Homunculus (Gattung)
Homunculus ist eine ausgestorbene Gattung der Primaten aus der Gruppe der Neuweltaffen. Sie lebte im Übergang vom Unteren zum Mittleren Miozän vor 18 bis 16 Millionen Jahren im südlichen Teil von Südamerika. Alle bisherigen Fossilreste wurden in der Santa-Cruz-Formation in Patagonien geborgen, die Gesteinseinheit stellt die südlichste Fundregion eines Affen auf dem Kontinent dar. Das Material besteht aus Schädeln, Gebissresten und einzelnen Teilen der Gliedmaßen. Sie gehören zu einem mittelgroßen Vertreter der Neuweltaffen, der sich durch einzelne urtümliche Merkmale von den heutigen Arten unterscheidet. Die Tiere wiesen ein verhältnismäßig kleines Gehirn auf, das Gehör war gut entwickelt. Die Männchen und Weibchen bildeten möglicherweise kleine Gruppen. Als Lebensraum dienten subtropische Wälder oder solche, die unter dem Einfluss eines jahreszeitlichen Wechsels standen. In diesen bewegten sich die Tiere kletternd in Bäumen fort und verrichteten ihre Aktivitäten bei Tageslicht. Die Nahrung setzte sich aus harten Samen und weichen Blättern und Früchten zusammen. Die ersten Funde kamen bereits Ende des 19. Jahrhunderts zum Vorschein, kurz danach wurde die Gattung wissenschaftlich eingeführt. Momentan sind zwei Arten anerkannt, das bisher bekannte Fossilmaterial lässt aber weitere vermuten. Zur systematischen Stellung gibt es eine kontroverse Diskussion, die aber auch andere frühe Neuweltaffen mit einbezieht.
| Homunculus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
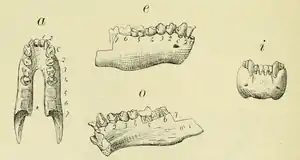
Unterkiefer von Homunculus (Holotyp, verloren gegangen) | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Unteres bis Mittleres Miozän | ||||||||||||
| 17,4 bis 16,3 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Homunculus | ||||||||||||
| Ameghino, 1891 | ||||||||||||
Merkmale
Homunculus war ein mittelgroßer Vertreter der Neuweltaffen, von dem überwiegend Schädelmaterial, aber auch einzelne Teile des Bewegungsapparates bekannt sind. Der Schädel wies eine Länge von rund 6,6 cm und eine Breite von 2,4 bis 2,8 cm auf, die Höhe des Gesichtsbereiches unterhalb der Augen variierte von knapp 1,0 bis 1,2 cm. Das Rostrum war bei Homunculus stärker vorgezogen (prognath) als bei den heutigen Neuweltaffen, was etwa einen ausladenderen Mittelkieferknochen hervorrief. Dagegen ist der Knochen bei den Springaffen und bei den Nachtaffen eher verkürzt. Das Nasenbein zeigte sich bei Homunculus im Bereich der Nasenöffnung sehr weit. Das Stirnbein war nicht aufgewölbt und die Zwischenaugenregion relativ weit. Die Orbita wies etwa die Ausmaße wie bei den heutigen Springaffen auf. Die absoluten Maße betrugen 1,2 bis 1,4 cm sowohl in der Höhe als auch in der Breite. Im Gegensatz zu den Nachtaffen oder zum ausgestorbenen Tremacebus war sie somit nicht vergrößert. Dadurch dehnten sich bei Homunculus auch einige Nasennebenhöhlen wie etwa die Kieferhöhle deutlich aus, die bei den Nachtaffen reduziert sind, da der Platz von den großen Augen eingenommen wird. Der Rand der Orbita war zudem vollständig geschlossen. Am Pterion (eine Region seitlich der Augen beim menschlichen Schädel) trafen sich bei einigen Schädeln von Homunculus das Keilbein und das Stirnbein und bildeten eine gemeinsame Knochennaht. Dies ähnelt eher den Altweltaffen, da bei den heutigen Neuweltaffen hier die Sutur des Scheitelbeins und des Jochbeins verläuft. Andere Schädelfunde wiesen bei Homunculus hier wiederum die Eigenheiten platyrrhiner Schädel auf, so dass das Merkmal insgesamt als eher polymorph eingestuft werden muss.[1] Die Hinterhauptsschuppe des Hinterhauptsbeins stand steil schräg zur Augen-Ohr-Ebene. Dies ist typisch für fast alle Neuweltaffen mit Ausnahme der Totenkopfaffen, da bei diesen die Schuppe sehr spitzwinklig einfällt.[2][3][4][5][6]

Der Unterkiefer von Homunculus war breit, aber weniger tief und robust als bei zahlreichen heutigen blattfressenden Neuweltaffen. Er besaß einen nach hinten tiefer werdenden horizontalen Knochenkörper, in Längsansicht wirkte er etwas gestaucht. In seiner generellen Form bestanden Ähnlichkeiten zum Unterkiefer der heutigen Nachtaffen und zu einigen Springaffen, bei letzteren ist er aber massiver ausgeprägt. Er unterschied sich aber von dem der Sakiaffen, bei denen der horizontale Knochenkörper vorn deutlich höher ist und von dem der Kapuzineraffen mit seiner gestreckteren Form und eher ausgeglichenen Profillinie. Die beiden Äste liefen bei Homunculus V-förmig aufeinander zu und fusionierten in einer geschlossenen, fest verwachsenen Symphyse. Hier neigte sich das Kinn um etwa 50° gegenüber den vorderen Zähnen ab. Das Foramen mentale lag unterhalb des vorderen Prämolaren etwa oberhalb der Knochenmitte. Der aufsteigende Ast war breit, der Winkelfortsatz lud weit nach hinten aus. Der Kronenfortsatz ragte steil auf und war hakenartig leicht nach hinten gebogen. Der Gelenkfortsatz saß tiefer, aber immer noch deutlich über der Kauebene der Zähne. Sein Hinterrand bildete eine steile Kante. Das Gelenkköpfchen war vorn und hinten verschmälert, aber stark gewölbt. An der Außenseite bestand eine nur seichte Fossa masseterica. Innen öffnete sich das Foramen mandibulare am Ende einer scharf begrenzten Rinne etwa auf Höhenlage der Alveolenebene der Prämolaren.[7][5]
Das vordere Gebiss wies in der oberen und unteren Reihe jeweils zwei Schneidezähne je Kieferbogen auf. Der obere innere Schneidezahn zeigte sich spatelförmig, der äußere war schmaler. Die unteren Schneidezähne besaßen schmale und moderat hohe Zahnkronen. Sie ragten leicht nach vorn und bildeten eine geschlossene Reihe mit einer Position deutlich vor dem Eckzahn. Zu diesem bestand kein ausgeprägtes Diastema. Sowohl der obere wie auch der untere Eckzahn erhoben sich nur wenig über den Rand der hinteren Zähne. Sie ähnelten in ihrem Größenverhältnis weitgehend denen der Springaffen oder Nachtaffen und waren nicht vergrößert wie bei den Bartsakis oder den Kapuzineraffen beziehungsweise verkleinert wie bei den Marmosetten. Wie nahezu alle Neuweltaffen und abweichend von den Altweltaffen kam ein zweiter Prämolar vor, der den Beginn der hinteren Zahnreihe markiert. Die oberen Prämolaren charakterisierte ein großer Haupthöcker, der Paraconus, die beiden hinteren Vormahlzähne besaßen außerdem einen gut entwickelten Protoconus und zusätzlich noch einen Hypoconus, der auf dem letzten Vormahlzahn besonders groß entwickelt war. Der Hypoconus fehlt bei den rezenten Neuweltaffen. Am unteren vordersten Prämolar trat gelegentliche eine Schleiffazette auf, die durch den oberen Eckzahn bei Gebissschluss entstand. An den oberen hinteren