Ceratosaurus
Ceratosaurus („Horn-Echse“) ist eine Gattung theropoder Dinosaurier aus dem Oberjura von Nordamerika und Europa. Charakteristisch für diesen zweibeinigen Fleischfresser waren drei markante Hörner auf dem Schädel sowie eine Reihe kleiner Osteoderme (Hautknochenplatten), die über Hals, Rücken und Schwanz verlief. Ceratosaurus ist vor allem durch fünf fragmentarische Skelette mit Schädel bekannt, die aus der Morrison-Formation der westlichen USA stammen. Er ist der namensgebende Vertreter der Ceratosauria, einer Gruppe basaler (ursprünglicher) Theropoden. Die einzige derzeit allgemein anerkannte Art ist Ceratosaurus nasicornis. Es wurden zwei weitere Arten beschrieben, Ceratosaurus dentisulcatus und C. magnicornis, deren Gültigkeit heute jedoch umstritten ist. Ceratosaurus zählt zu den in der Öffentlichkeit bekannten Dinosauriern und taucht regelmäßig in populären Büchern über Dinosaurier auf.[2]
| Ceratosaurus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettabguss eines Jungtieres im Dinosaur Discovery Museum in Wisconsin | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberjura (Kimmeridgium bis frühes Tithonium)[1] | ||||||||||||
| 157,3 bis 147,7 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Ceratosaurus | ||||||||||||
| Marsh, 1884 | ||||||||||||
| Arten | ||||||||||||
|


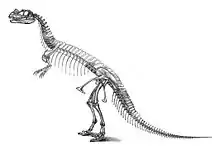



Merkmale
Das Holotyp-Exemplar von Ceratosaurus nasicornis wird auf eine Körperlänge von 5,3 Metern geschätzt[3], wobei unklar ist, ob dieses Individuum vollständig ausgewachsen war. Das größte gefundene Exemplar stammt aus dem Cleveland-Lloyd-Quarry und wird auf eine Länge von 8,8 Meter geschätzt;[2] dieses Exemplar wird von einigen Forschern als eigenständige Art, Ceratosaurus dentisulcatus, geführt.[4] Schätzungen des Körpergewichts variieren je nach Autor, so belaufen sich verschiedene Gewichtsschätzungen des Holotyp-Exemplars auf 418,4 kg, 524 kg und 670 kg.[5]
Der Schädel war wie bei anderen Ceratosauria verhältnismäßig groß.[6] Einzigartig ist ein markantes Horn, das sich auf der Mittellinie des Schädels hinter den Nasenöffnungen befindet. Fossil überliefert ist lediglich die knöcherne Hornbasis, die beim lebenden Tier mit Keratin überzogen war. Die Hornbasis ist rundlich und schmal. Beim Holotyp-Exemplar hat sie an ihrer Basis eine Länge von 13 Zentimeter und eine Breite von 2 Zentimeter; unmittelbar oberhalb der Basis nimmt die Breite jedoch auf 1,2 Zentimeter ab. Ihre Höhe beträgt 7 Zentimeter, bezogen auf die Oberseite des Nasenbeins.[7] Die Hornbasis wird von den miteinander verschmolzenen rechten und linken Hälften des paarigen Nasenbeins (Nasale) gebildet. Bei Jungtieren sind die beiden Hälften der Hornbasis noch nicht verschmolzen[8]. Zusätzlich zu dem Nasenhorn besaß Ceratosaurus ein Paar Hörner, das sich auf der Oberseite des Schädels vor den Augenöffnungen befindet und vom paarigen Tränenbein (Lacrimale) gebildet wird. Alle drei Hörner waren bei erwachsenen Tieren im Verhältnis größer als bei Jungtieren.[9] Die Hörner dienten wahrscheinlich ausschließlich der Zurschaustellung[10].
