Cro-Magnon-Mensch
Cro-Magnon-Mensch (ˌkroːmaˈɲɔ̃) ist eine – in der europäischen Forschungstradition begründete – Bezeichnung für den anatomisch modernen Menschen (Homo sapiens) des westlichen Eurasiens, der während der letzten Kaltzeit lebte. Als Epoche der Cro-Magnon-Menschen gilt die Zeitspanne vom ersten Nachweis von Homo sapiens in Europa vor annähernd 45.000 Jahren bis zum Übergang vom Pleistozän zum Holozän vor etwa 12.000 Jahren.[1]
| Cro-Magnon-Mensch | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
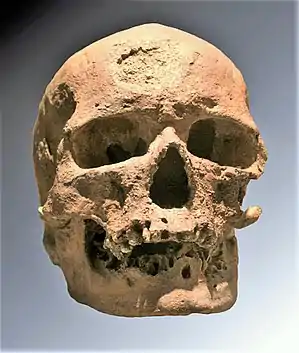
Schädel Cro-Magnon I („alter Mann von Cro-Magnon“) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Homo sapiens | ||||||||||||
Die früher gehegte Vermutung, der Cro-Magnon-Mensch sei ein evolutionäres Bindeglied zwischen Neandertaler und modernem Homo sapiens, gilt seit Mitte der 1970er-Jahre als widerlegt, und zwar u. a. aufgrund von anatomischen Merkmalen, die Neandertaler und modernen Homo sapiens trennscharf unterscheiden.[2] In jüngerer Zeit wurde dies anhand genetischer und archäologischer Merkmale bestätigt (vergl. Europäer). „Cro-Magnon-Mensch“ bezeichnet demnach weder eine Art noch eine Unterart.
Typusexemplar Cro-Magnon I
Der Holotyp Cro-Magnon I war der erste von fünf Schädeln und postcranialen Skelettresten, die 1868 durch Louis Lartet, den Sohn von Édouard Lartet, im Abri de Cro-Magnon (Dordogne) bei Ausgrabungen entdeckt wurden.[3][4] Jean Louis Armand de Quatrefages und Ernest Hamy beschrieben auf Basis dieser Funde erstmals 1874 eine „Cro-Magnon-Rasse“.[5] 1877 erfolgte eine vergleichende Darstellung im Buch L’Espèce humaine (Die menschliche Spezies).[6]
Versuche im späten 20. Jahrhundert, die Menschenreste mittels 14C-Methode direkt zu datieren, scheiterten entweder am zu geringen Kollagengehalt oder an der Tatsache, dass die postcranialen Skelettteile in den Sammlungsbeständen des Musée de l’Homme nicht mehr eindeutig den Schädeln zuzuordnen sind. So wurden die Gräber bislang nur indirekt durch die Beigaben datiert: Eine durchbohrte Meeresschnecke (Littorina littorea), die als Teil einer Halskette einem der Gräber von Cro-Magnon zuzuordnen ist, wurde mit der AMS-Methode auf 27.680 ± 270 BP (Beta 157439) datiert.[7][8] Damit stammen auch die Menschenreste wahrscheinlich aus dem Gravettien, sofern – zum derzeitigen Publikationsstand – keine Störungen des Befundzusammenhangs in Betracht gezogen werden. Da das Gravettien als Kulturstufe erst um die Mitte des 20. Jahrhunderts etabliert wurde, entspricht es der alten Klassifikation des Oberen Aurignacien und nicht – wie ursprünglich für die Menschenreste von Cro-Magnon angenommen wurde – dem älteren Aurignacien typique.
1899 wurde das Fossil Cro-Magnon I von Georges Vacher de Lapouge als Typusexemplar für die von ihm vorgeschlagene Art Homo spelaeus („Höhlenmensch“) benannt.[9]
Fossilbericht
-Cro-Magnon.JPG.webp)
Die drei ältesten gefundenen Fossilien des Cro-Magnon-Menschen (im Sinne des europäischen Homo sapiens) stammen aus der Batscho-Kiro-Höhle in Bulgarien, 46.790–42.810 Jahre cal BP,[10], der Grotta del Cavallo (Apulien, 45.000–43.000 cal BP)[11] und aus Kents Cavern (England, 44.200–41.500 cal BP).[12]
Etwas jünger sind die Knochenfunde aus einer Höhle in Rumänien (Peștera cu Oase):[13] Für den Unterkiefer Oase 1 wurde 2003 ein Alter von rund 40.500 Kalenderjahren (Radiokohlenstoffdatierung, kalibriert) berechnet,[14][15] für den Schädel Oase 2 rund 35.000 BP[16] (was kalibriert ebenfalls ein Alter von rund 40.000 Jahren ergibt); 2020 wurde für Oase 1 eine Neuberechnung publiziert, die für das Fossil ein Alter von 41.770 bis 37.310 Jahren in kalibrierten Kalenderjahren (cal BP) ergab.[17]
Weitere Menschenreste, die älter als 30.000 BP datieren, stammen von Mladeč in Mähren (zirka 31.000 BP)[18][19] und Kostenki am Don (39.000–37.000 cal BP).[20][21] Zwei morphologische Varianten der Menschen des Jungpaläolithikums – eine hochwüchsige und eine grazile – lassen sich deutlich unterscheiden. Die Menschen der Grotte des Enfants 4, Barma Grande 5, Předmosti 3, Pavlov und Sungir 1 gehörten dem sehr hochwüchsigen Typus an, diejenigen von Arene Candide 2, 3, 5 und Riparo Continenza dem ausgesprochen grazilen.[22][23] Eine Mittelstellung nehmen der Cro-Magnon 2, Předmosti 9, Předmosti 14, Paviland, Ohalo 2, und Wadi Kubbaniya ein und, etwas kleiner, die Menschen von Předmosti 5, Arene Candide 12, Riparo Continenza und das weit jüngere Skelett aus der bayerischen Klausenhöhle (zirka 18.000 BP). Von einigen Bearbeitern wird daher der Cro-Magnon-Typ dem Brünn-Typ (früher: Brünn-Rasse) gegenübergestellt.[24][25]
