Eionaletherium
Eionaletherium ist eine ausgestorbene Gattung der Faultiere, die bisher nur über einige Elemente der Hinterbeine und anderer Teile des Körperskelettes eines einzigen Individuums nachgewiesen ist. Diese stammen aus der Urumaco-Formation des nordwestlichen Venezuelas, welche in das Obere Miozän vor rund 10 bis 7 Millionen Jahren datiert. Die Gesteinsformation besteht aus Ablagerungen, die auf ein Flussdelta in unmittelbarer Nähe zur damaligen Meeresküste zurückgehen. Die Faultiergattung wird als naher Verwandter der Gruppe der Mylodontidae eingestuft. Als Besonderheit besaß sie abweichend von anderen Mylodonten einen sehr langen unteren Beinabschnitt. Ob dies eine Anpassung an ein Leben in einem eher wässerigen Milieu darstellt, ist bisher aufgrund der wenigen Funde nicht eindeutig. Die wissenschaftliche Erstbeschreibung von Eionaletherium erfolgte im Jahr 2015.
| Eionaletherium | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
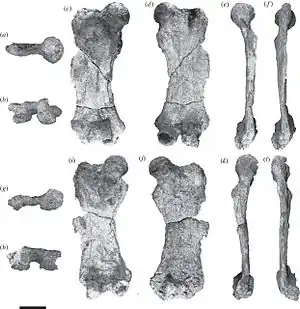
rechter und linker Oberschenkelknochen von Eionaletherium, Teil des Holotyps | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberes Miozän | ||||||||||||
| 10,1 bis 7 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Eionaletherium | ||||||||||||
| Rincón, McDonald, Solórzano, Flores & Ruiz-Ramoni, 2015 | ||||||||||||
Merkmale
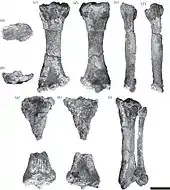
Eionaletherium war ein mittelgroßer Vertreter aus der Verwandtschaftsgruppe der Mylodontidae. Er ist bisher über einige Elemente der Hinterbeine, einzelne Wirbel und Rippen und über beide Schulterblätter bekannt. Anhand der Größe der Funde, hauptsächlich des Oberschenkelknochens, wird auf ein Tier mit einem Körpergewicht von rund 1,1 t geschlossen. Der Oberschenkelknochen erreichte eine Länge von 51 cm. Er war relativ breit und typisch für Mylodonten brettartig flach. Darin weicht er von den Femora der Megatheriidae ab, deren Querschnitt leicht gewölbt erschien. Der Schaft besaß einen eher geraden Verlauf und zeigte dadurch nur eine leichte Drehung, weniger als vergleichbar bei Pseudoprepotherium. Beide Seitenkanten waren auffällig nach innen gewölbt. Der Kopf des Oberschenkelknochens hob sich deutlich durch einen kurzen Hals ab. Zwischen diesem und dem Großen Rollhügel befand sich eine leichte, sattelartige Eindellung. Der Große Rollhügel ragte wie bei Pseudoprepotherium nicht über die Basis des Kopfansatzes hinaus. Dadurch war er niedriger als etwa bei Magdalenabradys. Der relativ kleine dritte Trochanter (dritter Rollhügel) lag seitlich unterhalb des Großen Rollhügels, etwas oberhalb der Mitte des Schaftes. Diese Position war im Vergleich zu Pseudoprepotherium und Magdalenabradys auffällig höher, bei letzterem war er zusätzlich noch weiter nach hinten versetzt und zudem deutlich größer ausgeprägt. Die beiden unteren, körperfernen Gelenkrollen wiesen eine asymmetrische Gestaltung auf, so dass die innere größer als die äußere war. Ungewöhnlich für Mylodonten erreichte das Schienbein mit einer Länge von 44 cm fast 87 % der Länge des Oberschenkelknochens. In der Regel liegt dieser Wert bei unter 65 %, bei einigen späten Vertretern der Faultiergruppe wie Glossotherium oder Lestodon sogar bei 50 % und weniger. Der Schaft des Schienbeins war oval geformt und hatte ebenfalls einen geraden Verlauf. Die oberen und unteren Gelenkenden wiesen in etwa die gleiche Breite auf. Das Wadenbein war nicht mit dem Schienbein verwachsen, das Merkmal ist aber recht unterschiedlich bei den Mylodonten ausgeprägt.[1]