In jeder Hälfte des Oberkiefers saßen zwischen 12 und 15 klingenförmige Zähne. Das paarige Zwischenkieferbein (Prämaxillare), ein Knochen vor dem Oberkiefer, wies auf jeder Hälfte nur drei Zähne auf, weniger als bei den meisten anderen Theropoden[11]. Im Unterkiefer saßen auf jeder Seite 11 bis 15 Zähne, die etwas gerader und weniger kräftig waren als die Zähne des Oberkiefers.[12] Die Zahnkronen des Oberkiefers waren außergewöhnlich lang und messen beim größten bekannten Ceratosaurus-Exemplar bis zu 9,3 cm Länge, was der geringsten Höhe des Unterkiefers entspricht. Beim kleineren Holotyp-Exemplar übertrifft die Länge der Zahnkronen (7 cm) sogar die geringste Höhe des Unterkiefers (6,3 cm) – dieses Merkmal findet sich bei anderen Theropoden lediglich bei dem möglicherweise nahe verwandten Genyodectes.[13] Im Kontrast dazu wiesen einige Vertreter der ebenfalls mit Ceratosaurus verwandten Abelisauridae sehr niedrige Zahnkronen auf.[14]
Die Anzahl der Wirbel lässt sich nicht genau bestimmen, da die Wirbelsäule des Holotyp-Exemplars mehrere Lücken aufweist. Das Kreuzbein besteht aus 6 Kreuzbeinwirbeln (Sakralwirbel); vor dem Kreuzbein lagen mindestens 20 Präsakralwirbel (Hals- und Rückenwirbel), während hinter dem Kreuzbein etwa 50 Schwanzwirbel folgten. Die Dornfortsätze der Schwanzwirbel waren ebenso wie die Chevronknochen verlängert, was dem Schwanz ein von der Seite betrachtet tiefes Profil gab.[15] Einzigartig unter Theropoden ist eine Reihe kleiner, länglicher und unregelmäßig geformter Osteoderme (Hautknochenplatten), die auf der Mittellinie des Körpers oberhalb der Wirbel auf Hals, Rücken und dem Großteil des Schwanzes verlief. Abseits der Mittellinie des Körpers wies Ceratosaurus andere Osteoderme auf, worauf eine etwa 6 mal 7 Zentimeter große Platte hinweist, die zusammen mit dem Holotyp-Exemplar gefunden wurde, deren Position auf dem Körper des Tieres jedoch unklar ist.[16]
Die Arme waren wie bei anderen Vertretern der Ceratosauria verhältnismäßig kurz und endeten in vier Fingern. Bisher ist lediglich ein unvollständiges Handskelett gefunden worden, weshalb die exakte Anzahl der Fingerglieder unklar ist. Der erste und vierte Mittelhandknochen waren in ihrer Größe reduziert, während der zweite Mittelhandknochen etwas länger war als der dritte.[17][18]
Forschungsgeschichte und Funde
Das erste Exemplar (Holotyp, Exemplarnummer USNM 4737) besteht aus einem relativ vollständigen Skelett mit Schädel und wurde von dem Farmer Marshall P. Felch in den Jahren 1883 bis 1884 entdeckt[19]. Es stammt aus dem Felch Quarry 1, einem der reichhaltigsten Steinbrüche der Morrison-Formation,[20] im Garden-Park-Gebiet nördlich von Cañyon City in Colorado.[3] Der Fund wurde aus hartem Sandstein freigelegt, Schädel und Wirbelsäule sind stark zerdrückt vorgefunden worden. Aus demselben Steinbruch wurde – neben zahlreichen weiteren Dinosaurier-Fossilien – zuvor bereits das Holotyp-Exemplar von Allosaurus geborgen. 1884 beschrieb Othniel Charles Marsh auf Basis dieses Skeletts eine neue Gattung und Art, Ceratosaurus nasicornis[21]. Dank der relativ vollständigen Überreste war Ceratosaurus zu dieser Zeit der am besten bekannte jurassische Theropode Amerikas. Der Name Ceratosaurus (gr. keras – „Horn“, sauros – „Echse“) bedeutet so viel wie „Hornechse“ und soll auf das markante Nasenhorn hinweisen[22].[2][3] 1920 veröffentlichte Charles Gilmore eine umfangreiche Neubeschreibung des Skeletts[23].
Eine erste Skelettrekonstruktion veröffentlichte Marsh 1892. Wie Gilmore (1920) anmerkte, zeigt die Rückenwirbelsäule dieser Rekonstruktion mindestens sechs überzählige Wirbel, was in einem deutlich zu langen Rumpf resultiert. Dieser Fehler wurde in verschiedenen Veröffentlichungen der Folgezeit übernommen, so in der ersten Lebendrekonstruktion, die Frank Bond 1899 unter Anleitung von Charles Knight anfertigte, jedoch erst 1920 veröffentlicht wurde. Eine korrektere Lebendrekonstruktion wurde von J. M. Gleeson erstellt, ebenfalls unter Anleitung von Charles Knight, und 1901 veröffentlicht. Das Holotyp-Skelett wurde in den Jahren 1910 und 1911 von Gilmore montiert und ist seitdem im National Museum of Natural History in Washington, D.C. zu sehen. Da das Skelett seitlich stark zerdrückt ist, montierte Gilmore das Skelett als Basrelief (Flachrelief). Frühe Rekonstruktionen zeigen Ceratosaurus meist in aufrechter Haltung und mit einem auf dem Boden aufliegenden Schwanz. Gilmores Montage dagegen war seiner Zeit voraus[24] – inspiriert durch den in stark angewinkelter Position vorgefundenen Oberschenkelknochen stellte Gilmore die Montage als laufendes Tier dar, mit waagerechter Körperhaltung und einem nicht den Boden berührenden Schwanz.[3]