Backenzähnen waren am ersten Molaren drei und am zweiten Molaren zwei Wurzeln ausgebildet. Die beiden vorderen oberen Molaren besaßen vier Haupthöcker mit einem auf der Wangenseite gegenüber dem Metaconus höheren Paraconus (wobei das Merkmal auf dem ersten Molaren weniger deutlich ist als auf dem zweiten). Beide standen weit auseinander mit langgestreckten Scherleisten dazwischen. Auf der Zungenseite nahm der große Hypoconus im Bezug auf den Protoconus eine leicht nach innen versetzte Position ein. Neben diesen vier Haupthöckern kam zusätzlich ein kleiner Nebenhöcker vor, der Paraconule. Sein Auftreten gilt als ursprüngliches Charakteristikum, das bei heutigen Neuweltaffen mit Ausnahme des Springtamarins nicht mehr ausgebildet ist. An der Zahninnen- und -außenseite schmiegte sich ein Cingulum an (ein niedriger Zahnschmelzwulst), wobei das innere deutlicher entwickelt war. Auf dem letzten Molaren fehlte der Hypoconus und der Paraconus war sehr groß, der Metaconus dagegen sehr klein. Die unteren Mahlzähne verfügten über jeweils zwei Wurzeln, was auch den hintersten Molaren einschließt. Das Merkmal kommt ebenso bei anderen frühen Neuweltaffen wie Branisella oder Carlocebus vor, bei den meisten heutigen Vertretern mit Ausnahme der Brüllaffen ist dieser Zahn einwurzelig. Die Kauoberfläche wurde durch vier Höckerchen geprägt. Teilweise war auf dem vordersten Molar ein zusätzlicher fünfter Höcker, das Paraconid, vorhanden, er ist bei modernen Neuweltaffen reduziert. Als ebenfalls auffallend urtümlich kann die Ausprägung des Hypoconulids, eines kleinen Nebenhöckers etwa in der Zahnmitte, angesehen werden.[3][4][5][6] Die obere Zahnreihe vom zweiten Schneidezahn bis zum letzten Mahlzahn wurde 2,7 cm lang, die Mahlzähne nahmen davon 1,1 cm in Anspruch. In der Regel war der vorderste Mahlzahn am größten ausgebildet.[2][6]
Vom postcranialen Skelett wurden bisher der Oberarmknochen, die Speiche und der Oberschenkelknochen beschrieben, die aufgrund der Proportionsverhältnisse zum Schädel im Vergleich mit heutigen Neuweltaffen zu Homunculus gestellt werden können. Sie ähneln weitgehend den entsprechenden Gliedmaßenknochen der rezenten Vertreter der Neuweltaffen, weisen aber einzelne Besonderheiten auf. Das obere Gelenkende und der Schaft waren deutlich breiter und robuster ausgeprägt als bei den heutigen Formen. Den Schaft umlief eine kräftige deltopectorale Leiste als Ansatzstelle der Armmuskulatur, was ein wenig an die Lemuren erinnert. Eine weitere Leiste zum unteren Ende hin fungierte als Ansatz für den Musculus brachioradialis. Sie war weiter nach oben verlängert als bei den meisten heutigen Neuweltaffen. Am unteren Ende des Oberarmknochens (Ellenbogengelenk) trat eine zylindrische Gelenkrolle (Trochlea) mit einer nur wenig erhabenen Mittellinie auf. Das seitlich anliegende Capitulum zeigte sich rund und oben breiter als unten. Die äußere sowie die innere Epicondyle erschienen beide eher groß, vor allem die innere war seitlich ausgezogen. Am insgesamt 11,2 cm langen Oberschenkelknochen hing der Große Rollhügel markant über dem vorderen Schaft und war außerdem sehr breit. Des Weiteren verlief zwischen dem Großen und dem Kleinen Rollhügel eine auffallende Knochenleiste. Die Gelenkfläche des Oberschenkelkopfes verlängerte sich zudem nach hinten. Am unteren Ende des Femurs trat eine tiefe Patellagrube auf, die nach oben gestreckt war und seitlich eine Lippe aufwies. Das Gelenkende war seitlich schmal, jedoch vorn und hinten verdickt.[7][8][9]
Fossilfunde
Die bekannten Fossilfunde von Homunculus stammen aus der Santa-Cruz-Formation in Patagonien. Diese Formation ist eine der bedeutendsten Fossillagerstätten des frühen Neogens in Südamerika. Ihre Ablagerungen erstrecken sich zwischen dem Lago Buenos Aires im Norden und dem Río Turbio im Süden, etwa zwischen dem 50. und 52. südlichen Breitengrad. Die wichtigsten Aufschlüsse finden sich perlschnurartig aufgereiht an der Atlantikküste in der argentinischen Provinz Santa Cruz zwischen den Flüssen Río Coyle und Río Gallegos. Einzelne bedeutende Fundpunkte liegen aber auch weiter nördlich des Río Coyle und im Landesinnern. An der Atlantikküste erreicht die Santa-Cruz-Formation eine Mächtigkeit von rund 240 m und setzt sich aus zwei nahezu gleichmächtigen Untereinheiten zusammen: dem Estancia-La-Costa-Schichtglied im Liegenden und dem Estancia-La-Angelina-Schichtglied im Hangenden. Beide Ablagerungsfolgen bestehen weitgehend aus Ton-, Ton-/Schluff- und Sandsteinen mit einigen eingeschalteten Lagen aus pyroklastischen Sedimenten. Die unteren, eher feinkörnigen Sedimente gehen auf einen ehemaligen Ästuar zurück, der nach und nach einer Überschwemmungsebene wich. Die oberen, gröberkörnigen Ablagerungen entstanden weitgehend unter fluviatilem Einfluss. Verschiedene radiometrische Datierungsverfahren geben der Santa-Cruz-Formation ein Alter von 18 bis 16 Millionen Jahren, was dem Übergang vom Unteren zum Mittleren Miozän entspricht.[10][11][12]
Die Entdeckung des Fossilreichtums der Santa-Cruz-Formation datiert in die Mitte des 19. Jahrhunderts, intensivere Erforschungen begannen aber erst in den 1880er bis 1890er Jahren und sind untrennbar mit den Namen Carlos und Florentino Ameghino verbunden.[13][14] Seit mehr als 100 Jahren im Fokus der wissenschaftlichen Forschung können heute gut zwei Dutzend Fossillagen innerhalb der Gesteinsformation unterschieden werden. Die Fossilfunde gehen in die Tausende und setzen sich aus fast allen höheren Gruppen der Wirbeltiere zusammen. Untergeordnet ließen sich auch Reste von Wirbellosen und von Pflanzen nachweisen. Herausragend ist die Säugetiergemeinschaft bestehend neben Beuteltieren vor allem aus den Vertretern der Höheren Säugetiere wie Nebengelenktieren, Nagetieren und Südamerikanischen Huftieren.[15] Die Bedeutung der Säugetierfunde drückt sich in der chronostratigraphischen Stufe des Santacruziums aus, für die die Santa-Cruz-Formation Pate stand und die als Abschnitt in der Entwicklung der südamerikanischen Landsäugetiere (South American Land Mammal Ages, SALMA) den Zeitraum von vor 17,5 bis 16,3 Millionen Jahren abdeckt. Die geologischen und paläontologischen Daten lassen eine Landschaft rekonstruieren, die aus weiten, offenen Savannengebieten durchsetzt mit Au- und Galeriewäldern bestand. Diese waren einem subtropischen Klima ausgesetzt mit einem im Vergleich zu heute relativ hohen Jahresniederschlag von 1000 bis 1500 mm. Die damals fehlende Blockade durch die Anden, die sich erst später auffalteten, ermöglichte die Passage kräftiger westlicher Winde.[16][17]
