Crenarchaeota
Die Crenarchaeota (vom griechischen crenos, Ursprung oder Quelle, veraltet: Eozyten, wissenschaftlich: Eocyta) sind einzellige Lebewesen und gehören zu der Domäne der Archaea. Früher wurden die Crenarchaeota ausschließlich für Extremophile gehalten (d. h. entweder (extrem) thermophil bzw. thermoacidophil oder psychrophil), jedoch zeigten weitere Untersuchungen, dass die Crenarchaeota zu den häufigsten Archaea im Meer gehören.[1] Diese Gruppe wird als mesophile, marine Gruppe I Crenarchaeota bezeichnet.
| Crenarchaeota | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
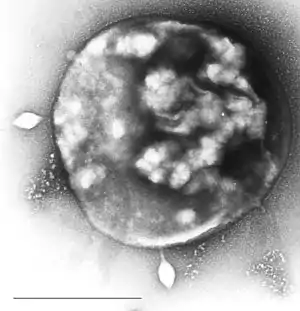
Sulfolobus, infiziert mit dem Sulfolobus-Virus STSV1 (ICTV: Sulfolobus spindle-shaped virus 1). | ||||||||||
| Systematik | ||||||||||
| ||||||||||
| Wissenschaftlicher Name | ||||||||||
| Crenarchaeota | ||||||||||
| George M. Garrit & John G. Holt, 2002 |
Eigenschaften
Innerhalb der Crenarchaeota findet man Organismen, die an extreme Umweltbedingungen angepasst und in Bereichen mit sehr hoher oder sehr niedriger Temperatur zu finden sind. Viele Vertreter leben im arktischen Plankton, wo sie bei Temperaturen oft unter 0 °C überleben. Diese Psychrophilen sind jedoch erst vereinzelt unter Laborbedingungen kultiviert worden (siehe auch Psychrophilie und Kryophile). Eine andere Gruppe innerhalb der Crenarchaeota lebt unter hyperthermischen Bedingungen, also bei Temperaturen von 80–110 °C, Pyrolobus fumarii lebt sogar bei 113 °C und überlebt auch einstündiges Autoklavieren. Viele der Crenarchaeota wie z. B. Sulfolobus acidocaldarius tolerieren auch hohe Säurekonzentrationen (pH-Werte von 1–2) und sind damit thermoacidophil.
Einer der bestcharakterisierten Organismen der Crenarchaeota ist Sulfolobus solfataricus. Ursprünglich wurde dieser Organismus aus schwefelhaltigen heißen Quellen in Italien isoliert und wächst bei 80 °C und einem pH-Wert von 2–4.[2]
Da in ihrem Lebensraum oft kein organisches Substrat vorhanden ist, leben viele von der Fixierung von Schwefel, Kohlenstoffdioxid oder Wasserstoff. Andere können jedoch auch organisches Material verstoffwechseln. Bei der Kohlenstoffdioxidassimilation verwenden sie entweder den 3-Hydroxypropionat/4-Hydroxybutyratzyklus (z. B. Sulfolobus) oder den Dicarboxylat/4-Hydroxybutyratzyklus (z. B. Desulfurococcales oder Thermoproteales).[3]
Die Vertreter der Crenarchaeota dienen den zwei Virusfamilien der Ligamenvirales als Wirt, den Rudiviridae und Lipothrixviridae.
Systematik
Innere Systematik
- Klasse Thermoprotei Reysenbach 2002
- Ordnung „Caldisphaerales“
- Familie „Caldisphaeraceae“
- Ordnung Cenarchaeales
- Familie „Cenarchaeaceae“
- Ordnung Desulfurococcales
- Familie Desulfurococcaceae
- Familie Pyrodictiaceae
- Ordnung Sulfolobales
- Familie Sulfolobaceae [en] (Gattungen: Acidianus, Metallosphaera, Saccharolobus, Stygiolobus, Sulfolobus, Sulfurisphaera, Sulfurococcus)
- Ordnung Thermoproteales
- Familie Thermoproteaceae
- Familie Thermofilaceae
- Ordnung „Caldisphaerales“
Nicht klassifiziert sind die Kladen „Soil Crenarchaeotic Group“ (SCG)[4][5] und „Miscellaneous Crenarchaeota Group“ (MCG).[4][6]
Äußere Systematik
In den letzten Jahren (Stand 2017) wurden Organismen anhand gefundener Genome nachgewiesen, die mit den Crenarchaeota verwandt sind. Zunächst wurden gefunden:
- Thaumarchaeota[7] Brochier-Armanet et al. 2008
- „Aigarchaeota“ Nunoura et al. 2010
- „Korarchaeota“ Barns et al. 1996
Sie wurden zusammen mit den Crenarchaeota vorläufig in eine sogenannte TACK-Supergruppe eingeordnet (nach den ersten Buchstaben der Thaum-, Aig-, Cren- und Korarchaeota). Nach diesem Vorschlag wurden noch weitere Archaeen aus dem Umfeld der Crenarchaeota entdeckt:
- „Geoarchaeota“ Kozubal et al. 2013
- „Bathyarchaeota“ Meng et al. 2014
Auch diese wurden unter die TACK-Supergruppe gestellt. Für weitere verwandte Taxa wurde eine eigene Supergruppe Asgard vorgeschlagen.