Nach der Entdeckung des Holotyp-Exemplars wurden anfangs nur sehr fragmentarische Überreste von Ceratosaurus gefunden. Erst in den frühen 1960er Jahren wurde unter der Leitung von James Madsen ein fragmentarisches Skelett mit Schädel (Exemplarnummer UMNH VP 5278) im Cleveland-Lloyd-Quarry in Utah geborgen – es handelt sich dabei um das größte bisher bekannte Ceratosaurus-Exemplar. Anders als das Holotyp-Exemplar ist dieses Skelett nicht artikuliert vorgefunden worden; die Knochen befanden sich bei der Entdeckung also nicht mehr in ihrer ursprünglichen anatomischen Position.[25] Ein weiteres, artikuliertes Skelett mit Schädel (Exemplarnummer MWC 1) entdeckte Thor Erikson, der Sohn des Paläontologen Lance Erikson, 1976 nahe der Stadt Fruita in Colorado.[2] Dieses Exemplar ist relativ vollständig, wichtige fehlende Skelettelemente schließen die Unterkiefer, Unterarme und Bauchrippen (Gastralia) mit ein. Es handelte sich um ein großes Individuum, das jedoch nicht vollständig ausgewachsen war, worauf zahlreiche offene Schädelsuturen hinweisen[26]. Beide Skelette wurden von Madsen und Welles (2000) in ihrer Revision der Gattung beschrieben, wobei das in Utah entdeckte Skelett den Holotypus der neuen Art Ceratosaurus dentisulcatus und das in Colorado entdeckte den der neuen Art Ceratosaurus magnicornis bildet.[4] 1992 wurde ein weiteres, fragmentarisches Skelett (Exemplarnummer BYUVP 12893) im Agate-Basin-Quarry in Utah entdeckt; die Beschreibung wird derzeit vorbereitet. Dieses Exemplar schließt einen vollständigen Gesichtsschädel, sieben fragmentarische Rückenwirbel und fragmentarische Beckenknochen mit ein. Es handelt sich um eines der beiden größten bekannten Skelette von Ceratosaurus.[27] 1999 berichtet Brooks Britt über die Entdeckung des bislang einzigen Ceratosaurus-Skeletts, das zu einem Jungtier gehörte. Dieses Skelett stammt aus dem Bone-Cabin-Quarry in Wyoming. Es ist um 34 % kleiner als das Holotyp-Exemplar und besteht aus einem vollständigen Schädel sowie dem zu 30 % erhaltenen Postkranium einschließlich eines vollständigen Beckens.[8]
Neben den fünf Skelettfunden sind weitere fragmentarische Funde aus verschiedenen bedeutenden Fundorten bekannt. So stammt aus dem Dinosaur National Monument in Utah stammt ein isoliert vorgefundenes, rechtes Zwischenkieferbein (Prämaxillare, Exemplarnummer DNM 972). Aus Como Bluff in Wyoming wurde der Fund eines großen Schulterblatts mit Rabenbein gemeldet. Auch aus dem Dry-Mesa-Quarry in Colorado stammt ein noch unbeschriebener, aus einem linken Schulterblatt mit Rabenbein sowie Fragmenten von Wirbeln und Gliedmaßen bestehender Fund. Im Mygatt-Moore-Quarry in Colorado ist die Gattung durch Zähne vertreten.[27]
Nachweise außerhalb Nordamerikas
In den Jahren 1909 bis 1913 brachten Expeditionen des Berliner Museum für Naturkunde die diverse Dinosaurierfauna der Tendaguru-Formation im heutigen Tansania zu Tage.[28] Die Tendaguru-Formation gilt heute als der wichtigste Fundort für Dinosaurierfossilien in Afrika,[28] obwohl große Theropoden lediglich durch wenige und sehr fragmentarische Funde überliefert sind[29]. 1920 schrieb Werner Janensch einige Rückenwirbel aus dem Steinbruch TL der Gattung Ceratosaurus zu, ohne sie einer bestimmten Art zuzuordnen (Ceratosaurus sp.). 1925 beschrieb Janensch eine neue Ceratosaurus-Art, Ceratosaurus roechlingi, basierend auf fragmentarischen Resten aus dem Steinbruch Mw, die ein Quadratbein, ein Wadenbein, Reste von Schwanzwirbeln und weitere Fragmente umfassen. Dieser Fund gehörte zu einem Tier, das deutlich größer war als das Holotyp-Exemplar von Ceratosaurus[29]. Madsen und Welles (2000) bestätigen die Zuordnung beider Funde zu Ceratosaurus und schreiben einige Zähne, die von Janensch 1920 als Labrosaurus (?) stechowi beschrieben wurden, ebenfalls Ceratosaurus zu.[27] Andere Autoren widersprechen jedoch und geben an, dass keine der aus Tendaguru stammenden Funde diagnostische Merkmale zeigt, die eine Zuordnung zu Ceratosaurus rechtfertigen würden[9][30][31]. Rowe und Gauthier (1990) erwähnen eine zweite Ceratosaurus-Art aus Tendaguru, Ceratosaurus ingens, die angeblich von Janensch (1920) aufgestellt worden sei und auf 25 isoliert vorgefundenen und mit bis zu 15 cm Länge sehr großen Zähnen basiert[31][32]. Tatsächlich hat Janensch diese Art jedoch der Gattung Megalosaurus zugeschrieben, weshalb es sich bei diesem Namen möglicherweise um einen Abschreibfehler handelt[27][32].