Bedeutende Funde von Homunculus wurden von Carlos Ameghino während seiner Expeditionen zur Santa-Cruz-Formation zwischen 1891 und 1894 gemacht. Dazu zählen Kiefer- und Schädelfragmente sowie einzelne Elemente des Körperskeletts. Ein linker Teilschädel mit dem erhaltenen Augenfenster (Exemplarnummer MACN-A 5968) von der Küstenfundstelle Puesto estancia La Costa (damals unter Corrigüen Aike bekannt) avancierte dabei zu dem ikonischen Fossil, das die gesamte Primatengattung repräsentierte. Die gleiche Fundstelle barg auch einen nahezu vollständigen Unterkiefer (MACN-A 5757), der möglicherweise mit einem Oberschenkelknochen sowie einer Elle und einer Speiche assoziiert war. Andere Funde, wie mehrere Unterkieferreste oder isolierte Zähne, förderte Carlos Ameghino an der ebenfalls küstennahen Fundstelle Monte Observación zu Tage. Für zwei weitere Unterkieferfragmente gab er nur eine ungefähre Position mit Río Gallegos an, während ein drittes Unterkieferfragment in seiner Originallage dagegen umstritten ist. Letzteres wies aber noch Teile des Milchgebisses auf und stellt so einen forschungsgeschichtlich frühen Nachweis eines Jungtiers dar.[18][5][9] Nur wenige Jahre darauf, zwischen 1898 und 1904, besuchte André Tournouër mehrfach die Santa-Cruz-Formation und entdeckte dabei unter anderem ein Unterkieferfragment nahe dem Río Coyle.[19]
Nach diesen wichtigen frühen Funden kamen erst fast 90 Jahre später wieder neue Fossilreste von Homunculus zum Vorschein. Getätigt wurden diese 1988 während einer Expedition der Stony Brook University von New York und dem Museo Argentino de Ciencias Naturales Bernardino Rivadavia. Das Material, zusammengesetzt aus einzelnen Zähnen und zwei Unterkieferfragmenten, lagerte an der Fundstelle Monte Observación.[20] Unabhängig davon erbrachte im gleichen Zeitraum eine Untersuchung der Universidad de Córdoba südlich des Río Coyle ein Schädeldach (CORD-PZ 1130) von der Fundstelle Puesto estancia La Costa.[2] Ein weiterer Primatenrest in Form eines Gesichtsschädels (MPM-PV 5000) kam bei einer dritten gleichzeitig stattfindenden Expedition einer Forschergruppe im Transit nach Antarktika in Killik Aike Norte zum Vorschein.[21][9] Im Jahr 2003 starteten größere Vorortuntersuchungen als Gemeinschaftsprojekt der Duke University und dem Museo de La Plata, bei dem auch zahlreiche internationale Forscher beteiligt waren. Die Feldforschungen dauern bis heute an. Ein früher Erfolg war die Entdeckung von drei Schädeln von Homunculus (MPM-PV-3501 bis 3503) zuzüglich einzelner Langknochen in Puesto estancia La Costa und Killik Aike Norte,[3][8] später kam noch der Schädel eines jungen Individuums hinzu (MPM-PV 3505).[22][6] Zunehmend stehen auch die küstenferneren Gebiete im Fokus. Unter anderem an den Aufschlüssen Barrancas Blancas und Segundas Barrancas Blancas am Río Santa Cruz und zusätzlich am Río Bote wurden zwischen 2014 und 2017 verschiedene Unterkieferreste und einzelne Zähne geborgen.[23] Die intensiven Grabungstätigkeiten führten dazu, dass das Fundmaterial von Humunculus auf mehr als ein halbes Dutzend Schädel einschließlich dem eines Jungtiers und zahlreiche Einzelzähne sowie verschiedene Elemente des Bewegungsapparates anwuchs.[5][9]
Paläobiologie
Körpergröße und Gehirnvolumen
Für die Berechnung des Körpergewichts von Homunculus können verschiedene Skelettelemente herangezogen werden. Anhand des Oberschenkelknochens lässt sich ein Gewicht von 1,9 bis 2,5 kg annehmen. Die Schädelmaße insgesamt führen zu einem vermutlichen Gewicht von 1,9 bis 2,4 kg, was sich bei Heranziehung des unteren ersten Mahlzahns bestätigen lässt; hier ergeben sich Vergleichsgrößen von 1,9 bis 2,7 kg. Unter Bezug auf die ersten oberen Mahlzähne wurden Werte von 2,5 bis 3,4 kg berechnet. Die leicht höheren Gewichtsangaben können mit der Hypertrophie der oberen Molaren zusammenhängen. Letzte Maße außenvorgelassen dürfte Homunculus etwa die Größe der heutigen Sakis oder Uakaris besessen haben. Außerhalb der hier angegebenen Variationsbreite lässt sich anhand des vorderen Molars eines extrem großen Unterkiefers von Homunculus ein Körpergewicht von 3 kg annehmen.[9]
Für zwei Schädelausgüsse ergibt sich ein Gehirnvolumen von rund 20 cm³. Dies ist relativ klein im Vergleich zur Körpergröße und liegt unterhalb der Werte heutiger Neuweltaffen. Erheblichen Einfluss auf die geringe Gehirngröße hat der nur wenig entwickelte Frontallappen bei Homunculus. Auch für den ebenfalls miozänen frühen Neuweltaffen Chilecebus wurde ein sehr geringes Gehirnvolumen bei noch kleinerer Körpergröße ermittelt.[24][25] Es ist daher wahrscheinlich, dass die Neuweltaffen im Laufe ihrer Stammesgeschichte größere Gehirnvolumen im Verhältnis zur Körpergröße entwickelten, wofür verschiedene Faktoren mit ernährungs- und soziobiologischem Hintergrund verantwortlich sind. Dies entspricht auch den Befunden innerhalb der Altweltaffen.[26][9]
Fortbewegung
In den Proportionen seiner Gliedmaßen ähnelt Homunculus weitgehend den heutigen Neuweltaffen. Anhand des Baus der Langknochen kann auf die Fortbewegungsweise geschlossen werden. So wirkt am Oberschenkelknochen der überhängende Große Rollhügel als Ansatzpunkt des Musculus vastus lateralis, der einen kräftigen Kniestrecker repräsentiert. Das wiederum seitlich schmale sowie vorn und hinten verdickte untere Gelenkende verlängert unter anderem den Hebelarm der Kniestreckermuskulatur. Der bei der Beinstreckung auftretenden starken mechanischen Beanspruchung der Kniescheibe wirkt die tiefe Patellagrube entgegen. All diese Charakteristika sind typisch für eine springende Fortbewegung bei den Neuweltaffen und finden sich etwa bei den Sakis und dem Satansaffen wieder. Vergleichbares lässt sich auch am Oberarmknochen erkennen. Sein robuster Bau mit den markanten Knochenleisten wie der deltopectoralen Leiste und dem breiten und charakteristisch gestalteten unteren Gelenkende (Ellenbogengelenk) spricht für eine hohe Bewegungsfreiheit des Unterarms, worauf letztendlich auch die kräftige und recht markant gebogene Speiche hindeutet. Die gestreckte und seitlich gerichtete innere Epicondyle von Homunculus kann entsprechend bei zahlreichen kletternden Primaten beobachtet werden, während diese sich bei bodenlebenden Arten nach hinten orientiert. Dagegen dehnt sich bei keinem der Oberarmknochen von Homunculus die Gelenkfläche der Trochlea nach oben aus. Eine solche vergrößerte Gelenkfläche ist typisch für zahlreiche Primaten, etwa Krallenaffen oder Lemuren, und ermöglicht als sogenannte „Klammerfazette“ das senkrechte Klettern mit weit ausholenden Armen (hier presst der Kronenfortsatz der Elle bei Armstreckung dagegen). Gemeinsam mit der auffälligen Form des Capitulums lässt sich daraus für das Ellenbogengelenk eine offensichtlich überwiegend gebeugte denn gestreckte Position bei der Fortbewegung schlussfolgern. Die gesamte Konfiguration indiziert darüber hinaus ein recht ausgedehntes Olecranon an der Elle, welche bisher nicht fossil überliefert ist. Ein verlängertes Olecranon erhöht die Hebelkraft der Unterarmmuskulatur. Ein größerer Teil der genannten Charakteristika ist auf eine baumbewohnende Lebensweise zurückzuführen und tritt bei heutigen bodenlebenden Primaten nicht auf. In der Summe gibt es aber nur wenige Hinweise auf eine schwinghangelnde oder senkrecht im Geäst kletternde Bewegung bei Homunculus. Vielmehr dürfe die Primatenform vierbeinig laufend unterwegs gewesen sein.[8][27][9]