Am Skelett Paglicci 23 aus der Paglicci-Höhle (Italien) wurde die Cambridge reference sequence der mtDNA eines Cro-Magnon-Menschen erstellt.[26] Eine weitere mtDNA-Sequenz liegt vom Grab aus Kostenki 14 vor, das der Haplogruppe U2 angehört.[27] Im Jahre 2013 wurde das Genom des etwa 24.000 Jahre alten Jungen aus Malta in Sibirien publiziert.[28]
Lange Zeit galt die Zuordnung des 1909 entdeckten Grabes von Combe Capelle[29] zum Châtelperronien als gesichert, da Artefakte in der Umgebung des Skeletts als Grabbeigaben interpretiert wurden.[30] Im Jahre 2011 wurden neue AMS-Daten bekannt, die die Bestattung wesentlich jünger ins Mesolithikum datieren.[31]
Werkzeugfunde aus Willendorf, Österreich, die den Cro-Magnon-Menschen zugeschrieben werden, wurden auf ein Alter von 43.500 Jahren cal BP datiert.[32]
Laut einer Berechnung von Isabell Schmidt und Andreas Zimmermann (Universität zu Köln) aus dem Jahr 2019 lebten in der Zeitspanne von etwa 42.000 bis etwa 33.000 Jahren vor heute (cal BP) maximal rund 3300 Individuen in West- und Mitteleuropa.[33] „Nur fünf Gebiete in Europa hatten nach diesen Schätzungen überhaupt eine überlebensfähige Population von etwa 150 Personen oder mehr: Nordspanien, Südwestfrankreich, Belgien, Teile Tschechiens und der obere Donauraum. Dass die Zentren dieser lebensfähigen Populationen etwa 400 Kilometer voneinander entfernt waren, ist ein europaweit einheitliches Muster.“[34] Daneben seien noch weitere Gebiete Europas zumindest zyklisch – während bestimmter Jahreszeiten – besiedelt gewesen. Es habe sich um „hochmobile Jäger-Sammler Gruppen“ gehandelt, „die regelmäßig Distanzen von 200 km zurücklegten und zudem an verschiedene Habitate angepasst waren.“
Die 2022 erfolgte Zuschreibung eines in der Grotte Mandrin (Département Drôme, Südfrankreich) einzeln gefundenen, stark beschädigten Zahns, der 56.800 bis 51.700 Jahre (cal BP) ist, zu Homo sapiens, ist umstritten.[35]
Beschreibung
Homo sapiens ist als Taxon vor zirka 200.000 bis 150.000 Jahren in Afrika entstanden (siehe archaischer Homo sapiens). Die Verwendung des Begriffs Cro-Magnon-Mensch im Sinne einer Chronospezies ist weder mit anatomischen Merkmalen noch aufgrund der Untersuchung alter DNA haltbar. Trotz eines sogenannten genetischen Flaschenhalses der Homo-sapiens-Populationen während der letzten Eiszeit konnte in einer 2011 publizierten Studie am menschlichen Genom nachgewiesen werden, dass ein genetischer Austausch der Populationen in Europa, Asien und Afrika bis vor etwa 40.000 bis 20.000 Jahren stattfand.[36]
Cro-Magnon-Menschen waren Jäger und Sammler und lebten überwiegend nomadisch. Viele Fundstellen zeigen lediglich kurzfristige oder über einen längeren Zeitraum jeweils saisonal genutzte Lagerplätze des Menschen an. Länger als ein Jahr wahrscheinlich permanent besiedelte Lagerplätze sind frühestens seit dem Gravettien bekannt, zum Beispiel in Dolní Věstonice und Pavlov (beide Mähren).
Eine Anfang 2013 publizierte Studie zur Genetik der Pigmentation der Haut ergab Anhaltspunkte dafür, dass die Erstbesiedler des Homo sapiens in Europa vermutlich dunklere Haut als heutige Mitteleuropäer hatten. Laut den Forschern war die Hautfarbe der frühen Europäer noch etwas dunkler als bei heutigen Nordafrikanern (etwa den Berbern).[37][38] 2017 erbrachte eine verfeinerte Genanalyse Hinweise darauf, dass auch die für helle Hautfarbe codierenden Gene ihren Ursprung bereits in Afrika hatten, da die relativ hellhäutigen San in Südafrika die gleichen Varianten der Gene HERC2 und OCA2 besitzen, die mit der hellen Haut- und Augenfarbe der Europäer in Verbindung gebracht werden.[39] In einem Begleitartikel in der Fachzeitschrift Science wurde erwogen, dass die aus Afrika ausgewanderten Menschen sowohl Genvarianten für helle als auch für dunkle Hauttönung besaßen und dass die dunklen Varianten später in Eurasien verloren gingen,[40] sodass auch aus diesem Grund die Einteilung des Homo sapiens anhand der Hautpigmentierung in Rassen ähnlich unsinnig sei wie es die Einteilung anhand der Körpergröße wäre.