Oktávio Mateus und Kollegen (2000, 2006) schreiben Ceratosaurus einen Knochenfund, bestehend aus einem Oberschenkelknochen und einem Schienbein, sowie verschiedene isoliert vorgefundene Zähne zu, die aus der Lourinhã-Formation in Portugal stammen.[33][34] Diese Forscher kommen zu dem Ergebnis, dass sich die portugiesische Theropodenfauna ähnlich zusammensetzt wie die der nordamerikanischen Morrison-Formation. So konnten neben Ceratosaurus zwei weitere für die Morrison-Formation typische Gattungen auch in Portugal nachgewiesen werden, Allosaurus und Torvosaurus. Europa war zur Zeit des Oberjura durch den erst jungen und schmalen Atlantik von Nordamerika getrennt, die Iberische Halbinsel war noch von anderen Teilen Europas getrennt. Die Ähnlichkeit der Theropodenfaunen Portugals und Nordamerikas lassen jedoch auf zeitweilig bestehende Landbrücken schließen, die einen Faunenaustausch ermöglichten.[33] Diese Autoren schreiben den Knochenfund der Art Ceratosaurus dentisulcatus zu[33]. Carrano und Sampson (2008) bestätigen zwar die Zuordnung dieser Fossilien zu Ceratosaurus, kommen jedoch zu dem Ergebnis, dass der Fund keiner bestimmten Art zugeordnet werden kann.[30]
Aus Moutier (Schweiz) stammt ein einzelner Zahn, der ursprünglich von Janensch (1920) als Labrosaurus meriani beschrieben, jedoch von Madsen und Welles (2000) als Ceratosaurus sp. bezeichnet wurde.[27] Soto und Perea (2008) beschreiben Zähne aus der Tacuarembó-Formation Uruguays, die Merkmale zeigen, die für die Gattung Ceratosaurus diagnostisch sind. Eine Zuordnung dieser Zähne zu Ceratosaurus ist laut diesen Forschern jedoch wegen des nur spärlichen Fossilmaterials ausgeschlossen. Auch die Zuordnungen von Fossilien aus Europa und Afrika zur Gattung Ceratosaurus seien laut diesen Forschern wegen des spärlichen Fossilmaterials mit Vorsicht zu betrachten.[35]
Systematik
Arten
Die Typusart und derzeit einzige allgemein anerkannte Art ist Ceratosaurus nasicornis. Madsen und Welles (2000) stellten zwei neue Arten auf: Ceratosaurus magnicornis aus Colorado und Ceratosaurus dentisulcatus aus Utah. Während Ceratosaurus magnicornis ein etwas runderes Horn zeigt als Ceratosaurus nasicornis, ist das Horn bei Ceratosaurus dentisulcatus nicht fossil überliefert. Beide Arten waren deutlich größer als Ceratosaurus nasicornis. Die Gültigkeit dieser Arten ist umstritten: Britt und Kollegen (2000) vermuten, dass es sich bei dem Holotyp-Exemplar von Ceratosaurus nasicornis tatsächlich um ein Jungtier handelte, und dass die beiden größeren Arten tatsächlich adulte Stadien von Ceratosaurus nasicornis repräsentieren.[9] Rauhut (2003) und Carrano und Sampson (2008) halten die beiden neuen Arten ebenfalls für identisch mit Ceratosaurus nasicornis und geben an, dass es sich bei den von Madsen und Welles beschriebenen Unterschieden wahrscheinlich um ontogenetische (entwicklungsspezifische) oder individuelle Variationen handelte.[36][30]
Äußere Systematik
| Kladogramm der Ceratosauria (vereinfacht nach Xu, 2009)[37] | |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||
| Systematische Position von Ceratosaurus. |
Die Gruppe Ceratosauria beinhaltet die Gruppe Abelisauroidea, welche alle abgeleiteten (fortgeschrittenen) Vertreter wie beispielsweise Carnotaurus und Noasaurus umfasst, sowie eine Reihe von basalen (ursprünglichen) Vertretern, darunter Ceratosaurus, Elaphrosaurus, Spinostropheus und Deltadromeus.[38] Die meisten Studien sehen Ceratosaurus als die abgeleitetste dieser basalen Gattungen und als Schwestertaxon der Abelisauroidea.[38][37] Oliver Rauhut (2004) vermutet jedoch, dass der lediglich durch Kieferknochen bekannte Genyodectes das Schwestertaxon von Ceratosaurus darstellt; beide Gattungen fasst er als Ceratosauridae zusammen.[39] Die Gruppe Ceratosauridae wurde bereits 1884 von Othniel Charles Marsh aufgestellt,[21] enthielt jedoch Ceratosaurus als einzigen Vertreter, weshalb sie von späteren Autoren als redundant betrachtet und nicht verwendet wurde.[38]