Fundstelle
Alle bisher bekannten Funde von Eionaletherium wurden in der Urumaco-Formation im venezolanischen Bundesstaat Falcón im Nordwesten des Landes aufgefunden. Die Urumaco-Formation ist Teil der Urumaco-Sequenz, einer Abfolge von drei lithostratigraphischen Einheiten, der Socorro-, der Urumaco- und der Codore-Formation, die einen Teil des rund 36.000 km² großen Falcón-Beckens bedecken. Die drei Gesteinsformationen datieren in den Zeitraum vom Mittleren Miozän bis zum Unteren Pliozän, wobei die Urumaco-Formation dem Oberen Miozän angehört und ein Alter von etwa 10 bis 7 Millionen Jahren besitzt. Sie ist beidseitig des Río Urumaco aufgeschlossen und erreicht eine Mächtigkeit von 1700 m. Die vorherrschenden Ablagerungen stellen verschiedene Sand-, Ton-/Schluff- und Kalksteine sowie einzelnen Kohlelagen dar. Informell wird die Urumaco-Formation in drei Untereinheiten untergliedert, dem Unteren, Mittleren und Oberen Schichtglied. Das Mittlere Schichtglied, aus dem die Funde stammen, ist bis zu 755 m mächtig. Es besteht aus einer Abfolge von Kalksteinen, mächtigen, graufarbenen Ton-/Schluffsteinen zuzüglich fein geschichteten, mit organischen Resten angereicherten Ton-/Schluffsteinen und Kohleflözen mit zahlreichen Wirbeltierresten. Diese Abfolge wird teilweise durch rinnenartige Strukturen von 10 bis lokal 20 m Mächtigkeit unterbrochen, in denen feinkörnige, kreuzgeschichtete Sandsteine abgelagert sind, die zumeist eine Abtragungsfläche überdecken. Diese Sandsteine werden nach oben hin feinkörniger und tragen dann häufig Rippelstrukturen. Die gesamte Ablagerungssequenz geht auf einen küstennahen Bereich zurück, der unter dem Einfluss eines Flussdeltas stand. Dabei geben die unterschiedlichen Sedimentgesteine das jeweilige Vorherrschen mariner (Kalk- und Ton-/Schluffsteine) oder fluviatiler (Sandsteine) Bedingungen im Laufe der einzelnen Ablagerungsphasen wieder.[2]
Die Urumaco-Sequenz birgt eine hohe Anzahl an Fossilfunden. Die ersten Reste wurden bereits Anfang der 1950er Jahre bei Erkundungen nach Erdöl entdeckt. Daraufhin erfolgten gegen Ende des Jahrzehnts erste wissenschaftliche Expeditionen in das Gebiet. Die intensive Erforschung über die nächsten Dekaden führten dazu, dass heute über 100 Fundstellen bekannt sind, die sich auf 60 unterschiedliche stratigraphische Ebenen verteilen. Das Fundmaterial kann zu einem großen Teil den Fischen zugewiesen werden, unter denen wiederum Haie und Rochen dominieren. Daneben sind auch Reptilien wie Schildkröten, Krokodile sowie vereinzelt Schlangen nachgewiesen, hinzu kommt auch eine umfangreiche Säugetierfauna. Unter den Säugetieren treten unter anderem Nagetiere, Südamerikanische Huftiere, Seekühe und Nebengelenktiere in Erscheinung. Dabei erreichen vor allem die Nebengelenktiere eine äußerst hohe Diversität, die der aus weitaus südlicheren Gebieten Südamerikas, etwa die Pamparegion oder Mesopotamia, dieser Zeit kaum nachsteht. So sind aus der Gruppe der Gepanzerten Nebengelenktiere Vertreter der drei großen Hauptlinien, die Gürteltiere, die Pampatheriidae und die Glyptodontidae, nachgewiesen. Unter den Zahnarmen dominieren die Faultiere.