Für die damals bekannten Vertreter dieser beiden Supergruppen[8] wurde als umfassende taxonomische Einheit der Name „Proteoarchaeota“[9] (auch „Proteoarchaea“)[10] vorgeschlagen. Damit umfassen die Proteoarchaeota die gesamten Supergruppen TACK und Asgard (implizit auch deren obige neue Mitglieder).
Eozyten-Hypothese
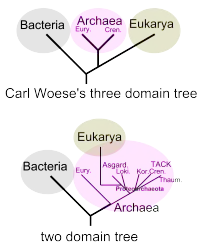
Die in den 1980er Jahren von James Lake vorgeschlagene Eozyten-Hypothese legt nahe, dass sich die Eukaryoten (Zellen mit komplexem Aufbau und Vielzeller wie Pflanzen, Pilze und Tiere, insbesondere auch der Mensch) aus den prokaryotischen Eocyten (ein alter Name für Crenarchaeota) entwickelten.[12][13]
Die V-Typ ATPase der Eukaryoten ähnelt der A-Typ ATP-Synthase der Archaeen – eine Tatsache, die einen Ursprung der Eukaryonten unter den Archaeen nahelegt. Die Ähnlichkeiten zu den bei Bakterien, Chloroplasten und Mitochondrien anzutreffenden F-Typ ATP-Synthasen sind dagegen weitaus geringer (das ausnahmsweise Auftreten des F-Typs bei einigen Archaeenlinien und des A-Typs bei einigen Bakterienlinien wird als Folge horizontalen Gentransfers angesehen).[14]
Proteasom genannte Proteinkomplexe haben offenbar bei dem Crenarchaeota-Mitglied Sulfolobus acidocaldarius in der Zellteilung die gleiche Funktion wie bei Eukaryoten.[15]
Ein Hinweis auf eine enge Beziehung zwischen Crenarchaeota und Eukaryoten ist das Vorhandensein eines Homologs der RNA-Polymerase-Untereinheit Rbp-8 in Crenarchaeota, die aber nicht bei den Euryarchaeota zu finden ist.[16]
Seit der Entdeckung weiterer den Crenarchaeota nahestehenden Archaeengruppen innerhalb der Supergruppe TACK wurde spekuliert, ob diese den Eukaryoten vielleicht noch näher stehen könnten als diese. Mit der Entdeckung der TACK-Schwestergruppe Asgard wurden Vertreter der Archaeen gefunden, die den Eukaryoten noch wesentlich näher stehen; möglicherweise sind die Eukaryoten sogar aus diesen hervorgegangen.[17][18] Diese Ergebnisse bestätigen und konkretisieren die Aussage der Endosymbiontentheorie bzgl. der Herkunft der Urkaryoten, auch wenn die Verwandtschaftsbeziehungen immer noch im Detail diskutiert werden.[11][19]
Eine im Herbst 2020 veröffentlichte Studie legt anhand von umfangreichen Genomanlysen tatsächlich nahe, dass – obwohl bisher noch keine primär amitochondrialen Eukaryoten gefunden wurden – die Vorfahren der Eukaryonten aus der Verwandtschaft der Asgard-Archaeen zuerst ihre komplexes Genom mit den zugehörigen Strukturen, und danach die Mitochondrien (oder Vorläufer davon) erworben haben.[20]
Weblinks
- microbewiki: Sulfolobus
- Traci Watson: The trickster microbes that are shaking up the tree of life, in: Nature vom 14. Mai 2019 (englisch), Trickser-Bakterien schütteln den Stammbaum des Lebens, in: Spektrum.de vom 20. Juni 2019 (deutsch)[21]
Einzelnachweise
- Brock: Biology of Microorganisms. 11. Edition. Prentice Hall, 2005, ISBN 0-13-144329-1.
- W. Zillig et al.: The Sulfolobus-„Caldariellard“ group: Taxonomy on the basis of the structure of DNA-dependent RNA polymerases. In: Arch. Microbiol. 125(3) 1980, S. 259–269, doi:10.1007/BF00446886.
- I. A. Berg et al.: Autotrophic carbon fixation in archaea. In: Nat Rev Microbiol. 8, 2010, PMID 20453874, S. 447–460, doi:10.1038/nrmicro2365.