Ein aus dem Mitteljura Englands stammender Schädel weist ein ähnliches Nasenhorn auf wie das des Ceratosaurus. Friedrich von Huene (1926)[40] beschrieb diesen Schädel als Proceratosaurus („Vor-Ceratosaurus“), da er annahm, es handle sich um einen Vorläufer des spätjurassischen Ceratosaurus. Heute gilt Proceratosaurus als ein Vertreter der Tetanurae; Oliver Rauhut und Kollegen (2010) klassifizieren diese Gattung als den ältesten bekannten Vertreter der Tyrannosauroidea.[41] Das charakteristische Nasenhorn hat sich bei Proceratosaurus und Ceratosaurus somit konvergent (unabhängig voneinander) entwickelt.[30]
Paläobiologie und Paläoökologie
Lebensraum und ökologische Nischen

Alle in Nordamerika gefundenen Fossilien von Ceratosaurus stammen aus dem Kimmeridgium und Tithonium der Morrison-Formation, einer der wichtigsten Fundschichten für Dinosaurierfossilien. Ceratosaurus teilte sich den Lebensraum mit anderen Theropoden, wie dem Megalosauriden Torvosaurus und dem Allosauriden Allosaurus. So enthielt die Fundstätte Garden Park in Colorado, neben den Überresten von Ceratosaurus, die Überreste von Allosaurus. Der Cleveland-Lloyd-Quarry in Utah, der Dry Mesa Quarry in Colorado und das Dinosaur National Monument an der Grenze von Colorado und Utah wiesen jeweils die Überreste von mindestens drei Großtheropoden auf: Ceratosaurus, Allosaurus und Torvosaurus.[42][43] Auch Como Bluff und benachbarte Fundorte in Wyoming bargen die Reste von Ceratosaurus, Allosaurus und einem großen Megalosauriden.[44]
Verschiedene Studien versuchen zu klären, welche Mechanismen die direkte Konkurrenz dieser Spezies eingeschränkt haben könnten. Henderson (1998) untersuchte Ceratosaurus sowie zwei Morphotypen von Allosaurus – eine Form mit einer verkürzten Schnauze, einem hohen und breiten Schädel und kurzen, nach hinten gerichteten Zähnen sowie eine Form mit einer längeren Schnauze, einem niedrigeren Schädel und langen, senkrechteren Zähnen. Die Konkurrenz unter sympatrisch lebenden Spezies ist im Allgemeinen umso größer, je ähnlicher sich die Arten in ihrer Morphologie, Physiologie und ihrem Verhalten sind. Henderson kommt zu dem Ergebnis, dass die kurzschnäuzige Allosaurus-Morphe eine andere ökologische Nische besetzte als Ceratosaurus und die langschnäuzige Allosaurus-Morphe: So führte der kürzere Schädel dieser Allosaurus-Morphe zu einer Verringerung der Biegemomente, die der Schädel bei einem Biss ausgesetzt ist, was zu einem insgesamt stärkeren Biss führte, ähnlich wie bei einer Katze. Der langschnäuzige Schädel von Ceratosaurus und der anderen Allosaurus-Morphe könnte hingegen mit dem eines Hundes verglichen werden: So hatten die längeren Zähne möglicherweise die Funktion von Reißzähnen zum Zufügen von Schlitzwunden, wobei die Bisskraft wegen des schmaleren Schädels auf einen kleineren Bereich fokussiert war.[42]
Hendersons Vergleich der langschnäuzigen Allosaurus-Morphe mit Ceratosaurus ergab hingegen große Ähnlichkeiten, woraus er folgert, dass diese beiden Formen in direkter Konkurrenz miteinander standen. Henderson vermutet, dass Allosaurus Ceratosaurus immer weiter zurückgedrängt hat. So zeigt der Cleveland-Lloyd-Quarry die Überreste der langschnäuzigen Allosaurus-Morphe sowie die von Ceratosaurus, wobei Ceratosaurus sehr selten gewesen schien. Im Dry Mesa Quarry jedoch, welcher Fossilien von Ceratosaurus und der kurzschnäuzigen Allosaurus-Morphe barg, wird Ceratosaurus weitaus häufiger entdeckt.[42]