[3][4][5] Hier konnten Ende des 20. und zu Beginn des 21. Jahrhunderts zahlreiche neue Gattungen aus der Urumaco-Sequenz beschriebenen werden. Aus der Urumaco-Formation stammen dabei unter anderem Urumacocnus und Pattersonocnus aus der Gruppe der Megalonychidae, Urumaquia als ein Vertreter der großen Megatheriidae sowie Magdalenabradys, Pseudoprepotherium, Bolivartherium und Urumacotherium aus der Linie der Mylodontidae und deren unmittelbaren Verwandten. Als ein besonderer Umstand der Taphonomie ist die häufige Überlieferung von Gliedmaßenelementen bei den Faultieren zu bewerten.[6][7][8][9] Innerhalb der Faultiere der Urumaco-Formation stellt Eionaletherium einen eher kleineren Vertreter dar. Andere Mylodonten werden auf ein Gewicht von 1,7 t für Magdalenabradys bis 2,1 t für Urumacotherium geschätzt. In der Größe ähnelte es eher dem Megatherien-Angehörigen Urumaquia.[1]
Paläobiologie
Besonders auffällig ist das gegenüber dem Oberschenkelknochen sehr lange Schienbein, was bei Mylodonten so nicht bekannt ist. Das Schienbein erreicht rund 87 % der Länge des Oberschenkelknochens. Dieser Wert entspricht etwa dem von Thalassocnus aus der Gruppe der Nothrotheriidae (nur einzelne Angehörige der Megatheriidae besitzen ein noch ausgewogeneres Verhältnis der beiden Knochen). Bei Thalassocnus handelt es sich um ein semi-aquatisch lebendes Tier, das zahlreiche anatomische Besonderheiten aufweist. Der verlängerte untere Beinabschnitt wird hier als Anpassung an paddelnde Bewegungen im Wasser interpretiert, da dadurch die Hebelwirkung beim Schwimmen erhöht und der Vortrieb erleichtert wird.[10] Prinzipiell könnten die geologischen Bedingungen der Urumaco-Formation auch für eine stärkere Adaption von Eionaletherium an ein wässriges Milieu sprechen und so das verhältnismäßig lange Schienbein erklären. Allerdings fehlen der Faultiergattung weitere Anpassungen, wie sie bei Thalassocnus vorkommen. So sind bei Eionaletherium keine Knochenschwellungen (Pachyostose) und Knochenverdichtungen (Osteosclerose) nachgewiesen, die häufig bei sekundär an ein Wasserleben angepasste Landwirbeltiere auftreten und den Auftrieb im Wasser bremsen. Ebenso fehlt bei Eionaletherium am Oberschenkelkopf die Hüftkopfgrube (Fovea capitis femoris), was für schwach ausgebildete Bänder und somit nur eine geringe Abspreizfähigkeit des Beines spricht und untypisch für semi-aquatische Tiere ist. Aufgrund dieser Befunde muss Eionaletherium vorerst als rein bodenbewohnendes Lebewesen angesehen werden.[1]
Systematik
Mögliche innere Systematik der Mylodontoidea nach Rincón et al. 2020[9] basierend auf Merkmalen der Hinterbeine
|
Eionaletherium ist eine ausgestorbene Gattung aus der Unterordnung der Faultiere (Folivora). Die Faultiere umfassen heute nur zwei Gattungen kleiner, baumlebender Tiere, in ihrer stammesgeschichtlichen Vergangenheit waren sie sehr formenreich und brachten mehrere Linien zumeist bodenbewohnender Vertreter hervor. Eine der bedeutendsten Linien stellen nach klassischer skelettanatomisch begründeter Ansicht die Mylodontoidea dar, eine sehr diverse Gruppe bestehend aus den Mylodontidae, den Scelidotheriidae und den Orophodontidae (letztere beiden werden teilweise auch als Unterfamilien innerhalb der Mylodontidae geführt[11]). Die zweite große Entwicklungslinie findet sich mit den Megatherioidea. Molekulargenetische Studien unterstützt durch Proteinanalysen erfassen noch eine dritte Großgruppe, die Megalocnoidea. Die Mylodontoidea stellen mit den Zweifinger-Faultieren (Choloepus) auch eine der beiden heute noch existierenden Gattungen.[12][13]