Ablösung der Neandertaler

Die vollständige Ablösung der Neandertaler (engl. replacement) durch anatomisch moderne Menschen vor mehr als 35.000 Jahren ist Gegenstand zahlreicher Theorien. Im Jahre 2012 wurde eine Studie der aDNA von Neandertalern veröffentlicht, die nahelegt, dass bereits vor dem Auftreten moderner Menschen in Europa eine starke Ausdünnung der Neandertaler-Population erfolgt war, was auf Umwelt- und andere Stressfaktoren zurückgeführt wird.[41] Die geringe Nachweisbarkeit von Neandertalern jünger als 40.000 BP wird durch die Neubewertung von 14C-Datierungen gestützt. Daten ohne Probenaufbereitung mit Ultrafiltration seien nach Ansicht von Forschern der CalPal-Arbeitsgruppe nur als Minimalalter anzusehen.[42][43] Demnach markieren die Fossilien aus der kaukasischen Mesmaiskaja-Höhle (39.700 ± 1.100 BP) sowie aus der kroatischen Vindija-Höhle die jüngsten Neandertalerfunde mit gesicherter 14C-Datierung.
Neandertaler-Genom beim Homo sapiens
.jpg.webp)
Obwohl im Genom anatomisch moderner Menschen Eurasiens ein Anteil von bis zu 4 % Neandertaler-spezifischer Gene festgestellt wurde,[45] gilt dies für taxonomische Fragen als unerheblich.
- Allerdings erbrachten in den Jahren 2013 bis 2015 durchgeführte genanalytische Untersuchungen an den Homo-sapiens-Funden von Peștera cu Oase in Rumänien und Ust-Ischim in Sibirien weitere Erkenntnisse zum Thema Hybridisierung: In beiden Fossilien wurde Neandertaler-DNA nachgewiesen.[46]
- Im Falle des Unterkiefers von Oase1 wurde ein Anteil von 5 bis 11 Prozent an DNA-Abschnitten des Neandertalers nachgewiesen und abgeschätzt, dass die Hybridisierung vier bis sechs Generationen vor Datierung der Fundlage (40.000 v.h.) stattgefunden hat – auch wenn es sich anscheinend um eine Linie ohne heute nachweisbare Nachkommen handelt.[47] Beim Fund von Ust-Ischim wurde ein Anteil von 2 Prozent an Neandertaler-DNA festgestellt. Der Zeitpunkt des Genflusses wurde auf rund 7.000 bis 13.000 Jahre vor Lebzeiten des Individuums (vor ca. 45.000 Jahren) datiert – mit genetischer Nähe zu den in Eurasien lebenden Menschen.[48]
- Auch wenn die Studien nichts darüber aussagen, ob der Genfluss im Umfeld der Fundstellen erfolgt ist oder die Hybriden aus entfernteren Gebieten zugewandert sind, belegen sie doch, dass Verpaarungen zwischen Neandertaler und Cro-Magnon-Mensch (bzw. frühem Eurasischen Homo sapiens) nicht nur in der Levante, sondern auch im Osten Europas und in Sibirien tatsächlich stattgefunden haben.[49]
Der europäische Cro-Magnon-Mensch tritt in seiner materiellen Kultur als Träger des Jungpaläolithikums mit der Klingenkultur des Aurignaciens in Erscheinung. Diese ist mit dem Ahmarien im Vorderen Orient bereits einige Tausend Jahre früher als in Mitteleuropa nachgewiesen. Wahrscheinlich drangen moderne Menschen erstmals vor etwa 36.000 Jahren BP (kalibriert 40.000 bis 42.000 v. Chr.) nach Europa vor. Einen starken Bevölkerungsrückgang gab es während des Kältemaximums der Weichseleiszeit (bzw. Würmeiszeit im alpinen Raum) vor etwa 22.000 bis 18.000 Jahren.[50]
Die Vermutung, dass die Cro-Magnon-Menschen, von Südosten kommend, die „kältegewohnten“ Neandertaler in der Zeit vor diesem Temperaturminimum in nördliche Refugien abgedrängt hätten, scheint jedoch irrig, da Neandertaler kurz vor ihrem Aussterben nur noch in Südeuropa nachgewiesen sind, während Siedlungsplätze der Cro-Magnon-Menschen vielfach nördlich der Alpen belegt sind. In Westeuropa befand sich ein früher Siedlungsraum von Cro-Magnon-Menschen in Südfrankreich und Nordspanien, während die Neandertaler den Süden der Iberischen Halbinsel bis nach Gibraltar besiedelten. In Osteuropa ergibt sich ein ähnliches Bild mit relativ nördlich gelegenen Fundorten moderner Menschen bei Kostenki am Don aus dem Early Upper Palaeolithic (EUP). Zu dieser Zeit lagen die bislang erwiesenen Siedlungsgebiete der Cro-Magnon-Menschen im Grenzgebiet zwischen Tundren-, Kaltsteppen- und Nadelbaumvegetation, während die Neandertaler im wärmeren Grenzgebiet zwischen Nadel- und Laubbaumvegetation siedelten.