Die Evolution der extrem verlängerten Zähne bei Ceratosaurus interpretiert Henderson als direkte Folge des Konkurrenzdrucks mit der langschnäuzigen Allosaurus-Morphe. Um in eine andere ökologische Nische auszuweichen, könnte Ceratosaurus andere Beute bevorzugt haben; als Aasfresser könnte Ceratosaurus andere Teile von Kadavern verzehrt haben. Auch könnten die verlängerten Zähne als visuelles Signal zur Erkennung von Artgenossen gedient oder andere soziale Funktionen besessen haben. Die Evolution einer großen Körpergröße dieser Theropoden (6–8 m) trägt laut Henderson ebenfalls zur Verringerung von Konkurrenz bei, da mit zunehmender Körpergröße das Beutespektrum erhöht wird.[42]
In einer jüngeren Studie vermuten Robert Bakker und Gary Bir (2004), dass Ceratosaurus vor allem auf in Wasser lebende Beute wie Lungenfische, Krokodile und Schildkröten spezialisiert war. Eine statistische Analyse der Verteilung von Zahnfunden in 50 Fundstellen in und um Como Bluff ergab, dass Zähne von Ceratosaurus und Megalosauriden vor allem in aquatisch dominierten Fundstellen gefunden werden, wobei Ceratosaurus gelegentlich auch in Fundstellen mit Hinweisen auf trockene Bedingungen auftaucht. Allosauriden wurden dagegen in Fundstellen mit Hinweisen auf trockene Bedingungen und in aquatisch dominierten Fundstellen gleichermaßen gefunden.[45]
Diese Forscher folgern, dass Ceratosaurus und die Megalosauriden vor allem am und im Wasser jagten, wobei Ceratosaurus gelegentlich an Kadavern großer Dinosaurier fraß. Weiter weisen die Forscher auf den ungewöhnlich tiefen und damit krokodilähnlichen Schwanz von Ceratosaurus hin, bei dem es sich um eine Anpassung an das Schwimmen gehandelt haben könnte. Außerdem war der Körper von Ceratosaurus und Megalosauriden relativ lang, niedrig und flexibel, während Allosauriden einen kurzen, hohen und steifen Körper zeigten – somit wäre Ceratosaurus ein besserer Schwimmer gewesen als ein Allosauride. Allosauriden dagegen seien auf schnelles Laufen im offenen Gelände und auf das Erbeuten großer pflanzenfressender Dinosaurier wie Sauropoden und Stegosaurier ausgelegt, seien aber saisonbedingt auf aquatische Beutetiere ausgewichen, wenn diese Pflanzenfresser nicht vorhanden waren.[45]
Funktion der Kopfhörner
Othniel Charles Marsh (1884) und Charles Gilmore (1920) vermuten, dass das auf der knöchernen Basis aufsitzende Nasenhorn groß und scharfkantig war und sowohl zur Verteidigung als auch zum Angriff aktiv eingesetzt wurde.[6][7] Rowe und Gauthier (1990) halten dagegen, das Nasenhorn und die Brauenhörner seien nicht stabil genug, um im Kampf eingesetzt werden zu können. Stattdessen haben diese Hörner wahrscheinlich ausschließlich der Zurschaustellung gedient.[10] Auch die Reihe von Osteodermen auf der Mittellinie des Körpers könnte laut diesen Forschern der Zurschaustellung gedient haben.[10]
Paläopathologie
Der Fuß des Holotyp-Exemplars zeichnet sich durch die miteinander verschmolzenen Mittelfußknochen 2 und 4 aus. Obwohl einige Autoren annahmen, dass dies ein charakteristisches Merkmal der Gattung sei, handelte es sich um ein pathologisches Merkmal, möglicherweise um eine verheilte Fraktur.[46][47]
In der Populärkultur
Ceratosaurus mit seinem charakteristischen Hörnern gehört zu den bekannteren Dinosauriern und erscheint in fiktionaler Literatur,[2] beispielsweise ist er der Protagonist in Dino Buzzatis 1942 erschienener phantastischer Erzählung L’uccisione del drago (in deutscher Übersetzung erschienen als Die Ermordung des Drachen). Ceratosaurus tritt auch in vielen „Dinosaurierfilmen“ auf, etwa als Hauptfigur im Kampf gegen „Höhlenmenschen“ in Brute Force (1914) von David Wark Griffith, dem vielleicht ersten Dinosaurier-Realfilm der Filmgeschichte.[48] Im Fantasyfilm Eine Million Jahre vor unserer Zeit (1966) gibt es einen Kampf zwischen einem Ceratosaurus und einem Triceratops. Auch in dem Science-Fiction-Film Jurassic Park III sowie in den Spielfilmen T-Rex und Age of Dinosaurs – Terror in L.A. haben Ceratosaurier einen Auftritt.