Aufgrund der Merkmale der hinteren Gliedmaßen, etwa dem extrem abgeplatteten Oberschenkelknochen und dessen seitlich eingedellten Schaftverlauf, steht Eionaletherium den Mylodontoidea nahe. Dabei wies die Gattung phylogenetischen Untersuchungen aus dem Jahr 2015 zufolge nähere Beziehungen zu Formen wie Glossotherium und weiteren entwickelten Mylodonten auf als zu anderen Angehörigen aus der Überfamilie. Möglicherweise könnte dadurch Eionaletherium auch direkt den Mylodonten zugesprochen werden, wobei unklar ist, ob es aufgrund der langen Unterbeine eine besonders basale oder weiter abgeleitete Form darstellt.[1] Eine neuere Studie aius dem Jahr 2020 konnte dies weitgehend bestätigen.[9]
Anders dagegen sieht dies eine Studie aus dem Jahr 2019, die Eionaletherium aus den Mylodonten ausgegliedert.[14] Problematisch bei der Untersuchung zu den stammesgeschichtlichen Verwandtschaftsverhältnissen ist, dass sie in diesem Fall auf Merkmalen der Hinterbeine beruht. Dadurch erscheinen Vertreter wie Hapalops oder Analcimorphus als basale Formen der Mylodontoidea, während Analysen des Schädels und der Zähne sie als solche der Megatherioidea ausweisen.[15] Es ist wahrscheinlich, dass die Morphologie der Hinterbeine bei sehr ursprünglichen Vertretern der Megatherioidea und Mylodontoidea sehr ähnlich war. Für eine genauere Beurteilung muss daher aussagekräftiges Schädelmaterial mit einbezogen werden, dass von Eionaletherium aber bisher nicht vorliegt.[1]
Entdeckungsgeschichte
Die Fossilreste wurden bei Untersuchungen des Instituto Venezolano de Investigaciones Científicas (IVIC) in bis dahin unerforschten Bereichen der Urumaco-Formation entdeckt. Sie entstammen der Fundstelle „Charlie“ im Bezirk Buchivacoa des venezolanischen Bundesstaat Falcón. Die Sedimente, in denen die Funde eingebettet waren, hatten sich unter niederenergetischen Fließbedingungen des Wassers an der Küste abgelagert. Da alle Knochen von Eionaletherium auf einer Fläche von nur 2 m² aufgefunden wurden und der Bereich ansonsten nur noch Schildkröten- und Krokodil-Reste enthielt, werden sie als zu einem Individuum gehörig betrachtet. Die wissenschaftliche Erstbeschreibung erfolgte im Jahr 2015 durch Ascanio D. Rincón und Forscherkollegen. Der Holotyp (Exemplarnummer IVIC-P-2870) umfasst beide Oberschenkelknochen, das vollständige rechte Schienbein mit Wadenbein, das fragmentierte linke Schienbein, Reste beider Schulterblätter, einige Wirbel und Rippen sowie ein Sprungbein. Die Funde werden in Caracas aufbewahrt. Der Gattungsname Eionaletherium setzt sich aus den griechischen Wörtern ἠϊών (eion „Küste“, „Ufer“), αλη (ale „Umherschweifen“, „Umherirren“) und θηρίον (thērion „Tier“) zusammen. Er bezieht sich auf die Paläolandschaft, in der das Faultier einst lebte. Die einzige anerkannte Art ist E. tanycnemius. Der Artname tanycnemius wiederum besteht aus den griechischen Begriffen τανη (tany „lang“) und κνημἱς (knemis „vom Knie bis zum Knöchel reichend“) und ist eine Referenz auf das lang gestreckte Schienbein.[1]
Literatur
- Ascanio D. Rincón, H. Gregory McDonald, Andrés Solórzano, Mónica Núñez Flores und Damián Ruiz-Ramoni: A new enigmatic Late Miocene mylodontoid sloth from northern South America. Royal Society Open Science 2, 2015, S. 140256, doi:10.1098/rsos.140256.