Die relativ großen Bogengänge des Innenohrs verweisen auf ein recht agiles Tier wie etwa die heutigen Springaffen, aber weniger schnell als etwa Totenkopfaffen oder Tamarine. Die Bogengänge stehen außerdem im rechten Winkel zueinander. Das gesamte Gleichgewichtsorgan ließ Kopfdrehbewegungen von rund 135° je Sekunde zu, was der Größenordnung heutiger Lemuren wie Bambuslemuren und verschiedenen Sifakas entspricht.[28][9]
Ernährungsweise
Der Unterkiefer und Schädel zeigen durch den fehlenden Scheitelkamm, die wenig ausgeprägten Temporallinien und den schwach ausladenden Jochbogen kaum starke Muskelansatzstellen im Vergleich zu gleich großen heutigen Neuweltaffen. Hinzu kommt die relative Prognathie der Schnauze von Homunculus, die andere Neuweltaffen übertrifft. Dadurch entstanden für die Kaumuskeln lange Lade-, aber kurze Hebelarme, was zu einer weitgehend schwach entwickelten Kaumuskulatur mit verminderter Beißkraft führt. Durch die vollständig verwachsene Symphyse bestand aber prinzipiell die Möglichkeit, erhebliche Muskelkraft vor allem an der gegenüber der Kaubewegung wirkenden Seite bereitzustellen. Ebenso verweist die ausgedehnte Gelenkfläche zur Verbindung des Unterkiefers mit der Schädelunterseite darauf, dass sich wiederholende Kraftentladungen beim Kauen oder beim Beißen mit den Schneidezähnen kompensieren ließen.[29][9]
Die spatelförmigen oberen und schmalen sowie vorstehenden unteren Schneidezähne konnten in Verbindung mit dem verhältnismäßig kleinen Eckzahn zum Knacken harter Früchte und Samen wie bei den Sakis eingesetzt werden. Entgegen den Sakis mit ihren größeren Eckzähnen und stärkeren Wurzeln dort durchbohrten diese bei Homunculus die Nahrung aber nicht. Allerdings ist für das vordere Gebiss auch eine baumrindenschabende Funktion zur Gewinnung von Baumsäften wie bei den Büschelaffen denkbar. An den relativ großen Mahlzähnen bestehen vergleichsweise ausgedehnte Scher- und Schneidekanten. Diese sind zum Zerkleinern von blatthaltiger Nahrung oder weichen Früchten geeignet, in Übereinstimmung mit den entsprechenden Zähnen der Wollaffen oder der Brüllaffen. Neuweltaffen, die überwiegend hartschalige Samen und Früchte verzehren, besitzen dem gegenüber kürzere Schneidkanten an den Mahlzähnen. Bei ihnen ist außerdem der Zahnschmelz massiver ausgebildet, was für Homunculus nicht zutrifft. Auch haben die rezenten, blätterfressenden Neuweltaffen größere Zahnwurzeln an den Backenzähnen als hartsamenverzehrende Arten, ein Merkmal dass ebenso bei Homunculus auftritt. Die häufig stärker abgekauten Zähne bei Homunculus führen aber wiederum zu der Vermutung, dass die weiche Pflanzennahrung stärker faserhaltig war und so einen höheren Abrieb erzeugte. Demnach erfolgte die Zerkleinerung der Nahrung im Maul trotz schwach entwickelter Kaumuskulatur mittels starker Kaubewegungen, was einerseits die großen Zahnwurzeln, andererseits die auffälligen Abrasionen hervorrief.[30] Der Gesamtbefund verweist somit auf eine überwiegende Ernährung von hartschaligen Samen mittels der Vorderzähne, während das hintere Gebiss die Aufnahme von weicheren Früchten und von Blättern befürworte. Diese funktionale Trennung beruht eventuell auf einem jahreszeitlich wechselnden Nahrungsangebot. Für Homunculus kann aus diesen Gründen ein Leben in tropischen Regenwäldern eher ausgeschlossen werden. Als mögliche Habitate kommen saisonal beeinflusste Wälder oder solche in Randlagen wie etwa Galeriewälder in Betracht. Das Ernährungsverhalten von Homunculus dürfte so weitgehend den heutigen Springaffen der atlantischen Küstenwälder entsprochen haben.[31][29][27][9]
Geschlechtsdimorphismus
Heutige Neuweltaffen besitzen variabel ausgebildete Eckzähne zwischen den einzelnen Geschlechtern mit einem starken Dimorphismus bei den Brüllaffen und einem geringen bei einigen Springaffen und dem Nachtaffen. Die unterschiedliche Ausprägung steht teilweise mit der Lebensweise in Verbindung, wobei ein starker Eckzahndimorphismus auf die Dominanzkämpfe der Männchen um das Paarungsvorrecht und ein territoriales Verhalten verweist, während ein geringer Eckzahndimorphismus ein Leben in mitunter monogamen Gruppen anzeigt. Im Fossilmaterial von Homunculus finden sich nur wenige Variationen bezüglich der Eckzahngröße, unter der Voraussetzung, dass das bisherige Fundmaterial beide Geschlechter repräsentiert, ist demzufolge kein sonderlicher Dimorphismus auszumachen. Daraus wäre zu schlussfolgern, dass Kämpfe unter den Männchen wohl eher unüblich waren und eine Selektion in Richtung starker Eckzähne nicht erfolgte. Konkurrenz bestand dann weniger zwischen den einzelnen Individuen der jeweiligen Geschlechter als eventuell vielmehr zwischen den verschiedenen Gruppen oder Paaren. In diesem Merkmal gleicht der ausgestorbene Primatenvertreter deutlicher den Springaffen oder den Nachtaffen.[31][9]
Sinneswahrnehmungen und Laute
Die Größe der Orbita im Vergleich zum Schädel verweist Homunculus zu den tagaktiven Primaten. Die Ansicht unterstützt der nahezu geschlossene Orbitarand, der unter anderem das umgebende Bindegewebe stabilisiert und das Auge so vor zu starken Bewegungen der Kaumuskulatur schützt. Der so besser geschützte Augapfel konnte eine stärkere Sehschärfe ausbilden, die bei tagaktiven Primaten prinzipiell höher ist als bei nachtaktiven. Darüber hinaus ist das Foramen opticum als Durchtrittspunkt des Sehnervs in den Canalis opticus bei Homunculus relativ groß und liegt innerhalb der Variationsbreite heutiger tagaktiver Neuweltaffen. Dadurch kann angenommen werden, dass die ausgestorbene Primatenform eine Fovea centralis mit hoher Anzahl an zapfenförmigen Lichtrezeptoren besaß. Dem gegenüber ist das Foramen infraorbitale stark reduziert. Hier verläuft der Unteraugennerv, der sensorische Impulse von den Vibrissen der Schnauzenregion zum Gehirn übermittelt. Ein kleines Unteraugenloch steht somit mit weniger gut entwickelten Tasthaaren in Verbindung, was wiederum für ein eher tageslichtaktives Leben spricht. Letztendlich ist auch die Fossa olfactoria, durch die der Riechkolben verläuft, bei Homunculus vergleichsweise klein. Im Gegensatz dazu haben heutige Nachtaffen eine große Orbita, ein großes Foramen infraorbitale und eine große Fossa olfactoria.[32][9]