Belege
Literatur
- Robert T. Bakker, Gary Bir: Dinosaur Crime Scene Investigations: Theropod Behavior at Como Bluff, Wyoming, and the Evolution of Birdness. In: Philip J. Currie, Eva B. Koppelhus, Martin A. Shugar, Joanna L. Wright (Hrsg.): Feathered Dragons. Indiana University Press, 2004, ISBN 0-253-34373-9, S. 301–342.
- Paul D. Brinkman: The Second Jurassic Dinosaur Rush. Museums and Paleontology in America at the Turn of the Twentieth Century. University of Chicago Press, 2010.
- B. B. Britt, C. A. Miles, K. C. Cloward, J. H. Madsen: A juvenile Ceratosaurus (Theropoda, Dinosauria) from Bone Cabin Quarry West (Upper Jurassic, Morrison Formation), Wyoming. 19 (Ergänzung zu 3), 33A. Journal of Vertebrate Paleontology, 1999.
- Kenneth Carpenter: The Cañon City Dinosaur Sites of Marsh and Cope. In: Society of Vertebrate Paleontology Field Trip. 1999.
- Matthew T. Carrano, Scott D. Sampson: The Phylogeny of Ceratosauria (Dinosauria: Theropoda). In: Journal of Systematic Palaeontology. Band 6, 2008, S. 183–236.
- John R. Foster: Paleoecological analysis of the vertebrate fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain region, USA. In: New Mexico Museum of Natural History and Science. Band 23, 2003, S. 1–95.
- C. W. Gilmore: Osteology of the carnivorous Dinosauria in the United States National Museum, with special reference to the genera Antrodemus (Allosaurus) and Ceratosaurus. Nr. 110. Bulletin of the United States National Museum, 1920, S. 1–154.
- Donald F. Glut: Dinosaurs, the encyclopedia. McFarland & Company, Inc. Publishers, 1997, ISBN 978-0-375-82419-7.
- Donald F. Glut, Michael K. Brett-Surman: Dinosaurs and the media. In: The Complete Dinosaur. Bloomington und Indianapolis: Indiana University Press, 1997, ISBN 0-253-33349-0, S. 675–706.
- Donald M. Henderson: Skull and tooth morphology as indicators of niche partitioning in sympatric Morrison Formation theropods. In: Gaia. Nr. 15, 1998, ISSN 0871-5424, S. 219–226.
- Werner Janensch: Die Coelurosaurier und Theropoden der Tendaguru-Schichten Deutsch-Ostafrikas. In: Palaeontographica, Supplement VIII: 1–100. 1925.
- F. v. Huene: On several known and unknown reptiles of the order Saurischia from England and France. In: Annals and Magazine of Natural History, Serie 9. Nr. 17, 1926, S. 473–489.
- James H. Madsen, Samuel Paul Welles: Ceratosaurus (Dinosauria, Theropoda): A Revised Osteology. Utah Geological Survey, 2000, ISBN 1-55791-380-3, S. 1–80.
- Othniel Charles Marsh: Principal characters of American Jurassic dinosaurs, part VIII: The order Theropoda. Band 27, Nr. 160. American Journal of Science, 1884, S. 329–340.
- O. Mateus, M. T. Antunes: Ceratosaurus (Dinosauria: Theropoda) in the Late Jurassic of Portugal. In: 31st International Geological Congress, Abstract Volume. Rio de Janeiro 2000.
- O. Mateus, A. Walen, M. T. Antunes: The large theropod fauna of the Lourinhã Formation (Portugal) and its similarity to the Morrison Formation, with a description of a new species of Allosaurus. In: J. R. Foster, S. G. Lucas (Hrsg.): Paleontology and Geology of the Upper Jurassic Morrison Formation. Bulletin 36. New Mexico Museum of Natural History and Science, 2006.
- R. E. Molnar: Theropod Paleopathology: A Literature Survey. In: D. H. Tanke, K. Carpenter (Hrsg.): Mesozoic Vertebrate Life. Indiana University Press, 2001, S. 337–363.
- Gregory S. Paul: Predatory Dinosaurs of the World. Simon & Schuster, 1988, ISBN 0-671-61946-2, Ceratosaurs.
- Oliver Rauhut: The interrelationships and evolution of basal theropod dinosaurs. In: Special Papers in Paleontology. Band 69, 2003.
- Oliver Rauhut, Angela Milner, Scott Moore-Fay: Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England. In: Zoological Journal of the Linnean Society. Band 158, Nr. 1, 2010, S. 155–195, doi:10.1111/j.1096-3642.2009.00591.x.