Einzelnachweise
- Ascanio D. Rincón, H. Gregory McDonald, Andrés Solórzano, Mónica Núñez Flores und Damián Ruiz-Ramoni: A new enigmatic Late Miocene mylodontoid sloth from northern South America. Royal Society Open Science 2, 2015, S. 140256, doi:10.1098/rsos.140256
- Luis I. Quiroz und Carlos A. Jaramillo: Stratigraphy and sedimentary environments of Miocene shallow to marginal marine deposits in theUrumaco trough,Falcón Basin, Western Venezuela. In: Marcelo R. Sánchez-Villagra, Orangel A. Aguilera und Alfredo A. Carlini (Hrsg.): Urumaco and Venezuelan palaeontology, the fossil record of the northern Neotropics. Indiana University Press 2010, S. 153–172
- Omar J. Linares: Biostratigrafia de la fauna de mammiferos de las formaciones Socorro, Urumaco y Codore (Mioceno Medio-Plioceno Temprano) de la region de Urumaco, Falcón, Venezuela. Paleobiologia Neotropical 1, 2004, S. 1–26
- Marcelo R. Sánchez-Villagra, Orangel A. Aguilera: Neogene vertebrates from Urumaco, Falcón State, Venezuela: diversity and significance. Journal of Systematic Palaeontology 4, 2006, S. 213–220
- Marcelo R. Sánchez-Villagra, Orangel A. Aguilera, Rodolfo Sánchez und Alfredo A. Carlini: The fossil vertebrate record of Venezuela of the last 65 Million years. In: Marcelo R. Sánchez-Villagra, Orangel A. Aguilera und Alfredo A. Carlini (Hrsg.): Urumaco and Venezuelan palaeontology, the fossil record of the northern Neotropics. Indiana University Press 2010, S. 19–51
- Alfredo A. Carlini, Diego Brandoni und Rodolfo Sánchez: First Megatherines (Xenarthra, Phyllophaga, Megatheriidae) from the Urumaco (late Miocene) and Codore (Pliocene) formations, Estado Falcón, Venezuela. Journal of Systematic Palaeontology 4, 2006, S. 269–278
- Alfredo A. Carlini, Gustavo J. Scillato-Yané und Rodolfo Sánchez: New Mylodontoidea (Xenarthra, Phyllophaga) from the middle Miocene–Pliocene of Venezuela. Journal of Systematic Palaeontology 4, 2006, S. 255–267
- Ascanio D. Rincon, Andres Solorzano, H. Gregory McDonald und Marisol Montellano-Ballesteros: Two new megalonychid sloths (Mammalia: Xenarthra) from the Urumaco Formation (late Miocene), and their phylogenetic affinities. Journal of Systematic Palaeontology 17 (5), 2018, S. 409–421, doi:10.1080/14772019.2018.1427639
- Ascanio D. Rincón und H. Gregory McDonald: Reexamination of the Relationship of Pseudoprepotherium Hoffstetter, 1961, to the Mylodont Ground Sloths (Xenarthra) from the Miocene of Northern South America. Revista Geológica de América Central 63, 2020, S. 1–20, doi:10.15517/rgac.v62i0.41278
- Eli Amson, Christine Argot, H. Gregory McDonald und Christian de Muizon: Osteology and Functional Morphology of the Hind Limb of the Marine Sloth Thalassocnus (Mammalia, Tardigrada). Journal of Mammal Evolution 2014, doi:10.1007/s10914-014-9274-5
- Luciano Varela, P. Sebastián Tambusso, H. Gregory McDonald und Richard A. Fariña: Phylogeny, Macroevolutionary Trends and Historical Biogeography of Sloths: Insights From a Bayesian Morphological Clock Analysis. Systematic Biology 68 (2), 2019, S. 204–218
- Frédéric Delsuc, Melanie Kuch, Gillian C. Gibb, Emil Karpinski, Dirk Hackenberger, Paul Szpak, Jorge G. Martínez, Jim I. Mead, H. Gregory McDonald, Ross D.E. MacPhee, Guillaume Billet, Lionel Hautier und Hendrik N. Poinar: Ancient mitogenomes reveal the evolutionary history and biogeography of sloths. Current Biology 29 (12), 2019, S. 2031–2042, doi:10.1016/j.cub.2019.05.043
- Samantha Presslee, Graham J. Slater, François Pujos, Analía M. Forasiepi, Roman Fischer, Kelly Molloy, Meaghan Mackie, Jesper V. Olsen, Alejandro Kramarz, Matías Taglioretti, Fernando Scaglia, Maximiliano Lezcano, José Luis Lanata, John Southon, Robert Feranec, Jonathan Bloch, Adam Hajduk, Fabiana M. Martin, Rodolfo Salas Gismondi, Marcelo Reguero, Christian de Muizon, Alex Greenwood, Brian T. Chait, Kirsty Penkman, Matthew Collins und Ross D. E. MacPhee: Palaeoproteomics resolves sloth relationships. Nature Ecology & Evolution 3, 2019, S. 1121–1130, doi:10.1038/s41559-019-0909-z
- Alberto Boscaini, François Pujos und Timothy J. Gaudin: A reappraisal of the phylogeny of Mylodontidae (Mammalia, Xenarthra) and the divergence of mylodontine and lestodontine sloths. Zoologica Scripta 48 (6), 2019, S. 691–710, doi:10.1111/zsc.12376
- Timothy J. Gaudin: Phylogenetic relationships among sloths (Mammalia, Xenarthra, Tardigrada): the craniodental evidence. Zoological Journal of the Linnean Society 140, 2004, S. 255–305