Ernährung
Untersuchungen stabiler Isotope (13C- und 15N-Gehalt) im Kollagen der Knochen von Cro-Magnon-Menschen konnten zeigen, dass sich das Nahrungsangebot im Vergleich zu den Neandertalern vervielfältigt hat. Während Neandertaler überwiegend das Fleisch großer Pflanzenfresser verzehrten,[51] ist bei Cro-Magnon-Menschen des mittleren Jungpaläolithikums (Gravettien) ein erhöhter Anteil von Fischen und Muscheln gemessen worden.[52][53][54] Einen prominenten Fall stellt die Analyse am „Prinzen“ aus der Höhle Arene Candide (bei Finale Ligure) dar. Bei diesem Jugendlichen (Datierung 23.440 + 190 BP) konnte Ernährung mit Fischarten aus dem Mittelmeer nachgewiesen werden, die heute nur noch im Atlantik vorkommen.[55] Der Anteil von Fisch an der Gesamtdiät wird mit 20 bis 25 % beziffert. Ernährung mit einem hohen Anteil von Großwild und Fisch (Lachse) wurde auch bei der Magdalénien-Bestattung einer Frau von Saint-Germain-de-la-Rivière festgestellt.[56]
Bei den britischen Fundplätzen Gough’s Cave und Sun Hole Cave aus dem spätpaläolithischen Creswellien lag der Anteil mariner Ressourcen (Fisch, Muscheln) bei etwa 30 %.[57] Dasselbe gilt für den spanischen Fundplatz Balma Guilanyà im südöstlichen Vorland der Pyrenäen.[58]
Archäologische Kulturen

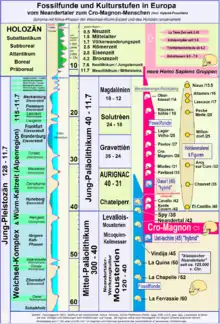
Mit dem Auftreten des „modernen Menschen“ beginnt zugleich die Periode des Jungpaläolithikums. Während die Koexistenz mit dem Neandertaler meist auf zirka 10.000 Jahre zwischen 40.000 und 30.000 Jahre beziffert wird,[59] deuten 14C-Daten unter Einbeziehung von Kalibrationsmodellen nur eine recht kurze Koexistenz von Werkzeugkulturen später Neandertaler und des frühen Jungpaläolithikums in einem Zeitfenster von zwei- bis dreitausend Jahren an.[60][61]
Die Werkzeuge und Waffen der Cro-Magnon-Menschen des Aurignacien waren denjenigen der letzten Neandertaler (Szeletien, Châtelperronien bis etwa 35.000 BP) nicht signifikant überlegen, weisen jedoch spezifisch neue Merkmale auf. Eine wechselseitige Beeinflussung der materiellen Kultur (Steinwerkzeuge, Schmuck) wird diskutiert, kann jedoch nur im Fall der Typuslokalität Châtelperron (Département de l'Allier) durch Interstratifikation der Schichten belegt werden.[62][63]
Mit dem Gravettien ist eine erhebliche technische Verfeinerung der Artefakte des Cro-Magnon-Menschen zu verzeichnen, wie das Einkleben von Rückenmessern in Speere, die spätestens seit dem Solutréen mit Speerschleudern abgeworfen wurden. Die Speerschleuder war im Magdalénien Südwesteuropas massenhaft verbreitet und zu dieser Zeit die wichtigste Jagdwaffe. Am Ende des Jungpaläolithikums ist mit zunehmender Bewaldung erstmals der Bogen als Waffe nachgewiesen.
Kunst
Während beim Neandertaler dekorativ-künstlerische Hinterlassenschaften bis dato als Ausnahme in Erscheinung getreten sind (Steinkreise in der Höhle von Bruniquel), entwickelte der Cro-Magnon-Mensch erste Werke der bildenden Kunst.[64] Das betrifft einerseits die Felsbilder (Höhlenmalerei, Felsmalerei und Petroglyphen), andererseits die jungpaläolithische Kleinkunst (zum Beispiel so genannte Venusfigurinen).
Die ältesten Höhlenmalereien von Cro-Magnon-Menschen sind aus der Zeit des Aurignacien und evtl. aus der zuvor liegenden Epoche des Moustérien bekannt (El-Castillo-Höhle, Chauvet-Höhle); meistens handelt es sich dabei um Tierdarstellungen.[65] Zur selben Zeit treten Petroglyphen (Felsritzungen) auf (La Ferrassie, Höhle von Pair-non-Pair). Der quantitative Höhepunkt der Felsbildkunst wurde im Magdalénien erreicht (zum Beispiel Höhle von Lascaux, Höhle von Altamira).
Die jungpaläolithische Kleinkunst ist sehr vielfältig. Überliefert sind zum Beispiel Schnitzereien aus Mammutelfenbein, Geweih, Knochen und schnitzfähigem Gestein (Steatit, Kalkstein).[66] Kunstwerke aus anderen organischen Materialien (zum Beispiel Holz) waren wahrscheinlich ebenfalls in Gebrauch, sind archäologisch aber nicht nachweisbar. Auf Tätowierung und/oder Körperbemalung kann aus den oberflächigen Verzierungen einiger figürlicher Darstellungen geschlossen werden. Als mögliche Beispiele können Kerbmuster auf dem Oberarm des Löwenmenschen vom Hohlenstein-Stadel oder der Venus vom Hohlefels angeführt werden. Die Interpretation ist hier insofern unsicher, als auch gleichalte Tierfiguren aus der Vogelherdhöhle und dem Geißenklösterle solche Ornamente aufweisen. Sofern es sich um Totem-Darstellungen handelt, wäre die Übertragung menschlicher Merkmale jedoch plausibel.[67][68]
Neben der bildenden Kunst sind aus der Zeit des Aurignacien auch die ältesten Musikinstrumente bekannt. Dabei handelt es sich um Flöten aus Tierknochen.