- Oliver Rauhut: Provenance and anatomy of Genyodectes serus, a large-toothed ceratosaur (Dinosauria: Theropoda) from Patagonia. In: Journal of Vertebrate Paleontology. Band 24, Nr. 4, 2004, S. 894–902.
- Timothy B. Rowe, Jacques Gauthier, David B Weishampel, P. Dodson, H. Osmolska: Ceratosauria. In: The Dinosauria. 1. Auflage. University of California Press, Berkeley 1990, ISBN 0-520-06726-6, S. 151–168.
- Matías Soto, Daniel Perea: A Ceratosaurid (Dinosauria, Theropoda) from the Late Jurassic–Early Cretaceous of Uruguay. In: Journal of Vertebrate Paleontology. Band 28, Nr. 2, 2008, S. 439–444.
- Darren H. Tanke, Bruce M. Rothschild: Dinosores: An Annotated Bibliography of Dinosaur Paleopathology and Related Topics – 1838–2001. In: New Mexico Museum of Natural History and Science. Bulletin 20, 2001.
- François Therrien, Donald M. Henderson: My theropod is bigger than yours … or not: estimating body size from skull length in theropods. In: Journal of Vertebrate Paleontology. Band 27, Nr. 1, 2007, S. 108–115.
- R. S. Tykoski, T. Rowe: Ceratosauria. In: D. B. Weishampel, P. Dodson, H. Osmólska (Hrsg.): The Dinosauria. 2. Auflage. University of California Press, Berkeley 2004, ISBN 0-520-24209-2, S. 47–70.
- Xing Xu, James M. Clark, Jinyou Mo, Jonah Choiniere, Catherine A. Forster, Gregory M. Erickson, David W. E. Hone, Corwin Sullivan, David A. Eberth, Sterling Nesbitt, Qi Zhao, Rene Hernandez, Cheng-kai Jia, Feng-lu Han, Yu Guo: A Jurassic ceratosaur from China helps clarify avian digital homologies. In: Nature. Band 459, 18. Mai 2009, S. 940–944, doi:10.1038/nature08124.
- Wolfgang Zils, Christa Werner, Andrea Moritz, Charles Saanane: Tendaguru, the most famous dinosaur locality of Africa. Review, survey and future prospects. In: documenta naturae. Band 97. München 1995, S. 1–41.
Weblinks
- Ben Creisler: Dinosauria Translation and Pronunciation Guide (Memento vom 13. Oktober 2011 im Internet Archive)
Einzelnachweise
- Gregory S. Paul: The Princeton Field Guide To Dinosaurs, 2010. ISBN 978-0-691-13720-9, S. 84–85 Online (Memento des Originals vom 13. Juli 2015 im Internet Archive) Info: Der Archivlink wurde automatisch eingesetzt und noch nicht geprüft. Bitte prüfe Original- und Archivlink gemäß Anleitung und entferne dann diesen Hinweis.
- Glut 1997, S. 266–270
- Gilmore 1920 S. 114–115
- Madsen und Welles 2000
- Therrien und Henderson 2007, S. 109
- Marsh 1884, S. 330–331
- Gilmore 1920, S. 82
- Britt et al. 1999
- Tykoski und Rowe 2004, S. 66
- Rowe et al. 1990, S. 167
- Tykoski und Rowe 2004, S. 52
- Madsen und Welles 2000, S. 23
- Rauhut 2004, S. 900
- Tykoski und Rowe 2004, S. 54
- Tykoski und Rowe 2004, S. 58
- Gilmore 1920, S. 113–114
- Tykoski und Rowe 2004, S. 59
- Gilmore 1920, S. 104
- Brinkman 2010, S. 10
- Carpenter 1999, S. 8
- Marsh 1884
- Ben Creisler. Abgerufen am 29. Januar 2011
- Gilmore 1920
- Paul 1988, S. 274–279
- Madsen und Welles 2000, S. 21
- Madsen und Welles 2000, S. 2–3
- Madsen und Welles 2000, S. 35–37
- Zils et al. 1995, S. 1
- Janensch 1925, S. 61–65
- Carrano und Sampson, S. 192
- Rowe et al. 1990, S. 165
- Janensch 1925, S. 90
- Mateus et al. 2006, S. 1–2
- Mateus 2000
- Soto und Perea, 2008
- Rauhut 2003, S. 25
- Xu et al. 2009, Supplementary Information, S. 27
- Carrano und Sampson, S. 185 f.
- Rauhut 2004, S. 895
- Von Huene 1926
- Rauhut 2010
- Henderson, 1998
- Foster 2003, S. 20
- Bakker und Bir 2004, S. 106–107
- Bakker und Bir 2004, S. 123–139
- Tanke et al. 2001, S. 9, 25, 42–43
- Molnar 2001, S. 339–340
- Glut und Brett-Surman 1997, S. 676