Diplodocus
Diplodocus („Doppelbalken“; altgriechisch διπλόος diplóos „doppelt“ und δοκός dokós „Balken“)[2] ist eine Gattung von sauropoden Dinosauriern aus dem Oberjura (Kimmeridgium bis Tithonium) des westlichen Nordamerikas.
| Diplodocus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettabguss von Diplodocus im Berliner Hauptbahnhof. | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberjura (Kimmeridgium bis Tithonium)[1] | ||||||||||||
| 157,3 bis 145 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Diplodocus | ||||||||||||
| Marsh, 1878 | ||||||||||||
Der Gattungsname bezieht sich auf die V-förmigen Chevron-Knochen auf der Unterseite der Schwanzwirbelsäule. Anfangs hielt man die V-förmigen Fortsätze für eine Autapomorphie von Diplodocus, sie sind seit der Erstbeschreibung jedoch auch bei anderen Diplodociden entdeckt worden.
Diplodocus war eine der häufigeren Gattungen der ca. 156 bis 147 Millionen Jahre alten Oberen Morrison-Formation und gehörte einer von riesigen Sauropoden wie Camarasaurus, Barosaurus, Apatosaurus und Brachiosaurus dominierten Fauna an.[3] Die Dinosauriergattung ist eine der am einfachsten zu identifizierenden und galt für viele Jahre als diejenige mit der größten Körperlänge. Seine enorme Größe mag Schutz vor Raubsauriern wie Allosaurus und Ceratosaurus geboten haben.
Beschreibung
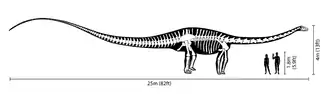
Diplodocus ist einer der am besten bekannten Sauropoden. Das sehr große, quadrupede Tier besaß einen langen Hals und einen langen, peitschenartigen Schwanz. Dass die Vorderbeine etwas kürzer waren als die Hinterbeine, führte zu einer horizontalen Körperhaltung, so dass Diplodocus angesichts des langen Halses, Schwanzes und der stämmigen Beine mechanisch einer Hängebrücke glich.[4] Tatsächlich wurde der Gattung Diplodocus das längste vollständige Dinosaurierskelett zugeordnet.[4] Manche Sauropoden wie Seismosaurus (dessen Skelettfunde eventuell große Vertreter der Gattung Diplodocus repräsentieren) und Supersaurus erreichten vermutlich noch größere Körperlängen als Diplodocus, sind jedoch nur fragmentarisch fossil belegt.[5] Von den bis zu 27 m Körperlänge entfielen 6 Meter auf den Hals. Der Schädel war in Relation zur Körpergröße sehr klein.[6] Diplodocus besaß kleine, stiftförmige Zähne, die nur den vorderen Teil der Kiefer ausfüllten und von der Zahnwurzel schräg nach vorn gerichtet waren.[7] Seine Schädelkapsel war klein. Der Hals bestand aus mindestens fünfzehn Halswirbeln und man nimmt derzeit an, dass das Tier ihn meist parallel zum Grund hielt und kaum darüber hinaus anheben konnte.[8] Schätzungen zum Körpergewicht variieren, aktuelle Studien geben 10,[9] 11,5,[10] 12,7[11] oder 16 Tonnen[12] an.

Diplodocus besaß einen extrem langen Schwanz, bestehend aus rund 80 Schwanzwirbeln,[13] was fast dem doppelten der Schwanzwirbelzahl mancher frühen Sauropoden (wie Shunosaurus: 43) entsprach und die zur gleichen Zeit lebenden Vertreter der Macronaria (Camarasaurus: 53) bei weitem übertraf. Es gibt Spekulationen darüber, inwiefern der Schwanz eher eine direkte Verteidigungsfunktion hatte[14] oder der (Abschreckung durch) Lärmerzeugung diente.[15]
Der Schwanz könnte auch als Gegengewicht zum massigen Hals gedient haben. Die namensgebenden „Doppelbalken“ der Chevron-Knochen treten im Mittelteil des Schwanzes als Fortsätze an der Unterseite auf. Sie könnten die Last des Schwanzes mitgetragen oder, im Falle dass der Schwanz auf dem Boden auflag, die Blutgefäße vor dem Zerdrücktwerden geschützt haben. In mehreren verwandten Gattungen kommen „Doppelbalken“ ebenfalls vor.