Der nach hinten tiefer werdende Unterkiefer findet seine Entsprechung bei den Spring- und Nachtaffen wieder und steht hier im Zusammenhang mit sackartigen Erweiterungen des Kehlkopfes (Laryngeal air sacs), die bei vielen, aber nicht allen Primaten vorkommen und zur Erzeugung kräftiger Laute als Teil des ritualisierten Territorialverhaltens dienen. Dadurch können sich kleine Gruppen relativ weit verteilen und über Lautkommunikation in Verbindung bleiben, was wiederum effektiv in weniger ertragreichen Landschaften ist.[31] Wahrgenommene Frequenzbereiche lassen sich über die Anatomie des Ohrs rekonstruieren. Prinzipiell haben heutige Trockennasenprimaten (Alt- und Neuweltaffen) ein ausgeprägteres Gehör für niedrigere Frequenzbereiche, während Feuchtnasenprimaten (Lemuren und Loris) auf eher höhere Frequenzen spezialisiert sind. Innerhalb der Trockennasenprimaten können Altweltaffen niedrige Frequenzen besser wahrnehmen als Neuweltaffen. Die Menschenartigen dem gegenüber sind wenig sensitiv für höhere Frequenzen. Bei Homunculus erreicht das Trommelfell am Eingang zum Mittelohr eine Fläche von 23,8 mm², was etwa dem des heutigen Springtamarins entspricht, der aber kleiner als Homunculus ist. Auch haben zahlreiche heutige Primaten im Verhältnis zu ihrer Körpergröße flächenmäßig ausgedehntere Trommelfelle als Homunculus. Als weiterer Wert kann die Größe der Fußplatte des Steigbügels (beziehungsweise des Ovalen Fensters zum Innenohr, in dem die Fußplatte verankert ist) herangezogen werden. Beides beträgt bei Homunculus etwa 0,6 mm², was etwas weniger ist als bei zahlreichen rezenten Primaten. Sehr genaue Ergebnisse liefert die Hörschnecke des Innenohrs, da beispielsweise die Anzahl ihrer Umdrehungen sowie ihre Gesamtlänge in direktem Zusammenhang mit der Wahrnehmung niederfrequenter Töne stehen. Die Hörschnecke weist bei Homunculus drei vollständige Umdrehungen bei einer Gesamtlänge der Windungen von 24,6 mm auf. Der Umdrehungswert liegt im Bereich heutiger Neuweltaffen, aber über dem zahlreicher Feuchtnasenprimaten mit ihren weniger als drei Windungen. In allen drei Merkmalen verfügte Homunculus über eine gute Sensitivität für sowohl niedrigere als auch höhere Laute. Niederfrequente Töne um 250 Hz wurden bei einem Schalldruck zwischen 24 und 36 dB wahrgenommen (zum Vergleich: Altweltaffen 12 bis 19 dB, Neuweltaffen 24 bis 26 dB, Lemuren und Loris 32 bis 48 dB). Im höheren Bereich betrug die Wahrnehmschwelle von Frequenzen um 32 kHz rund 13 dB (zum Vergleich: Altweltaffen 22 bis 39 dB, Neuweltaffen 8 bis 14 dB, Lemuren und Loris 7 bis 23 dB). Damit entspricht Homunculus weitgehend den heutigen Neuweltaffen. Das Hörvermögen der Neuweltaffen war damit wohl schon im Unteren/Mittleren Miozän weitgehend ausgeprägt, erfuhr aber in der Folgezeit noch einzelne Verfeinerungen.[33][34][9]
Systematik
Innere Systematik der Neuweltaffen gemäß der Long Lineage Hypothesis nach Silvestro et al. 2019[35]
gekürzt; dargestellt sind neben den rezenten Gruppen nur die Fossilformen vom Eozän bis zum Unteren Miozän |
Innere Systematik der Neuweltaffen gemäß der Stem Platyrrhine Hypothesis nach Marivaux et al. 2016[36]
gekürzt; dargestellt sind neben den rezenten Gruppen nur die Fossilformen vom Eozän bis zum Unteren Miozän |
Homunculus ist eine Gattung aus der Ordnung der Primaten (Primates). Innerhalb dieser wird die Gattung zur Gruppe der Neuweltaffen (Platyrrhini) gezählt. Hierbei handelt es sich um zumeist baumlebende Affen, die die tropischen sowie subtropischen Gebiete Süd- und Mittelamerikas bewohnen. Äußerlich unterscheiden sie sich von den Altweltaffen durch ihre verhältnismäßig breitere Nase, ihre durchschnittlich geringere Körpergröße und den generell ausgebildeten Schwanz, der bei einigen Formen als Greiforgan einsetzbar ist. Die Neuweltaffen umfassen mehrere Familien, von denen die Sakiaffen (Pitheciidae), die Klammerschwanzaffen (Atelidae), die Kapuzinerartigen (Cebidae) und die Krallenaffen (Callitrichidae) die bedeutendsten sind. In der Regel gelten die Sakiaffen als Schwestergruppe aller anderen Neuweltaffen, was sich molekulargenetisch belegen lässt.[37][38][39] Der früheste Fossilbeleg der Neuweltaffen stammt mit Perupithecus aus dem peruanischen Bereich des Amazonasbeckens und datiert in den Übergang vom Eozän zum Oligozän vor rund 35 Millionen Jahren.[40]
Die systematische Stellung von Homunculus ist gegenwärtig nicht eindeutig. Mitunter wird die Form keiner bestimmten Familie zugeordnet und ihr Status als incertae sedis ausgewiesen.[6] Sie gehört dann zu einer Gruppe früher patagonischer Formen, die als Stammgruppe der Neuweltaffen fungiert (teilweise werden diese in eine eigenständige Familie, die Homunculidae, eingeordnet, welche 1894 von Florentino Ameghino eingeführt worden war[41]).[42][36] Andere Autoren gruppieren Homunculus wiederum innerhalb der Sakiaffen, der Klammerschwanzaffen oder der Kapuzinerartigen (manchmal mit den Homunculinae im Rang einer Unterfamilie oder als Homunculini im Rang einer Tribus).[5][43][44]
Die abweichenden Zuweisungen gehen auf zwei konkurrierende Hypothesen zur Phylogenese der Neuweltaffen zurück: einerseits die Long Lineage Hypothesis oder Morphological Stasis Hypothesis, nach welcher sich die ausgestorbenen miozänen Gattungen als ursprüngliche Vertreter in die Kronengruppe der Neuweltaffen einreihen lassen, andererseits die Stem Platyrrhine Hypothesis oder Successive Radiations Hypothesis, die die urtümlichen Formen als Teil einer basalen Radiationsphase einstuft. Beide Ansichten werden gegenwärtig kontrovers diskutiert.[45][46][36] Die zur Untermauerung der jeweiligen Ansicht herangezogenen molekulargenetischen Untersuchungen an rezenten Neuweltaffen sind nicht eindeutig, da ihnen unterschiedliche Kalibrationsmodelle zugrunde liegen. Demnach setzen die Verfechter der Long Lineage Hypothesis die Aufspaltung der heutigen Neuweltaffen vor 29 bis 31 Millionen Jahren an,[47] die der Stem Platyrrhine Hypothesis vor rund 20 Millionen Jahren.[48]
Taxonomie- und Forschungsgeschichte

