jurassic ornithischian dinosaur Heterodontosaurus tucki Crompton & Charig, 1962: cranial anatomy, functional morphology, taxonomy, and relationships. In: Zoological Journal of the Linnean Society. Bd. 163, Nr. 1, 2011, ISSN 0024-4082, S. 182–276, doi:10.1111/j.1096-3642.2011.00697.x.
- Paul C. Sereno: Taxonomy, morphology, masticatory function and phylogeny of heterodontosaurid dinosaurs (= ZooKeys. Nr. 226, Special ssue). Pensoft, Sofia 2012, ISBN 978-954-642-652-9, S. 1–2, 30, doi:10.3897/zookeys.226.2840.
- Richard A. Thulborn: A new heterodontosaurid dinosaur (Reptilia: Ornithischia) from the Upper Triassic Red Beds of Lesotho. In: Zoological Journal of the Linnean Society. Bd. 55, Nr. 2, 1974, S. 151–175, doi:10.1111/j.1096-3642.1974.tb01591.x.
- Richard A. Thulborn: Aestivation among ornithopod dinosaurs of the African Trias. In: Lethaia. Bd. 11, Nr. 3, 1978, S. 185–198, doi:10.1111/j.1502-3931.1978.tb01226.x.
- David B. Weishampel, Lawrence M. Witmer: Heterodontosauridae. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. University of California Press, Berkeley CA u. a. 1990, ISBN 0-520-06726-6, S. 486–497.
Weblinks
Einzelnachweise
- Sereno 2012 S. 85
- Richard J. Butler, Peter M. Galton, Laura B. Porro, Luis M. Chiappe, Donald M. Henderson, Gregory M. Erickson: Lower limits of ornithischian dinosaur body size inferred from a new Upper Jurassic heterodontosaurid from North America. In: Proceedings of the Royal Society. Series B: Biological Sciences. Bd. 277, Nr. 1680, 2010, ISSN 0080-4649, S. 375–381, hier S. 375, 380, doi:10.1098/rspb.2009.1494.
- Weishampel und Witmer 1990, S. 486–491
- Sereno 2012 S. 161
- Frank Seebacher: A new method to calculate allometric length-mass relationships of dinosaurs. In: Journal of Vertebrate Paleontology. Bd. 21, Nr. 1, 2001, S. 51–60, hier S. 53, doi:10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2, Digitalisat (PDF; 141 kB).
- Richard J. Butler, Laura B. Porro, Peter M. Galton, Luis M. Chiappe: Anatomy and Cranial Functional Morphology of the Small-Bodied Dinosaur Fruitadens haagarorum from the Upper Jurassic of the USA. In: PLoS ONE. Bd. 7, Nr. 4, 2012, S. e31556, doi:10.1371/journal.pone.0031556.
- Christopher E. Gow: A tooth-bearing maxilla referable to Lycorhinus angustidens Haughton, 1924 (Dinosauria, Ornithischia). In: Annals of the South African Museum. Bd. 99, Nr. 10, 1990, S. 367–380, hier S. 379, Digitalisat.
- Norman et al. 2011, S. 194–195
- Norman et al. 2011, S. 227–228
- Weishampel und Witmer 1990, S. 491–494
- Santa Luca 1980 S. 163–166
- Santa Luca 1980 S. 197–198
- Sereno 2012 S. 187–193
- Santa Luca 1980 S. 181
- Norman et al. 2004, S. 408
- Sereno 2012 S. 132
- Sereno 2012 S. 1
- Sereno 2012 S. 193–204
- Beispiel für eine noch junge Studie, nach deren Ergebnis die Heterodontodauriden basale Pachycephalosaurier sind: P.-E. Dieudonné, P. Cruzado-Caballero, P. Godefroit, T. Tortosa: A new phylogeny of cerapodan dinosaurs. In: Historical Biology. 20. Juli 2020, ISSN 0891-2963, doi:10.1080/08912963.2020.1793979. (im Druck)
- Feng-Lu Han, Paul M. Barrett, Richard J. Butler, Xing Xu: Postcranial anatomy of Jeholosaurus shangyuanensis (Dinosauria, Ornithischia) from the Lower Cretaceous Yixian Formation of China. In: Journal of Vertebrate Paleontology. Bd. 32, Nr. 6, 2012, S. 1370–1395, hier S. 1391, doi:10.1080/02724634.2012.694385.
- Richard J. Butler, David B. Norman, Alfred W. Crompton, Laura B. Porro, Alan J. Charig: The postcranial osteology and phylogenetic position of the small ornithischian dinosaur Changchunsaurus parvus from the Quantou Formation (Cretaceous: Aptian–Cenomanian) of Jilin Province, north‐eastern China. In: Palaeontology. Bd. 54, Nr. 3, 2011, S. 667–683, doi:10.1111/j.1475-4983.2011.01046.x.
- Sereno 2012 S. 10
- Sereno 2012 S. 14–17
- Crompton und Charig 1962
- Norman et al. 2011, S. 189
- Donald F. Glut: Dinosaurs. The Encyclopedia. McFarland & Company, Jefferson NC 1997, ISBN 0-89950-917-7, S. 467–469.
- Norman et al. 2011, S. 182
- Fabien Knoll: The tetrapod fauna of the Upper Elliot and Clarens formations in the main Karoo Basin (South Africa and Lesotho). In: Bulletin de la Société Géologique de France. Bd. 176, Nr. 1, 2005, ISSN 0037-9409, S. 81–91, hier S. 81–85, doi:10.2113/176.1.81.
- Butler et al. 2008 S. 702
- Sereno 2012 S. 10–11
- Michael R. Cooper: The prosauropod dinosaur Massospondylus carinatus Owen from Zimbabwe: its biology, mode of life and phylogenetic significance (= Occasional Papers of the National Museums and Monuments of Rhodesia.) Series B: Natural Sciences. Bd. 6, Nr. 10, 1981, ZDB-ID 405377-1, S. 812.
- Butler et al. 2008 S. 709–710
- Butler et al. 2008 S. 704
- Thulborn 1974
- Thulborn 1978
- Hopson 1980
- Norman et al. 2011, S. 220–221
- Sereno 2012 S. 163
- Teresa Maryańska, Halszka Osmólska: On ornithischian phylogeny. In: Acta Palaeontologica Polonica. Bd. 30, Nr. 3/4, 1985, ISSN 0567-7920, S. 137–150, hier S. 147, online.
- Gregory S. Paul: The Science and Art of Restoring the Life Appearance of Dinosaurs and Their Relatives; a Rigorous How-to Guide. In: Sylvia J. Czerkas, Everett C. Olson (Hrsg.): Dinosaurs, Past and Present. Band 2. University of Washington Press u. a., Seattle WA u. a. 1987, ISBN 0-295-96570-3, S. 33.
- Weishampel und Witmer 1990, S. 497
- Norman et al. 2004, S. 412
- Herman Pontzer, Vivian Allen, John R. Hutchinson: Biomechanics of Running Indicates Endothermy in Bipedal Dinosaurs. In: PLoS ONE. Bd. 4, Nr. 11, 2009, e7783, doi:10.1371/journal.pone.0007783.
- Paul M. Barrett: Prosauropod dinosaurs and iguanas: speculations on the diets of extinct reptiles. In: Hans-Dieter Sues: Evolution of herbivory in terrestrial vertebrates. Perspectives from the fossil record. Cambridge University Press, Cambridge u. a. 2000, ISBN 0-521-59449-9, S. 42–78, hier S. 64, doi:10.1017/CBO9780511549717.004.
- Norman et al. 2011, S. 231