Paläobiologie
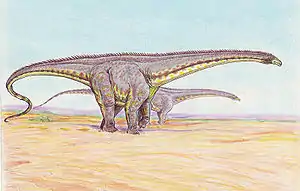
Dank des Reichtums an Skelettresten zählt Diplodocus zu den am besten untersuchten Dinosauriern. Viele Aspekte der Lebensweise sind in den Jahren seit der Erstbeschreibung der Gegenstand verschiedener Theorien geworden.
Lebensraum
Othniel Charles Marsh und später John Bell Hatcher[16] nahmen aufgrund der Lage der Nasenöffnungen an der Schädelspitze an, dass es sich um ein aquatisches Tier handele. Ähnliche Auffassungen flossen oftmals in die zeichnerische Darstellung anderer großer Sauropoden wie Brachiosaurus und Apatosaurus ein.
Die Idee einer aquatischen Lebensweise mit dem langen Hals als „Schnorchel“ wurde später verworfen, da der hydrostatische Druck des Wassers auf den Brustkorb für das Atmen zu groß gewesen wäre. Seit den 1970er Jahren besteht weitgehend Konsens über eine streng terrestrische Lebensweise der Sauropoden als Baumabweider. Später wurde die vorhergehende Ansicht, Diplodocus könnte möglicherweise Wasserpflanzen bevorzugt haben, wieder aufgegriffen – in der Idee, Diplodocus habe einen ufernahen Lebensraum bevorzugt, hallt die ursprüngliche Theorie einer aquatischen Lebensweise nach.
Körperhaltung


Die Darstellung der Körperhaltung von Diplodocus hat sich über die Jahre beträchtlich verändert. Beispielsweise zeigt die klassische Rekonstruktion von Oliver P. Hay (1910) zwei Individuen der Gattung mit gespreizten eidechsenartigen Beinen am Ufer eines Flusses. Hay argumentierte für eine gestreckte eidechsenartige Gangart mit weit gespreizten Beinen[18] und wurde darin von Gustav Tornier unterstützt. Diese Hypothese wurde jedoch von W. J. Holland widerlegt, der zeigt, dass Diplodocus als Spreizgänger einen Graben benötigt hätte, um seinen Bauch mitzuschleifen.[19]
Später wurden Diplodociden oft mit hoch aufgerichteten Hälsen dargestellt, was sie zum Abweiden hoher Bäume befähigt hätte. Von dieser Ansicht kam man jedoch wieder ab, da das Herz nicht problemlos in der Lage gewesen wäre, einen ausreichend hohen Blutdruck zu erzeugen, um das Gehirn mit Sauerstoff zu versorgen. Zudem zeigen aktuellere Studien, dass die Struktur der Halswirbel keine steile Aufrichtung des Halses erlaubte.[20][21]
Wie bei der verwandten Gattung Barosaurus ist der sehr lange Hals von Diplodocus eine Quelle vieler Kontroversen unter Wissenschaftlern. Eine Studie der Columbia University zur Halsstruktur der Diplodociden aus dem Jahr 1993 hatte zum Ergebnis, dass die längsten Hälse ein 1,6 Tonnen schweres Herz benötigt hätten, um das Gehirn mit Sauerstoff zu versorgen. Demzufolge ergäbe sich die Notwendigkeit rudimentärer „Hilfsherzen“ entlang des Halses, um kaskadenartig Blut in den Kopf zu pumpen.[4]
Während lange Hälse traditionell als Anpassung an eine bestimmte Ernährungsstrategie interpretiert werden, schlägt eine Studie von 2006 vor, dass der überlange Hals von Diplodocus und seiner Verwandten primär der Zurschaustellung als Teil des Paarungsverhaltens gedient haben könnte und Vorteile bei Ernährung möglicherweise zunächst nur nachrangig von Bedeutung waren.[22]
Ernährung