Die wissenschaftliche Erstbeschreibung von Homunculus stammt von Florentino Ameghino aus dem Jahr 1891. Er tätigte sie kurz nach der Entdeckung der ersten Funde durch seinen Bruder Carlos Ameghino in der Santa-Cruz-Formation. Florentino Ameghinos Einführung von Homunculus bedeutete damals den ältesten Nachweis der Neuweltaffen in Südamerika und den erst zweiten Beleg eines ausgestorbenen Primaten auf dem Kontinent nach Peter Wilhelm Lunds Benennung von Protopithecus aus dem Pleistozän Brasiliens im Jahr 1840. Die Erstbeschreibung von Homunculus basiert auf einigen Unterkieferfragmenten, von denen ein rechter Kieferast mit der Symphyse und den Zähnen vom zweiten Schneidezahn bis zum ersten Mahlzahn als Holotyp gilt (Exemplarnummer MACN-A 634). Er stammt aus der Region des Río Gallegos. Genauere Angaben legte Florentino Ameghino nicht vor, doch es wird vermutet, dass der Fund von dessen Nordseite stammt und möglicherweise aus Killik Aike Norte zu Tage gefördert wurde.[9] Das Objekt ist seit mehreren Jahrzehnten verschollen, weswegen im Jahr 2008 mit einem nahezu vollständigen Unterkiefer, dessen Zähne aber stärker abgekaut sind, aus der wenig weiter nördlich gelegenen Fundstelle Puesto estancia La Costa ein Neotyp (Exemplarnummer MACN-A 5757) festgelegt wurde.[5] Florentino Ameghino benannte als einzige Art H. patagonicus. Provisorisch sprach er die neue Primatenform als lemurenartig an, sah sie aber aufgrund seiner Meinung nach sehr modernen Merkmalen in direkter Entwicklungslinie zu den Menschen und dessen näherer Verwandtschaft.[49][50][9]
Noch im gleichen Jahr benannte Florentino Ameghino Anthropops perfectus bezugnehmend auf einen Unterkiefer mit stärker V-förmig verlaufenden Kieferbögen und breiterer Symphyse als im Vergleich zu Homunculus. Der Fund kam im Küstengebiet der Santa-Cruz-Formation zu Tage ohne Angabe einer genaueren Fundregion.[50] Drei Jahre später wiederum führte er Pitheculus australis anhand eines linken Unterkieferfragments vom Monte Observación ein. Beide werden heute zu H. patagonicus gestellt. Als problematisch ist die von Alcides Mercerat 1891 unabhängig von Florentino Ameghinos Publikationen zur Santa-Cruz-Formation aufgestellte Primatenform Ecphantodon ceboides zu betrachten, die auf einem fragmentierten Unterkiefer mit Mahlzahn fußt.[51] Die Beschreibung ist knapp gehalten und nicht illustriert, Florentino Ameghino setzte die Art noch im selben Jahr mit H. patagonicus gleich ohne aber Gründe dafür anzugeben.[50] Da der Fund verloren gegangen ist und keine weiteren Details darüber vorliegen, folgen die meisten Wissenschaftler heute dieser Einschätzung.[5][9][6]
Hans Bluntschli, der 1912 kurz nach dem Tod von Florentino Ameghino während eines Aufenthalts in Argentinien dessen Sammlung einsah und 1931 eine umfangreiche Neubeschreibung der Primatenreste vorlegte, postulierte die Art H. ameghinoi,[7] die jedoch weitgehend nicht anerkannt ist. Die ebenfalls von Florentino Ameghino 1894 benannte Art H. imago, für die er einen Oberschenkelknochen und eine Elle heranzog, ist höchstwahrscheinlich keine Primatenform. Dem gegenüber verwies Florentino Ameghino zum gleichen Zeitpunkt einen rechten Backenzahn zu Stilotherium grande, einen Vertreter der Beuteltiere.[41] Mehr als 80 Jahre später erkannte Philip Hershkovitz in dem Fundstück den Milchzahn eines Primaten und benannte die Form mit H. grandis.[18] Sie wird heute als synonym zu H. patagonicus eingestuft. Eine weitere Art benannte Carlos Rusconi mit H. harringtoni im Jahr 1933.[52] Aufgestellt anhand eines nahezu vollständigen Schädels aus der argentinischen Provinz Chubut datiert dieser in das Untere Miozän und ist somit etwas älter als Homunculus. Hershkovitz stellte für ihn im Jahr 1974 die neue Gattung Tremacebus auf, da die Form mit ihren deutlich größeren Augenfenstern markant von Homunculus abweicht.[53] In den 1980er Jahren brachte John Fleagle einzelne isolierte Zähne aus der Pinturas-Formation mit Homunculus in Verbindung, verschob sie aber 1990 in die von ihm etablierte Gattung Carlocebus. Die Pinturas-Formation ist ebenfalls in der argentinischen Provinz Santa Cruz aufgeschlossen und entspricht in ihrem Alter den unteren Abschnitten der Santa-Cruz-Formation.[54][5][9]
Im Jahr 2019 führten Richard F. Kay und Jonathan M. G. Perry mit H. vizcainoi eine weitere Art ein, deren Belegmaterial vom Río Santa Cruz stammt.[23] Erwähnt werden sollen hier noch ein Gesichtsschädel und ein Oberkieferfragment aus der Fundstelle Killik Aike Norte der Santa-Cruz-Formation, die im Jahr 2006 einem Forscherteam um Marcelo F. Tejedor als Grundlage zur Aufstellung der neuen Gattung Killikaike mit der Art K. blakei dienten. Die Autoren vermuteten aufgrund des kurzen Gesichts und der Größe des Gehirnschädels eine nähere Verwandtschaft mit den Kapuzineraffen.[21] Andere Wissenschaftler sahen das in der folgenden Zeit kritischer und stuften die Form als identisch mit Homunculus und dessen Nominatform ein,[6] während wiederum H. blakei von einzelnen Forschern als eigenständige Art innerhalb der Gattung Homunculus anerkannt ist. Demnach könnten mit H. patagonicus, H. vizcainoi und H. blakei drei Arten bestehen. Weitere bisher unbenannte Arten werden aufgrund eines untypisch robusten Unterkiefers von der Fundstelle Puesto estancia La Costa und aufgrund eines einzelnen Molars vom Río Bote angenommen.[9][23]
Innerhalb der Gattung lassen sich demnach bisher zwei eindeutige Arten unterscheiden:[23]
- H. patagonicus Ameghino, 1891
- H. vizcainoi Kay & Perry, 2019
Hierbei stellt H. patagonicus die Typusart dar und ist der größere von beiden Vertretern.[23]
Bedeutung
Die Fossilreste von Homunculus stellen mit einer Fundlage rund um den 51. südlichen Breitengrad die südlichsten einer in Amerika aufgefundenen Primatenart dar. Heutige Neuweltaffen erreichen am 29. südlichen Breitengrad ihre Ausbreitungsgrenze. Die Gattung Homunculus ist eingebettet in eine Gruppe früher Neuweltaffen aus Patagonien, die mit Tremacebus und Dolichocebus vor etwa 20 Millionen Jahren erschien und mit Proteropithecia vor rund 15,5 Millionen Jahren wieder aus dem Fossilbericht verschwand. Nach diesem Zeitpunkt sind bis heute keine Affen mehr aus dem südlichen Südamerika belegt. Im Vergleich zu dem eher spärlichen Fundmaterial der anderen miozänen Gattungen Patagoniens erlauben die reichen Reste von Homunculus den bisher besten Einblick in die biologischen Anpassungen eines frühen Neuweltaffen.[9]
Literatur
- Richard F. Kay, Jonathan M. G. Perry, Michael Malinzak, Kari L. Allen, E. Christopher Kirk, J. Michael Plavcan und John G. Fleagle: Paleobiology of Santacrucian primates. In: Sergio F. Vizcaíno, Richard F. Kay und M. Susana Bargo (Hrsg.): Early Miocene Paleobiology in Patagonia: High-Latitude Paleocommunities of the Santa Cruz Formation. Cambridge University Press, New York, 2012, S. 306–330