Höhepunkt und Ende der Cro-Magnon-Phase
Während des letzten Kältemaximums (in Norddeutschland als Brandenburg- bzw. Frankfurt-Phase vor ca. 25.000 bis 20.000 Jahren bezeichnet), glich Europa bis in das Gebiet des heutigen Südfrankreichs einer baumlosen Tundra. Die Temperaturen lagen um 5 bis 13 °C niedriger als heute. Der Meeresspiegel lag um etwa 120 m tiefer als jetzt, Gletscher erstreckten sich mit ihren bis zu 3000 m dicken Eisschilden teilweise bis nach Mitteleuropa. Im Süden waren die Alpen bis ins Vorland vergletschert, auch die Pyrenäen. Der Raum zwischen dem nördlichen Eisschild und den Alpen war wie heute in Sibirien von Permafrostböden gekennzeichnet.[69][70]
In der Jüngeren Dryaszeit (ca. 12.000 vor heute) ging die mittlere Jahrestemperatur nochmals stark zurück. Zu dieser Zeit waren die Höhlenkunst und die Kleinkunst (Venusfiguren) des Cro-Magnon-Menschen bereits im Niedergang begriffen. Es herrschte ein großer Mangel an Nahrung und die Bevölkerungsdichte ging allmählich zurück. Am Ende dieser Phase waren weite Teile Mitteleuropas vom Homo sapiens verlassen.[71]
Erst mit dem Einzug der neolithischen Wirtschaftsweise in Mitteleuropa wurden die wildbeuterisch lebenden Cro-Magnon-Menschen, die im nacheiszeitlichen Mesolithikum genetisch noch fortbestanden, durch eingewanderte neue Populationen abgelöst, was durch Untersuchungen der mtDNA plausibel gemacht werden kann.[72] Demzufolge entstammen die spätglazialen und holozänen europäischen Wildbeuter des Mesolithikums überwiegend der Haplogruppe U, während bei frühneolithischen wie auch heutigen europäischen Menschen die Haplogruppe H dominiert.[73] Beim geographisch unscharf definierten Begriff des Cro-Magnon-Menschen als eiszeitlichem, europäischem Typen lässt sich daraus zwar ableiten, dass es in Mitteleuropa einen Bevölkerungswandel gab. Dieser bezieht sich jedoch nur auf Populationen des modernen Menschen, deren Haplogruppen aus Skelettmaterial der letzten 20.000 Jahre rekonstruiert wurden.
Literatur
- Friedemann Schrenk: Die Frühzeit des Menschen. Der Weg zum Homo sapiens (= Beck'sche Reihe. 2059). C. H. Beck, München 1997, ISBN 3-406-41059-6, S. 114.
- Henri-Victor Vallois: La Découverte des hommes de Cro-Magnon, son importance anthropologique. In: L'homme de Cro-Magnon. Anthropologie et archéologie. 1868–1968. Avant-propos de Gabriel Camps et Georges Olivier. Arts et métiers graphiques, Paris 1970, S. 11–20.
Weblinks
Einzelnachweise
- www.talkorigins.org
- Chris Stringer: Population relationships of later Pleistocene hominids: A multivariate study of available crania. In: Journal of Archaeological Science. Band 1, Nr. 4, 1974, S. 317–342, doi:10.1016/0305-4403(74)90051-X.
- Louis Lartet: Une sépultre des Troglodytes du Périgord (crânes des Eyzies). In: Bulletins de la Société d'Anthropologie de Paris. Band 3, 1868, S. 335–49.
- Louis Lartet: Mémoire sur une sépulture des anciens troglodytes de Périgord. In: Annales des sciences naturelles II Zoologie. 5ème Série, X, 1868.
- Jean Louis Armand de Quatrefages, Ernest Hamy: La race de Cro-Magnon dans l’espace et dans le temps. In: Bulletins de la Société d’Anthropologie de Paris. Band 9, 1874, S. 260–266.
- Jean Louis Armand de Quatrefages, Ernest Hamy: L’Espèce humaine. 1877.
- D. Henry-Gambier, R. W. White: New chrono-cultural data on the Cro-Magnon and Combe-Capelle human remains (Dordogne, France): consequences for the biocultural origins of modern humans in Europe. In: Annual Meetings of the Paleoanthropology Society. Tempe, Arizona 2003.
- D. Henry-Gambier: Les fossiles de Cro-Magnon (Les Eyzies-de-Tayac, Dordogne): Nouvelles donnees sur leur Position chronologique et leur attribution culturelle. In: Bull. et Mém. de la Société d’Anthropologie de Paris. Band 14, 2002, S. 89–112.
- Georges Vacher de Lapouge: L'Aryen, son rôle social. Cours libre de science politique, professé à l'Université de Montpellier (1889–1890). Ancienne Librairie Thorin et Fils, Paris 1899, S. 178, Digitalisat.