Verglichen mit anderen Sauropoden besaß Diplodocus sehr ungewöhnliche Zähne: Die Zahnkronen waren lang und schmal, elliptisch im Querschnitt, während die Spitze eine abgestumpft-dreieckige Form besaß.[7] Die deutlichsten Abnutzungsspuren zeigen die Zahnkronenspitzen. Im Unterschied zu allen anderen Abnutzungsmustern, die man bisher bei Sauropoden beobachtet hat, zeigt Diplodocus Abnutzung auf der Wangenseite sowohl der unteren als auch der oberen Zahnreihe.[7] Demnach besaßen Diplodocus und andere Diplodociden einen Fressmechanismus, der sich deutlich von dem anderer Sauropoden unterschied – einseitiges Abstreifen von Ästen stellt für Diplodocus die wahrscheinlichste Fressweise dar[23][24][25], da nur dieser Mechanismus die ungewöhnlichen Abnutzungsmuster als Folge des Kontakts der Zähne mit der Nahrung erklären kann. Beim einseitigen Abstreifen der Zweige hätte eine Zahnreihe dazu gedient, das Laub vom Stamm abzulösen, während die jeweils andere Zahnreihe als Führung und Stabilisator gedient hätte. Durch die verlängerte Präorbitalregion (d. h. der vor den Augen gelegene Bereich) des Schädels konnten längere Abschnitte eines Zweigs während einer Fressbewegung entlaubt werden.[7] Auch die rückwärtige Bewegung der Unterkiefer könnte auf zweierlei Weise zum Fressvorgang beigetragen haben: 1) Vergrößerung des Abstands zwischen den Zahnreihen; 2) Feinjustierung der relativen Lage der Zahnreihen und ihrer Bewegung zueinander, was zu einem gleichmäßigeren Abweidevorgang geführt hätte.[7]
Mit einem flexiblen Hals, der zur Seite sowie auf und ab bewegt werden konnte, und der Möglichkeit, den Schwanz als Stütze beim Aufrichten auf die Hinterbeine zu nutzen, dürfte Diplodocus die Fähigkeit besessen haben, in verschiedenen Höhen bis maximal 10 m über dem Boden zu weiden.[26]
Interessanterweise hätte der Bewegungsradius des Halses auch ein Grasen bis unterhalb des Körperniveaus erlaubt, was manche Forscher zu der Spekulation veranlasste, Diplodocus könnte sich auch von Unterwasserpflanzen entlang von Flussufern ernährt haben. Für dieses Konzept der Fresshaltung spricht das Längenverhältnis der Vorder- und Hintergliedmaßen. Zudem scheinen die stiftförmigen Zähne durchaus an das Fressen weicher Wasserpflanzen angepasst zu sein.[20]
Andere Aspekte der Anatomie
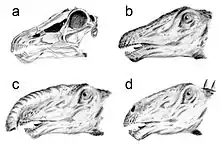
a Schädel
b klassische Darstellung des Kopfes mit hochgelegenen Nüstern
c Bakkers Rüsselhypothese
d tiefe Nüsternposition nach Witmer 2001
Der Kopf von Diplodocus wird aufgrund der Lage der Nasenöffnungen an der Spitze des Schädels meistens mit Nüstern in entsprechender Lage oberhalb der Augen dargestellt. Es gibt Spekulationen darüber, ob die Morphologie des Schädels auch bedeuten könnte, dass Diplodocus einen Rüssel besaß.[27] Eine Studie von 2006[28] legt dar, dass es keine paläoneurologischen Belege für einen Rüssel gibt. Während der Gesichtsnerv bei Rüsselträgern wie Elefanten zur Steuerung des Rüssels relativ groß ist, scheint der des Diplodocus sehr klein gewesen zu sein. Untersuchungen von Lawrence Witmer (2001) deuten an, dass trotz der hohen Lage der Nasenöffnungen im Schädel die Nüstern weitaus tiefer in Richtung der Schnauze gelegen haben könnten.[29]
Kürzlich entdeckte Fossilien legen nahe, dass Diplodocus und andere Diplodociden eine Reihe niedriger, spitz zulaufender keratinöser Stacheln entlang der Rückenlinie ähnlich denen der Leguane besaßen.[30][31] Diese Erkenntnis ist in die jüngeren Lebendrekonstruktionen, unter anderem in die TV-Serie Walking with Dinosaurs, eingeflossen. Jedoch ist ungewiss, wie viele der Diplodociden dieses Merkmal aufweisen und ob es auch bei anderen Sauropoden vorhanden war.
Wachstumsrate
Laut einer Anzahl knochenhistologischer Untersuchungen zeigen Diplodocus und eine Anzahl weiterer Sauropoden sehr hohe Wachstumsraten und erreichten die Geschlechtsreife in etwas mehr als einem Jahrzehnt, obwohl sich das Größenwachstum dennoch (langsamer) bis zum Lebensende fortsetzte.[32][33][34] Vormals meinte man, dass Sauropoden während ihrer gesamten Lebenszeit nur langsam wuchsen und erst nach Jahrzehnten geschlechtsreif und ausgewachsen waren.