- Marcelo F. Tejedor und Alfred L. Rosenberger: A neotype for Homunculus patagonicus Ameghino, 1891, and a new Interpretation of the Taxon. PaleoAnthropology 2008, S. 68–82
Einzelnachweise
- Ethan L. Fulwood, Doug M. Boyer und Richard F. Kay: Stem members of Platyrrhini are distinct from catarrhines in at least one derived cranial feature. Journal of Human Evolution 100, 2016, S. 16–24
- Adan Tauber: Homunculus patagonicus Ameghino, 1891 (Primates, Ceboidea), Mioceno Temprano, de la costa Atlantica Austral, prov. de Santa Cruz, Republica Argentina. Academia Nacional de Ciencias Córdoba, Argentina Miscelanea 82, 1991, S. 1–32 ()
- R. F. Kay, S. F. Vizcaino, A. A. Tauber, M. S. Bargo, B. A. Williams, C. Luna und M. W. Colbert: Three newly discovered skulls of Homunculus patagonicus support its position as a stem platyrrhine and establish its diurnal arboreal folivorous habits. American Journal of Physical Anthropology 126 (S40), 2005, S. 127
- Richard F. Kay, J. G. Fleagle, T. R. T. Mitchell, Matthew Colbert, Tom Bown und Dennis W. Powers: The anatomy of Dolichocebus gaimanensis, a stem platyrrhine monkey from Argentina. Journal of Human Evolution 54, 2008, S. 323–382
- Marcelo F. Tejedor und Alfred L. Rosenberger: A neotype for Homunculus patagonicus Ameghino, 1891, and a new Interpretation of the Taxon. PaleoAnthropology 2008, S. 68–82
- Jonathan M. G. Perry, Richard F. Kay, Sergio F. Vizcaíno und M. Susana Bargo: Oldest known cranium of a juvenile New World monkey (Early Miocene, Patagonia, Argentina): Implications for the taxonomy and the molar eruption pattern of early platyrrhines. Journal of Human Evolution 74, 2014, S. 67–81
- Hans Bluntschli: Homunculus patagonicus und die ihm zugereihten Fossilfunde aus den Santa Cruz Schichten Patagoniens. eine morphologische Revision an Hand der Originaltücke in der Sammlung Ameghino zu La Plata. Morphologisches Jahrbuch 67, 1931, S-811–892
- John Fleagle und Richard Kay: A new humerus of Homunculus from the Santa Cruz Formation (Early-Middle Miocene, Patagonia). Journal of Vertebrate Paleontology 26 (3suppl.), 2006, S, 62A
- Richard F. Kay, Jonathan M. G. Perry, Michael Malinzak, Kari L. Allen, E. Christopher Kirk, J. Michael Plavcan und John G. Fleagle: Paleobiology of Santacrucian primates. In: Sergio F. Vizcaíno, Richard F. Kay und M. Susana Bargo (Hrsg.): Early Miocene Paleobiology in Patagonia: High-Latitude Paleocommunities of the Santa Cruz Formation. Cambridge University Press, New York, 2012, S. 306–330
- Sergio F. Vizcaíno, M. Susana Bargo, Richard F. Kay, Richard A. Fariña, Mariana Di Giacomo, Jonathan M. G. Perry, Francisco J. Prevosti, Néstor Toledo, Guillermo H. Cassini und Juan C. Fernicola: A baseline paleoecological study for the Santa Cruz Formation (late–early Miocene) at the Atlantic coast of Patagonia, Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 292, 2010, S. 507–519
- Sergio D. Matheos und M. Sol Raigenborn: Sedimentology and paleoenvironment of the Santa Cruz Formation. In: Sergio F. Vizcaíno, Richard F. Kay und M. Susana Bargo (Hrsg.): Early Miocene paleobiology in Patagonia: High-latitude paleocommunities of the Santa Cruz Formation. Cambridge University Press, New York, 2012, S. 59–82
- José I. Cuitiño, Juan Carlos Fernicola, Matthew J. Kohn, Robin Trayler, Maximiliano Naipauere, M. Susana Bargo, Richard F. Kay und Sergio F. Vizcaíno: U-Pb geochronology of the Santa Cruz Formation (early Miocene) at the Río Bote and Río Santa Cruz (southernmost Patagonia, Argentina): Implications for the correlation of fossil vertebrate localities. Journal of South American Earth Sciences 70, 2016, S. 198–210
- Sergio F. Vizcaíno, Richard F. Kay und M. Susana Bargo: Background for a paleoecological study of the Santa Cruz Formation (late Early Miocene) on the Atlantic Coast of Patagonia. In: Sergio F. Vizcaíno, Richard F. Kay und M. Susana Bargo (Hrsg.): Early Miocene paleobiology in Patagonia: High-latitude paleocommunities of the Santa Cruz Formation. Cambridge University Press, New York, 2012, S. 1–22
- Juan Carlos Fernicola, José I. Cuitiño, Sergio F. Vizcaíno, M. Susana Bargo und Richard F. Kay: Fossil localities of the Santa Cruz Formation (Early Miocene, Patagonia) prospected by Carlos Ameghino in 1887 revisited and the location of the Notohippidian. Journal of South American Earth Sciences 52, 2014, S. 94–107
- Darin A. Croft: What Constitutes a Fossil Mammal Community in the Early Miocene Santa Cruz Formation? Journal of Vertebrate Paleontology 33 (2), 2013, S. 401–409
- Adán A. Tauber: Paleoecología de la Formación Santa Cruz (Mioceno inferior) en el extremo sudeste de la Patagonia. Ameghiniana 34, 1997, S. 517–529
- Richard F. Kay, Sergio F. Vizcaíno und M. Susana Bargo: A review of the paleoenvironment and paleoecology of the Miocene Santa Cruz Formation. In: Sergio F. Vizcaíno, Richard F. Kay und M. Susana Bargo (Hrsg.): Early Miocene paleobiology in Patagonia: High-latitude paleocommunities of the Santa Cruz Formation. Cambridge University Press, New York, 2012, S. 331–365
- Philip Hershkovitz: More on the Homunculus Dpm4 and M1, and Comparisons With Alouatta and Stirtonia (Primates, Platyrrhini, Cebidae). American Journal of Primatology 7, 1984, S. 261–283
- Nelson M. Novo, Marcelo F. Tejedor und Laureano R. González-Ruiz: Previously unknown fossil platyrrhines (Primates) of Patagonia from the Tournouër collection at the Muséum national d’Histoire naturelle, Paris. Geodiversitas 40 (22), 2018, S. 529–535
- J. G. Fleagle, G. A. Buckley und M. E. Schloeder: New Primate Fossils from Monte Observación, Santa Cruz Formation (Lower Miocene), Santa Cruz Province, Argentina. Journal of Vertebrate Paleontology 8 (3suppl.), 1988, S. 14A
- Marcelo F. Tejedor, Adán A. Tauber, Alfred L. Rosenberger, Carl C. Swisher III und María E. Palacios: New primate genus from the Miocene of Argentina. PNAS 103 (14), 2006, S. 5437–5441
- Jonathan M. G. Perry, Richard F. Kay, Sergio F. Vizcaino und Susana M. Bargo: First cranial material of a juvenile monkey from the Tertiary of South America: cranial anatomy and dental eruption sequence. Journal of Vertebrate Paleontology 28 (3suppl.), 2008, S, 127A
- Richard F. Kay und Jonathan M.G. Perry: New primates from the Río Santa Cruz and Río Bote (Early–Middle Miocene), Santa Cruz Province, Argentina. Publicación Electrónica de la Asociación Paleontológica Argentina 19 (2), 2019, S. 230–238
- Karen E. Sears, John A. Finarelli, John J. Flynn und Andre R. Wyss: Estimating body mass in New World “monkeys” (Platyrrhini, Primates), with a consideration of the Miocene platyrrhine, Chilecebus carrascoensis. American Museum Novitates 3617, 2008, S. 1–29