- Jean-Jacques Hublin, Nikolay Sirakov, Vera Aldeias et al.: Initial Upper Palaeolithic Homo sapiens from Bacho Kiro Cave, Bulgaria. In: Nature. Band 581, Nr. 7808, 2020, S. 1–4, doi:10.1038/s41586-020-2259-z.
- Stefano Benazzi et al.: Early dispersal of modern humans in Europe and implications for Neanderthal behaviour. In: Nature. Band 479, 2011, S. 525–528, doi:10.1038/nature10617.
- Tom Higham et al.: The earliest evidence for anatomically modern humans in northwestern Europe. In: Nature. Band 479, 2011, S. 521–524, doi:10.1038/nature10484.
- Erik Trinkaus: European early modern humans and the fate of the Neandertals. In: PNAS. Band 104, 2007, S. 7367–7372, doi:10.1073/pnas.0702214104.
- Erik Trinkaus et al.: Early modern human cranial remains from the Peștera cu Oase, Romania. In: Journal of Human Evolution. Band 45, 2003, S. 255–259, doi:10.1016/j.jhevol.2003.08.003.
- Erik Trinkaus et al.: An early modern human from Peștera cu Oase, Romania. In: PNAS. Band 100, Nr. 20, 2003, S. 11231–11236, doi:10.1073/pnas.2035108100.
- Hélène Rougier et al.: Peștera cu Oase 2 and the cranial morphology of early modern Europeans. In: PNAS. Band 104, Nr. 4, 2007, S. 1165–1170, doi:10.1073/pnas.0610538104.
- Johannes van der Plicht et al.: Recent developments in calibration for archaeological and environmental samples. In: Radiocarbon. Online-Veröffentlichung vom 21. April 2020, doi:10.1017/RDC.2020.22.
- E. M. Wild, M. Teschler-Nicola, W. Kutschera, P. Steier, E. Trinkaus, W. Wanek: Direct dating of Early Upper Palaeolithic human remains from Mladeč. In: Nature. Band 435, 2005, S. 332–335 doi:10.1038/nature03585.
- Zur Datierung von Mladeč.
- Rob Dinnis et al.: New data for the Early Upper Paleolithic of Kostenki (Russia). In: Journal of Human Evolution. Band 127, 2019, S. 21–40, doi:10.1016/j.jhevol.2018.11.012.
- A. A. Sinitsyn, P. Allsworth-Jones, R. A. Housley: Kostenki 14 (markina gora): New AMS dates and their significance within the context of the site as a whole. In: Prehistoire Europeene. 1996, S. 269–271.
A. A. Sinitsyn: Kostenki 14 (markina gora): Data, problems, and perspectives. In: Prehistoire Europeene. 1996, S. 273–313. - Paul B. Pettitt, M. Richards, R. Maggi and V. Formicola: The Gravettian burial known as the Prince („Il Principe“): new evidence for his age and diet. In: Antiquity. Band 77, Nr. 295, 2003, S. 15–19.
- K. Absolon, B. Klíma: Předmosti. Ein Mammutjägerplatz in Mähren. In: Fonte Archaeologiae Moraviae. VIII (Praha), 1977.
- E. Vlček: Relations morphologiquies des types humains fossiles de Brno et Cro-Magnon au Pleistocene Supérieur d'Europe. In: G. Camps, G. Olivier (Hrsg.): L’Homme de Cro-Magnon. Arts et Métiers Graphiques. Paris, 1970, S. 59–72.
- Martin Oliva: The Brno II Upper Palaeolithic burial. In: Wil Roebroeks, Margherita Mussi, J. Svoboda, K. Fennema (Hrsg.): Hunters of the Golden Age. Kolloquium Pavlov. Annalecta Praehistorica Leidensia 31. Leiden. 2000, S. 143–154.
- David Caramelli et al.: A 28,000 years old Cro-Magnon mtDNA sequence differs from all potentially contaminating modern sequences. In: PLOS ONE. Band 3, Nr. 7, e2700 (2008), doi:10.1371/journal.pone.0002700.
- Johannes Krause et al.: A complete mtDNA genome of an early modern human from Kostenki, Russia. In: Current Biology. Band 20, Nr. 3, 2010, S. 231–236, doi:10.1016/j.cub.2009.11.068.
- Maanasa Raghavan, Pontus Skoglund et al.: Upper Palaeolithic Siberian genome reveals dual ancestry of Native Americans. In: Nature. Band 505, 2014, S. 87–91, doi:10.1038/nature12736.
- H. Klaatsch, O. Hauser: Homo Aurignaciensis Hauseri – Ein paläolithischer Skelettfund aus dem unteren Aurignacien der Station Combe Capelle bei Montferrand (Périgord). In: Prähistorische Zeitschrift I. Band 3, Nr. 4, 1910, S. 273–338.
- Almut Hoffmann, Dietrich Wegner: Homo Aurignaciensis Hauseri – Ein paläolithischer Skelettfund aus dem unteren Aurignacien der Station Combe Capelle bei Montferrand/Périgord. In: Acta Praehistorica et Archaeologica. Band 35, 2003, S. 113–137 ISSN 0341-1184.
- Almut Hoffmann et al.: The Homo aurignaciensis hauseri from Combe-Capelle - A Mesolithic burial. In: Journal of Human Evolution. Band 61, Nr. 2, 2011, S. 211–214, doi:10.1016/j.jhevol.2011.03.001.