Fortpflanzung
Während es keine Belege für die Brutgewohnheiten von Diplodocus gibt, konnte man andere Sauropoden wie den Titanosaurier Saltasaurus mit Gelegen in Verbindung bringen.[35][36] Diese Gelege zeigen, dass Titanosaurier ihre Eier gemeinschaftlich, verteilt über ein großes Gebiet, in vielen flachen Gruben ablegten und mit Vegetation bedeckten. Möglicherweise tat Diplodocus dasselbe.
Entdeckungsgeschichte und Arten

Mehrere Arten von Diplodocus sind zwischen 1878 und 1924 beschrieben worden. Das erste Skelett wurde 1878 von Benjamin Franklin Mudge und Samuel Wendell Williston in Como Bluff, Wyoming entdeckt und von dem Paläontologen Othniel Charles Marsh noch im selben Jahr als Typusart wissenschaftlich erstbeschrieben und Diplodocus longus („langer Doppelbalken“) genannt.[37] Überreste von Diplodocus sind seither in der Morrison-Formation in den westlichen US-Bundesstaaten Colorado, Utah, Montana und Wyoming entdeckt worden. Mit Ausnahme des Schädels, der auch oft bei ansonsten vollständigen Skeletten fehlt, sind Fossilien dieses Dinosauriers häufig. Obwohl es sich nicht um die Typusart handelt, hat D. carnegiei aufgrund der Vielzahl von Abgüssen des nahezu vollständigen Originalskeletts, die in Museen in der gesamten Welt ausgestellt sind, eine größere Bekanntheit erlangt.
Die Sauropodengattungen Diplodocus und Barosaurus, die beide aus der Morrison-Formation stammen, besaßen sehr ähnliche Gliedmaßenknochen. In der Vergangenheit wurden viele isolierte Gliedmaßenknochen darum einfach Diplodocus zugeschrieben, obwohl sie möglicherweise zu Barosaurus gehören.[38]
Gültige Arten
- D. longus, die Typusart, beruht auf zwei Schädeln und einer Schwanzwirbelsäule aus der Morrison-Formation von Colorado und Utah.[39]
- D. carnegii[40][41] (in älterer Literatur auch D. carnegiei)[42], benannt nach Andrew Carnegie, ist die am besten bekannte Art, vor allem aufgrund eines nahezu vollständigen Skeletts, das von Jacob Wortman vom Carnegie Museum of Natural History in Pittsburgh, Pennsylvania entdeckt und im Jahr 1901 von John Bell Hatcher wissenschaftlich beschrieben und benannt wurde.
- D. hayi beruht auf einem Teilskelett, das von William H. Utterback im Jahr 1902 nahe Sheridan, Wyoming entdeckt und 1924 beschrieben wurde.[43] Die Art wurde 2015 bei einer Revision der Diplodocidae in die neu errichtete Gattung Galeamopus gestellt.[44]
- D. hallorum ist auch bekannt als Seismosaurus hallorum. Ein Vortrag[45] auf der Jahrestagung der Geologischen Gesellschaft von Amerika im Jahr 2004 stellte plausibel dar, dass Seismosaurus ein mutmaßliches jüngeres Synonym von Diplodocus repräsentiert. Seismosaurus hallorum wurde im Jahr 1979 entdeckt und 1991 als eigene Gattung beschrieben.
Zweifelhafte Arten
- D. lacustris ist ein nomen dubium. Die Art wurde von Othniel Marsh im Jahr 1884 beschrieben. Die Beschreibung erfolgte auf der Grundlage von Knochenresten eines kleineren Individuums, die in Morrison, Colorado gefunden wurden.[46] Diese Fossilien werden heute als Überreste eines Jungtiers und nicht für Vertreter einer eigenen Art gedeutet.