- Xijun Ni, John J. Flynn, André R. Wyss und Chi Zhang: Cranial endocast of a stem platyrrhine primate and ancestral brain conditions in anthropoids. Science Advances 5, 2019, S. eaav7913, doi:10.1126/sciadv.aav7913
- Richard Kay und Michael Malinzak: New data on encephalization in Miocene New World monkeys: implications for anthropoid brain evolution. Journal of Vertebrate Paleontology 28 (3suppl.), 2008, S, 99A
- Dionisios Youlatos und Jeff Meldrum: Locomotor Diversification in New World Monkeys: Running, Climbing, or Clawing Along Evolutionary Branches. The Anatomical Record 294, 2011, S. 1991–2012
- Timothy M. Ryan, Mary T. Silcox, Alan Walker, Xianyun Mao, David R. Begun, Brenda R. Benefit, Philip D. Gingerich, Meike Köhler,László Kordos, Monte L. McCrossin, Salvador Moyà-Solà, William J. Sanders, Erik R. Seiffert, Elwyn Simons, Iyad S. Zalmout und Fred Spoor: Evolution of locomotion in Anthropoidea: the semicircular canal evidence. Proceedings of the Royal Society B 279, 2012, S. 3467–3475
- Jonathan M. G. Perry, Richard F. Kay, Sergio F. Vizcaíno und M. Susana Bargo: Tooth Root Size, Chewing Muscle Leverage, and the Biology of Homunculus patagonicus (Primates) from the late early Miocene of Patagonia. Ameghiniana 47 (3), 2010, S. 355–371
- Jonathan Perry und Matthew Colbert: Tooth root size and chewing muscle leverage in Homunculus, a Miocene primate from Patagonia. Journal of Vertebrate Paleontology 26 (3suppl.), 2006, S, 110A
- Alfred L. Rosenberger, Marcelo F. Tejedor, Siobhán B. Cooke und Stephen Pekar: Platyrrhine ecophylogenetics in space and time. In: P. A. Garber, A. Estrada, J. C. Bicca-Marques, E. W. Heymann EW und K. B. Strier (Hrsg.): South American primates: comparative perspectives in the study of behavior, ecology and conservation. New York, Springer, 2009, S. 69–113
- Richard Kay, Abraham Lincoln, Matthew Colbert und Michael Malinzak: Brain size, activity pattern, and visual acuity in Homunculus patagonicus, an Early Miocene stem platyrrhine: the mosaic evolution of brain size and visual acuity in Anthropoidea. Journal of Vertebrate Paleontology 26 (3suppl.), 2006, S, 83A
- Mark N. Coleman, Richard F. Kay und Matthew W. Colbert: Auditory Morphology and Hearing Sensitivity in Fossil New World Monkeys. The Anatomical Record 293, 2010, S. 1711–1721
- Mark N. Coleman und Matthew W. Colbert: Correlations Between Auditory Structures and Hearing Sensitivity in Non-Human Primates. Journal of Morphology 271, 2010, S. 511–532
- Daniele Silvestro, Marcelo F. Tejedor, Martha L. Serrano-Serrano, Oriane Loiseau, Victor Rossier, Jonathan Rolland, Alexander Zizka, Sebastian Höhna, Alexandre Antonelli und Nicolas Salamin: Early Arrival and Climatically-Linked Geographic Expansion of New World Monkeys from Tiny African Ancestors. Systematic Biology 68 (1), 2019, S. 78–92
- Laurent Marivaux, Sylvain Adnet, Ali J. Altamirano-Sierra, Myriam Boivin, François Pujos, Anusha Ramdarshan, Rodolfo Salas-Gismondi, Julia V. Tejada-Lara und Pierre-Olivier Antoine: Neotropics provide insights into the emergence of New World monkeys: New dental evidence from the late Oligocene of Peruvian Amazonia. Journal of Human Evolution 97, 2016, S. 159–175
- Horacio Schneider und Iracilda Sampaio: The systematics and evolution of New World primates – A review. Molecular Phylogenetics and Evolution 82, 2015, S. 348–357, doi:10.1016/j.ympev.2013.10.017
- F. Dumas und S. Mazzoleni: Neotropical primate evolution and phylogenetic reconstruction using chromosomal data: Italian Journal of Zoology 84 (1), 2017, S. 1–18, doi:10.1080/11250003.2016.1260655
- Xiaoping Wang, Burton K. Lim, Nelson Ting, Jingyang Hu, Yunpeng Liang, Christian Roos und Li Yu: Reconstructing the phylogeny of new world monkeys (platyrrhini): evidence from multiple non-coding loci. Current Zoology, 2018, S. 1–10, doi:10.1093/cz/zoy072
- Mariano Bond, Marcelo F. Tejedor, Kenneth E. Campbell Jr., Laura Chornogubsky, Nelson Novo und Francisco Goin: Eocene primates of South America and the African origins of New World monkeys. Nature 520 (7548), 2015, S. 538–541
- Florentino Ameghino: Enumération synoptique des espèces de mammifères fossiles des formations éocènes de Patagonia. Buenos Aires, 1894, S. 1–196 (S. 9–11 und 102) ()
- Richard F. Kay: Biogeography in deep time – What do phylogenetics, geology, and paleoclimate tell us about early platyrrhine evolution? Molecular Phylogenetics and Evolution 82, 2015, S. 358–374
- Alfred L. Rosenberger: Evolutionary Morphology, Platyrrhine Evolution, and Systematics. The Anatomical Record 294, 2011, S. 1955–1974
- Marcelo F. Tejedor: Sistemática, evolución y paleobiogeografía de los primates Platyrrhini. Revista del Museo de La Plata Sección Zoología 20 (176), 2013, S. 20–39
- Alfred L. Rosenberger: Platyrrhines, PAUP, parallelism, and the Long Lineage Hypothesis: A reply to Kay et al. (2008). Journal of Human Evolution 59, 2010, S. 214–217
- Jessica W. Lynch Alfaro, Liliana Cortés-Ortiz, Anthony Di Fiore und Jean P. Boubli: Comparative biogeography of Neotropical primates. Molecular Phylogenetics and Evolution 82, 2015, S. 518–529
- S. Ivan Perez, Marcelo F. Tejedor, Nelson M. Novo und Leandro Aristide: Divergence Times and the Evolutionary Radiation of New World Monkeys (Platyrrhini, Primates): An Analysis of Fossil and Molecular Data. PLoS ONE 8 (6), 2013, S. e68029, doi:10.1371/journal.pone.0068029
- Jason A. Hodgson, Kirstin N. Sterner, Luke J. Matthews, Andrew S. Burrell, Rachana A. Jani, Ryan L. Raaum, Caro-Beth Stewart und Todd R. Disotell: Successive radiations, not stasis, in the South American primate fauna. PNAS 106 (14), 2009, S. 5534–5539
- Florentino Ameghino: Nuevos restos de mamiferos fosiles decubiertos par Carlos Ameghino en el eoceno inferior de la Patagonia austral. - Especies nuevas, adiciones y correcciones. Revista Argentina de Historia Natural 1, 1891, S. 289–328 ()
- Florentino Ameghino: Los monos fósiles del eoceno de la república Argentina. Revista Argentina de Historia Natural 1, 1891, S. 383–397 ()
- Alcides Mercerat: Sobre la presencia de restos de monos en el eoceno de Patagonia. Revista del Museo de La Plata 2, 1891, S. 73–74 ()
- Carlos Rusconi: Nuevos restos de monos fósiles del Terciario antiguo de la Patagonia. Anales de la Sociedad Científica Argentina 116, 1933, S. 286–289 ()
- Philip Hershkovitz: A new genus of Late Oligocene monkey (Cebidae, Platyrrhini) with notes on postorbital closure and platyrrhine evolution. Folia Primatologica 21, 1974, S. 1–35
- John G. Fleagle: New fossil platyrrhines from the Pinturas Formation, southern Argentina. Journal of Human Evolution 19, 1990, S. 61–85