- Philip R. Nigst et al.: Early modern human settlement of Europe north of the Alps occurred 43,500 years ago in a cold steppe-type environment. In: PNAS. Band 111, Nr. 40, 2014, S. 14394–14399, doi:10.1073/pnas.1412201111.
- Isabell Schmidt und Andreas Zimmermann: Population dynamics and socio-spatial organization of the Aurignacian: Scalable quantitative demographic data for western and central Europe. In: PLoS ONE. Band 14, Nr. 2, 2019. e0211562, doi:10.1371/journal.pone.0211562.
- In Europa lebten im frühen Jungpaläolithikum im Schnitt nur 1.500 Menschen. Auf: idw-online.de vom 5. März 2019.
- Ludovic Slimak et al.: Modern human incursion into Neanderthal territories 54,000 years ago at Mandrin, France. In: Science Advances. Band 8, Nr. 6, 2022, doi:10.1126/sciadv.abj9496.
Did Neanderthals and modern humans take turns living in a French cave? Auf: science.org vom 9. Februar 2022.
Evidence of Europe’s first Homo sapiens found in French cave. Auf: nature.com vom 9. Februar 2022. - Heng Li, Richard Durbin: Inference of human population history from individual whole-genome sequences. In: Nature. Band 475, 2011, S. 493–496, doi:10.1038/nature10231
- Sandra Beleza et al.: The Timing of Pigmentation Lightening in Europeans. In: Molecular Biology and Evolution. Band 30, Nr. 1, 2013, S. 24–35,doi:10.1093/molbev/mss207
- Laura Geggel 2019-01-29T17:32:46Z History: Faces Re-Created of Ancient Europeans, Including Neanderthal Woman and Cro-Magnon Man. Abgerufen am 1. Dezember 2019 (englisch): „"their skin color to be at least like today's people living in North Africa, or in fact, a bit darker"“
- Nicholas G. Crawford et al.: Loci associated with skin pigmentation identified in African populations. In: Science. Band 358, Nr. 6365, 2017, eaan8433, doi:10.1126/science.aan8433
- New gene variants reveal the evolution of human skin color. Auf: sciencemag.org vom 12. Oktober 2017.
- Love Dalén u. a.: Partial genetic turnover in neandertals: continuity in the east and population replacement in the west. In: Molecular Biology and Evolution. 2012 doi:10.1093/molbev/mss074.
- Olaf Jöris, Martin Street: At the end of the 14C time scale – the Middle to Upper Paleolithic record of Western Eurasia. In: Dan S. Adler, Olaf Jöris (Hrsg.): Setting the Record Straight: Toward a Systematic Chronological Understanding of the Middle to Upper Paleolithic Boundary in Eurasia. In: Journal of Human Evolution. Band 55, 2008, S. 782–802 doi:10.1016/j.jhevol.2008.04.002.
- Ron Pinhasi u. a.: Revised age of late Neanderthal occupation and the end of the Middle Paleolithic in the northern Caucasus. In: PNAS. Band 108, Nr. 21, 2011, S. 8611–8616, doi:10.1073/pnas.1018938108.
- Qiaomei Fu, Mateja Hajdinjak, Oana Teodora Moldovan, Silviu Constantin, Swapan Mallick: An early modern human from Romania with a recent Neanderthal ancestor. In: Nature. Band 524, Nr. 7564, 13. August 2015, ISSN 0028-0836, S. 216–219, doi:10.1038/nature14558, PMID 26098372, PMC 4537386 (freier Volltext).
- Richard E. Green u. a.: A Complete Neandertal Mitochondrial Genome Sequence Determined by High-Throughput Sequencing. In: Cell. Band 134, Nr. 3, 2008, S. 416–426, doi:10.1016/j.cell.2008.06.021
- Ewen Callaway: Early European may have had Neanderthal great-great-grandparent. Auf: nature.com vom 13. Mai 2015, doi:10.1038/nature.2015.17534
- Frühe Europäer haben sich mit Neandertalern vermischt. Auf: mpg.de vom 22. Juni 2015, mit einer Abbildung des Unterkiefers Oase 1
- Erbgut des bisher ältesten modernen Menschen entschlüsselt. Max-Planck-Gesellschaft vom 22. Oktober 2014.
- Ann Gibbons: Ancient DNA pinpoints Paleolithic liaison in Europe. In: Science. Band 348, Nr. 6237, 2015, S. 847, doi:10.1126/science.348.6237.847
- Wighard von Koenigswald: Lebendige Eiszeit. Wissenschaftliche Buchgesellschaft, Darmstadt 2002, S. 167.