Klassifikation
Diplodocus ist sowohl die Typus-Gattung als auch der Namensgeber der Familie Diplodocidae.[46] Die Mitglieder der Familie sind beträchtlich schlanker gebaut als Titano- und Brachiosaurier. Sie sind charakterisiert durch lange Hälse und Schwänze und eine horizontale Haltung. Ihre Vorderbeine sind kürzer als die Hinterbeine. Diplodociden hatten ihre Blütezeit im Oberen Jura. In dieser Zeit waren sie in Nordamerika und Afrika häufig.[47] Sie wurden während der Kreidezeit von den Titanosauriern abgelöst, die ähnliche ökologische Nischen belegten.
Die Unterfamilie Diplodocinae umfasst Diplodocus und seine nächsten Verwandten, darunter Seismosaurus, der zur selben Gattung gehören könnte, und Barosaurus. Der zeitgleich im Fossilbericht erscheinende Apatosaurus scheint mit Diplodocus ferner verwandt zu sein – die Gattung wird zur selben Familie gezählt, jedoch in die Unterfamilie Apatosaurinae gestellt.[48][49]
Der portugiesische Dinheirosaurus und die afrikanische Gattung Tornieria sind von manchen Autoren ebenfalls als nahe Verwandte von Diplodocus klassifiziert worden.[50][51]
Die Überfamilie Diplodocoidea umfasst die Diplodociden sowie Dicraeosauriden, Rebbachisauriden und die Gattung Suuwassea,[48][49] Amphicoelias[51] und möglicherweise Haplocanthosaurus,[52] und/oder die Nemegtosauriden.[6] Dieses Taxon bildet die Schwestergruppe zu Camarasaurus, den Brachiosauriden und den Titanosauriern (die zur Gruppe Macronaria zusammengefasst werden).[6][52] Diplodocoidea und Macronaria bilden zusammen die Neosauropoda, welche die größte, diverseste und erfolgreichste Gruppe der sauropodomorphen Dinosaurier darstellt.
Darstellung in der allgemeinen Öffentlichkeit

Diplodocus ist eine der allgemein bekannten und oft dargestellten Dinosauriergattungen, wahrscheinlich aufgrund der Vielzahl an Skelettfunden und des Status als längster Dinosaurier. Die Vergabe vieler Skelettabgüsse an Museen weltweit trug dazu bei, Menschen vieler Kulturen mit Diplodocus in Kontakt zu bringen und vertraut zu machen. Diplodocus-Skelette werden noch immer in vielen Museen ausgestellt: D. hayi im Houston Museum of Natural Science, D. carnegiei im Natural History Museum in London, im Naturwissenschaftlichen Museum in Madrid, Spanien, im Senckenberg-Museum in Frankfurt, Deutschland, im Field Museum of Natural History in Chicago and natürlich im Carnegie Museum of Natural History in Pittsburgh. Ein montiertes Skelett von D. longus ist im Smithsonian Museum of Natural History in Washington, D. C. zu besichtigen
Diplodocus war Protagonist einer Episode der TV-Serie der BBC Dinosaurier – Im Reich der Giganten. Unter dem Titel „Zeit der Titanen“ wurde der Werdegang eines Individuums vom Ei bis zum Tod und der Fossilisation nachgestellt.
Einzelnachweise
- Gregory S. Paul: The Princeton Field Guide To Dinosaurs. Princeton University Press, Princeton NJ u. a. 2010, ISBN 978-0-691-13720-9, S. 190.
- Henry George Liddell, Robert Scott: A Lexicon abridged from Liddell and Scott's Greek-English Lexicon. 24th edition, carefully revised throughout. Ginn, Boston 1891 (Reprinted edition. Clarendon Press, Oxford 2010, ISBN 978-0-19-910207-5).
- Christine E. Turner, Fred Peterson: Reconstruction of the Upper Jurassic Morrison Formation extinct ecosystem – a synthesis. In: Sedimentary Geology. Bd. 167, Nr. 3/4, 2004, ISSN 0037-0738, S. 309–355, doi:10.1016/j.sedgeo.2004.01.009.
- David Lambert: The Ultimate Dinosaur Book. RD Press in association with The Natural History Museum London, Surry Hills NSW 1993, ISBN 0-86438-417-3.
- Mathew J. Wedel, Richard L. Cifelli: Sauroposeidon: Oklahoma's Native Giant. In: Oklahoma Geology Notes. Bd. 65, Nr. 2, 2005, ISSN 0030-1736, S. 39–57, Digitalisat (PDF; 2,15 MB).
- Paul Upchurch, Paul M. Barrett, Peter Dodson: Sauropoda. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 259–324, hier S. 316.
- Paul Upchurch, Paul M. Barrett: The evolution of sauropod feeding mechanisms. In: Hans-Dieter Sues (Hrsg.): Evolution of herbivory in terrestrial vertebrates. Perspectives from the fossil record. Cambridge University Press, Cambridge u. a. 2000, ISBN 0-521-59449-9, S. 79–122.
- Kent A. Stevens, J. Michael Parrish: Neck Posture and Feeding Habits of Two Jurassic Sauropod Dinosaurs. In: Science. Bd. 284, Nr. 5415, 1999, S. 798–800, doi:10.1126/science.284.5415.798.
- Peter Dodson, Anke K. Behrensmeyer, Robert T. Bakker, John S. McIntosh: Taphonomy and paleoecology of the dinosaur beds of the Jurassic Morrison Formation. In: Paleobiology. Bd. 6, 1980, ISSN 0094-8373, S. 208–232, Digitalisat.
- Gregory S. Paul: Big Sauropods – Really, Really Big Sauropods. In: The Dinosaur Report. Fall 1994, S. 12–13, Digitalisat (PDF; 2,12 MB).
- John R. Foster: Paleoecological analysis of the vertebrate fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain region, U.S.A. (= New Mexico Museum of Natural History and Science. Bulletin. 23, ISSN 1524-4156). New Mexico Museum of Natural History, Albuquerque NM 2003, Digitalisat.
- M. J. Coe, David L. Dilcher, James O. Farlow, David M. Jarzen, Dale A. Russell: Dinosaurs and land plants. In: Else Marie Friis, William G. Chaloner, Peter R. Crane (Hrsg.): The Origins of Angiosperms and their biological Consequences. Cambridge University Press, Cambridge u. a. 1987, ISBN 0-521-32357-6, S. 225–258.
- Jeffrey A. Wilson: Overview of Sauropod Phylogeny and Evolution. In: Kristina Curry Rogers, Jeffrey A. Wilson (Hrsg.): The Sauropods. Evolution and Paleobiology. University of California Press, Berkeley CA u. a. 2005, ISBN 0-520-24623-3, S. 15–49.
- William J. Holland: Heads and Tails; a few notes relating to the structure of sauropod dinosaurs. In: Annals of the Carnegie Museum. Bd. 9, Nr. 3/4, Article 16, 1915, ISSN 0097-4463, S. 273–278, Digitalisat.
- Nathan P. Myhrvold, Philip J. Currie: Supersonic sauropods? Tail dynamics in the diplodocids. In: Paleobiology. Bd. 23, Nr. 4, 1997, S. 393–409, Digitalisat.
- John B. Hatcher: Diplodocus (Marsh), Its Osteology, Taxonomy, and Probable Habits, with a Restoration of the Skeleton. In: Memoirs of the Carnegie Museum. Bd. 1, Nr. 1, 1901, ISSN 0885-4645, S. 1–64, Digitalisat.
- Oliver P. Hay: On the Manner of Locomotion of the Dinosaurs especially Diplodoctus, with Remarks on the Origin of the Birds. In: Proceedings of the Washington Academy of Sciences. Bd. 12, Nr. 1, 1910, ISSN 0363-1095, S. 1–25, Digitalisat.
- Oliver P. Hay: On the Habits and Pose of the Sauropod Dinosaurs, especially of Diplodocus. In: The American Naturalist. Bd. 42, Nr. 502, 1908, S. 672–681, Digitalisat.
- William J. Holland: A Review of Some Recent Criticisms of the Restorations of Sauropod Dinosaurs Existing in the Museums of the United States, with Special Reference to that of Diplodocus carnegiei in the Carnegie Museum. In: The American Naturalist. Bd. 44, Nr. 521, 1910, S. 259–283, Digitalisat.
- Kent A. Stevens, J. Michael Parrish: Neck Posture, Dentition, and Feeding Strategies in Jurassic Sauropod Dinosaurs. In: Virginia Tidwell, Kenneth Carpenter (Hrsg.): Thunder-lizards. The Sauropodomorph dinosaurs. Indiana University Press, Bloomington IN u. a. 2005, ISBN 0-253-34542-1, S. 212–232.
- Paul Upchurch: Neck Posture of Sauropod Dinosaurs. In: Science. Bd. 287, Nr. 5453, 2000, S. 547b, doi:10.1126/science.287.5453.547b, Digitalisat (PDF; 169,82 KB).