- Hervé Bocherens, Dorothée G. Drucker, Daniel Billioud, Marylène Patou-Mathise, and Bernard Vandermeersch: Isotopic evidence for diet and subsistence pattern of the Saint-Césaire I Neanderthal: review and use of a multi-source mixing model. In: Journal of Human Evolution. Band 49, Nr. 1, 2005, S. 71–87 doi:10.1016/j.jhevol.2005.03.003
- Michael P. Richards, Paul B. Pettitt, Mary C. Stiner, Erik Trinkaus: Stable isotope evidence for increasing dietary breadth in the European mid-Upper Paleolithic. In: PNAS. Band 98, Nr. 11, 2001, S. 6528–6532 doi:10.1073/pnas.111155298
- D. Drucker, H. Bocherens: Carbon and nitrogen stable isotopes as tracers of change in diet breadth during Middle and Upper Palaeolithic in Europe. In: International Journal of Osteoarchaeology. Band 14, Nr. 3–4, S. 162–177 doi:10.1002/oa.753
- M.P. Richards, R. Jacobi, C. Stringer, P.B. Pettitt, J. Cook: Isotope evidence for the intensive use of marine foods by Late Upper Palaeolithic humans. In: Journal of Human Evolution. Band 49, Nr. 3, 2005, S. 390–394 doi:10.1016/j.jhevol.2005.05.002
- P. B. Pettitt, M. Richards, R. Maggi, V. Formicola: The Gravettian burial known as the Prince („Il Principe“): new evidence for his age and diet. In: Antiquity. Band 77, Nr. 295, 2003, S. 15–19, Volltext (PDF)
- D. G. Drucker, D. Henry-Gambier: Determination of the dietary habits of a Magdalenian woman from St Germain-la-Rivière in southwestern France using stable isotopes. In: Journal of Human Evolution. Band 49, 2005, S. 19–35 doi:10.1016/j.jhevol.2005.02.007
- M. P. Richards, R. E. M. Hedges, R. Jacobi, A. Current, C. Stringer: FOCUS: Gough's Cave and Sun Hole Cave Human Stable Isotope Values Indicate a High Animal Protein Diet in the British Upper Palaeolithic. In: Journal of Archaeological Science. Band 27, Nr. 1, S. 1–3 doi:10.1006/jasc.1999.0520
- E. Garcia-Guixé, J. Martínez-Moreno, R. Mora, M. Núñez, M.P. Richards: Stable isotope analysis of human and animal remains from the Late Upper Palaeolithic site of Balma Guilanyà, southeastern Pre-Pyrenees, Spain. In: Journal of Archaeological Science. Band 36, Nr. 4, 2009, S. 1018–1026 doi:10.1016/j.jas.2008.12.001
- Wil Roebroeks: Time for the Middle to Upper Paleolithic transition in Europe. In: Journal of Human Evolution. Band 55, Nr. 5, 2008, S. 918–926 doi:10.1016/j.jhevol.2008.08.008
- Olaf Jöris, Martin Street: At the end of the 14C time scale – the Middle to Upper Paleolithic record of Western Eurasia. In: D. S. Adler, O. Jöris (Hrsg.): Setting the Record Straight: Toward a Systematic Chronological Understanding of the Middle to Upper Paleolithic Boundary in Eurasia. In: Journal of Human Evolution. Band 55, 2008, S. 782–802 doi:10.1016/j.jhevol.2008.04.002
- Thomas Higham u. a.: Problems with radiocarbon dating the Middle to Upper Palaeolithic transition in Italy. In: Quaternary Science Reviews. 28/13–14, 2009, S. 1257–1267 doi:10.1016/j.quascirev.2008.12.018
- João Zilhão u. a.: Analysis of Aurignacian interstratification at the Chatelperronian-type site and implications for the behavioral modernity of Neandertals. In: PNAS. Band 103, Nr. 33, 2006, S. 12643–12648 doi:10.1073/pnas.0605128103
- Paul Mellars, Brad Gravina, Christopher Bronk Ramsey: Confirmation of Neanderthal/modern human interstratification at the Chatelperronian type-site. In: PNAS. Band 104, Nr. 9, 2007, S. 3657–3662.
- Leif Steguweit (Hrsg.): Menschen der Eiszeit: Jäger – Handwerker – Künstler. Praehistorika, Fürth 2008, ISBN 978-3-937852-01-0, (PDF-Download)
- Victoria Cabrera Valdes, James L. Bischoff: Accelerator 14C dates for early upper paleolithic (basal Aurignacian) at El Castillo Cave (Spain). In: Journal of Archaeological Science. Band 16, Nr. 6, November 1989, S. 577–584, doi:10.1016/0305-4403(89)90023-X (Online [abgerufen am 16. Juni 2019]).
- J. Hahn: Kraft und Aggression. Die Botschaft der Eiszeitkunst im Aurignacien Süddeutschlands? In: Archaeologica Venatoria. Band 7, Tübingen 1986.
- Martin Porr: Reflections of human beings: The Aurignacian art of central Europe. Unpublished PhD Thesis, University of Southampton, 2002.
- H. R. Bell, M. Porr: ‘Rock-art’, ‘animism’ and two-way thinking. Towards a complementary epistemology in the understanding of material culture and ‘rock-art’ of hunting and gathering people. In: Journal of Archaeological Method and Theory. doi:10.1007/s10816-011-9105-4
- Schwarzbach, S. 247 ff.; Lamb, S. 129 ff.
- Cunliffe, S. 55–60; Clottes/Courtin: Cosquer, S. 33–40; Hoffmann, S. 91 ff.
- Pummel aus dem Eis. Auf: spiegel.de vom 14. April 2008.
- D. Caramelli, L. Milani, S. Vai, A. Modi, E. Pecchioli u. a.: A 28,000 Years Old Cro-Magnon mtDNA Sequence Differs from All Potentially Contaminating Modern Sequences. In: PLoS ONE. 3(7), 2008: e2700. doi:10.1371/journal.pone.0002700
- B. Bramanti u. a.: Genetic Discontinuity Between Local Hunter-Gatherers and Central Europe’s First Farmers. In: Science. Band 326, Nr. 5949, 2009, S. 137–140 doi:10.1126/science.1176869