- Phil Senter: Necks for Sex: Sexual Selection as an Explanation for Sauropod Neck Elongation. In: Journal of Zoology. Bd. 271, Nr. 1, 2007, ISSN 0952-8369, S. 45–53, doi:10.1111/j.1469-7998.2006.00197.x.
- David Norman: The illustrated Encyclopedia of Dinosaurs. Salamander Books, London 1985, ISBN 0-86101-225-9.
- Peter Dodson: Sauropod paleoecology. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. University of California Press, Berkeley CA u. a. 1990, ISBN 0-520-06726-6, S. 402–507.
- Paul M. Barrett, Paul Upchurch: Feeding mechanisms of Diplodocus. In: Martin B. Lockley, Vanda Faria dos Santos, Christian A. Meyer, Adrian P. Hunt (Hrsg.): Aspects of Sauropod Paleobiology (= Gaia. Revista de Geociências. Bd. 10, ISSN 0871-5424). Museu Nacional de História Natural, Lisboa 1994, S. 195–204, Digitalisat (PDF; 2,18 MB).
- Paul M. Barrett, Paul Upchurch: Sauropodomorph Diversity through Time: Paleoecological and Macroevolutionary Implications. In: Kristina Curry Rogers, Jeffrey A. Wilson (Hrsg.): The Sauropods. Evolution and Paleobiology. University of California Press, Berkeley CA u. a. 2005, ISBN 0-520-24623-3, S. 125–156.
- Robert T. Bakker: The Dinosaur Heresies. New Theories Unlocking the Mystery of the Dinosaurs and their Extinction. Morrow, New York NY 1986, ISBN 0-688-04287-2.
- Fabien Knoll, Peter M. Galton, Raquel López-Antoñanzas: Paleoneurological evidence against a proboscis in the sauropod dinosaur Diplodocus. In: Geobios. Bd. 39, Nr. 2, 2006, ISSN 0016-6995, S. 215–221, doi:10.1016/j.geobios.2004.11.005.
- Lawrence M. Witmer: Nostril Position in Dinosaurs and other Vertebrates and its Significance for Nasal Function. In: Science. Bd. 293, Nr. 5531, 2001, S. 850–853, doi:10.1126/science.1062681.
- Stephen A. Czerkas: Discovery of dermal spines reveals a new look for sauropod dinosaurs. In: Geology. Bd. 20, Nr. 12, 1992, ISSN 0091-7613, S. 1068–1070, doi:10.1130/0091-7613(1992)020<1068:DODSRA>2.3.CO;2.
- Stephen Czerkas: The history and interpretation of sauropod skin impressions. In: Martin B. Lockley, Vanda Faria dos Santos, Christian A. Meyer, Adrian P. Hunt (Hrsg.): Aspects of Sauropod Paleobiology (= Gaia. Revista de Geociências. Bd. 10). Museu Nacional de História Natural, Lisboa 1994, S. 173–182, Digitalisat (PDF; 2,69 MB).
- P. Martin Sander: Longbone histology of the Tendaguru sauropods: implications for growth and biology. In: Paleobiology. Bd. 26, Nr. 3, 2000, S. 466–488, doi:10.1666/0094-8373(2000)026<0466:LHOTTS>2.0.CO;2.
- P. Martin Sander, Nicole Klein, Eric Buffetaut, Gilles Cuny, Varavudh Suteethorn, Jean Le Loeuff: Adaptive radiation in sauropod dinosaurs: bone histology indicates rapid evolution of giant body size through acceleration. In: Organisms Diversity & Evolution. Bd. 4, Nr. 3, 2004, ISSN 1439-6092, S. 165–173, doi:10.1016/j.ode.2003.12.002.
- P. Martin Sander, Nicole Klein: Developmental plasticity in the life history of a prosauropod dinosaur. In: Science. Bd. 310, Nr. 5755, 2005, S. 1800–1802, doi:10.1126/science.1120125.
- Luis M. Chiappe, Lowell Dingus: Walking on Eggs. The Astonishing Discovery of Thousands of Dinosaur Eggs in the Badlands of Patagonia. Scribner, New York NY 2001, ISBN 0-7432-1211-8.
- Gerald Grellet-Tinner, Luis M. Chiappe, Rodolfo Coria: Eggs of titanosaurid sauropods from the Upper Cretaceous of Auca Mahuevo (Argentina). In: Canadian Journal of Earth Sciences. Bd. 41, Nr. 8, 2004, ISSN 0008-4077, S. 949–960, doi:10.1139/e04-049.
- Othniel C. Marsh: Principal characters of American Jurassic dinosaurs. Part I. In: American Journal of Science. Bd. 116 = Series 3, Bd. 16, Nr. 95, Article 44, 1878, ISSN 0002-9599, S. 411–416, Digitalisat.
- John S. McIntosh: The Genus Barosaurus Marsh (Sauropoda, Diplodocidae). In: Virginia Tidwell, Kenneth Carpenter (Hrsg.): Thunder-lizards. The Sauropodomorph dinosaurs. Indiana University Press, Bloomington IN u. a. 2005, ISBN 0-253-34542-1, S. 38–77.
- Paul Upchurch, Paul M. Barrett, Peter Dodson: Sauropoda. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 259–324, hier S. 305.
- Carnegie Museum of Natural History, Diplodocus carnegii (Memento vom 28. Juni 2013 im Internet Archive)
- so auch in David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2.
- Othenio Abel: Die Rekonstruktion des Diplodocus (= Abhandlungen der Zoologisch-Botanischen Gesellschaft in Wien. Bd. 5, Heft 3, ISSN 0255-0458). Fischer, Jena 1910, Digitalisat (PDF; 8 MB).
- William J. Holland: The skull of Diplodocus. In: Memoirs of the Carnegie Museum. Bd. 9, Nr. 3, 1924, S. 379–403.
- Emanuel Tschopp, Octávio Mateus, Roger B.J. Benson. A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda). PeerJ, 2015; 3: e857 DOI: 10.7717/peerj.857
- Spencer G. Lucas, Matthew C. Herne, Andrew B. Heckert, Adrian P. Hunt, Robert M. Sullivan: Reappraisal of Seismosaurus, A Late Jurassic Sauropod Dinosaur from New Mexico. In: Geological Society of America Abstracts with Programs. Bd. 36, Nr. 5, 2004, ISSN 0016-7592, S. 422, Abstract.
- Othniel C. Marsh: Principal characters of American Jurassic dinosaurs. Part VII. On the Diplodocidæ, a new family of the Sauropoda. In: American Journal of Science. Bd. 127 = Series 3, Bd. 27, Nr. 98, Article 21, 1884, S. 161–168, Digitalisat.
- J. A. Wilson: The Sauropods:Evolution and Paleobiology. Hrsg.: K. A. Rogers & J. A. Wilson. Indiana University Press, 2005, ISBN 0-520-24623-3, S. 15–49.
- Michael P. Taylor, Darren Nash: The phylogenetic taxonomy of Diplodocoidea (Dinosauria: Sauropoda). In: PaleoBios. Bd. 25, Nr. 2, 2005, ISSN 0031-0298, S. 1–7, online.
- Jerald D. Harris: The significance of Suuwassea emilieae (Dinosauria: Sauropoda) for flagellicaudatan intrarelationships and evolution. In: Journal of Systematic Palaeontology. Bd. 4, Nr. 2, 2006, ISSN 1477-2019, S. 185–198, doi:10.1017/S1477201906001805.
- José F. Bonaparte, Octavio Mateus: A new diplodocid, Dinheirosaurus lourinhanensis gen. et sp. nov., from the Late Jurassic beds of Portugal. In: Revista del Museo Argentino de Ciencias Naturales Bernardino Rivadavia e Instituto Nacional de Investigación de las Ciencias Naturales. Paleontología. Bd. 5, Nr. 2, 1999, ISSN 0524-9511, S. 13–29, online.
- Oliver W. M. Rauhut, Kristian Remes, Regina Fechner, Gerardo Cladera, Pablo Puerta: Discovery of a short-necked sauropod dinosaur from the Late Jurassic period of Patagonia. In: Nature. Bd. 435, Nr. 7042, 2005, S. 670–672, doi:10.1038/nature03623.
- Jeffrey A. Wilson: Sauropod dinosaur phylogeny: critique and cladistic analysis. In: Zoological Journal of the Linnean Society. Bd. 136, Nr. 2, 2002, ISSN 0024-4082, S. 215–275, doi:10.1046/j.1096-3642.2002.00029.x.