- Rüdiger Ortiz Álvarez, Emilio O. Casamayor: High occurrence of Pacearchaeota and Woesearchaeota (Archaea superphylum DPANN) in the surface waters of oligotrophic high-altitude lakes. In: Environmental Microbiology Reports. 8, Nr. 2, April 2016, S. 210–217. doi:10.1111/1758-2229.12370. PMID 26711582.
- Archaea (domain), auf: Silva Databases
- Jun Meng, Jun Xu, Dan Qin, Ying He, Xiang Xiao, Fengping Wang: Genetic and functional properties of uncultivated MCG archaea assessed by metagenome and gene expression analyses, in: ISME Journal, Band 8, S. 650–659(2014), 10. Oktober 2013, doi:10.1038/ismej.2013.174
- Céline Brochier-Armanet, Bastien Boussau, Simonetta Gribaldo, Patrick Forterre: Mesophilic crenarchaeota: Proposal for a third archaeal phylum, the Thaumarchaeota. In: Nature Reviews Microbiology. Band 6, Nr. 3, 2008, S. 245–252, doi:10.1038/nrmicro1852, PMID 18274537.
- NCBI taxonomy page on Archaea
- Petitjean, C., Deschamps, P., López-García, P., and Moreira, D.: Rooting the Domain archaea by phylogenomic analysis supports the foundation of the new kingdom proteoarchaeota.. In: Genome Biol. Evol.. 7, 2014, S. 191–204. doi:10.1093/gbe/evu274.
- Jonathan Lombard: The multiple evolutionary origins of the eukaryotic N-glycosylation pathway, in: Biology Direct Band 11, Nr. 36, August 2016, doi:10.1186/s13062-016-0137-2
- C. J. Cox, P. G. Foster, R. P. Hirt, S. R. Harris, T. M. Embley: The archaebacterial origin of eukaryotes. In: Proc Natl Acad Sci USA. 105, Nr. 51, 2008, S. 20356–20361. bibcode:2008PNAS..10520356C. doi:10.1073/pnas.0810647105. PMID 19073919. PMC 2629343 (freier Volltext).
- (UCLA) The origin of the nucleus and the tree of life, UCLA via Web-Archiv vom 7. Februar 2003
- Natalya Yutin, Eugene V. Koonin: The Dispersed Archaeal Eukaryome and the Complex Archaeal Ancestor of Eukaryotes. In: Cold Spring Harbor Perspectives in Biology. 6, Nr. 4, April 2014, S. a016188. doi:10.1101/cshperspect.a016188. PMID 24691961. PMC 3970416 (freier Volltext).
- E. Hilario E, J. P. Gogarten: Horizontal transfer of ATPase genes--the tree of life becomes a net of life. In: Bio Systems. 31, Nr. 2–3, 1993, S. 111–119. doi:10.1016/0303-2647(93)90038-E. PMID 8155843.
- Gabriel Tarrason Risa, Fredrik Hurtig, Sian Bray, Anne E. Hafner, Lena Harker-Kirschneck, Peter Faull, Colin Davis, Dimitra Papatziamou, Delyan R. Mutavchiev, Catherine Fan, Leticia Meneguello, Andre Arashiro Pulschen, Gautam Dey, Siân Culley, Mairi Kilkenny, Diorge P. Souza, Luca Pellegrini, Robertus A. M. de Bruin, Ricardo Henriques, Ambrosius P. Snijders, Andela Šaric, Ann-Christin Lindås, Nicholas P. Robinson, Buzz Baum: The proteasome controls ESCRT-III–mediated cell division in an archaeon, in: Science vom 7. August 2020, doi:10.1126/science.aaz2532, siehe auch Proteasom.
Evolutionary Insight: New Connection Discovered Between Primordial Organisms and Complex Life, auf: SciTechDaily vom 28. August 2020, Quelle: Lancaster University. - M. Kwapisz, F. Beckouët, P Thuriaux: Early evolution of eukaryotic DNA-dependent RNA polymerases. In: Trends Genet.. 24, Nr. 5, 2008, S. 211–215. doi:10.1016/j.tig.2008.02.002. PMID 18384908.
- Eva F. Caceres: Genomic and evolutionary exploration of Asgard archaea, Doctoral thesis, Uppsala University, Disciplinary Domain of Science and Technology, Biology, Department of Cell and Molecular Biology, 12. November 2019
- Katarzyna Zaremba-Niedzwiedzka et al.: Asgard archaea illuminate the origin of eukaryotic cellular complexity, in: Nature 541, S. 353–358 vom 19. Januar 2017, doi:10.1038/nature21031.
- Bram Henneman: Genomic diversity, lifestyles and evolutionary origins of DPANN archaea: Chapter 1: Introduction, Leiden University Repository, 5. Dezember 2019. Siehe Fig. 1.1
- Julian Vosseberg, Jolien J. E. van Hooff, Marina Marcet-Houben, Anne van Vlimmeren, Leny M. van Wijk, Toni Gabaldón, Berend Snel: Timing the origin of eukaryotic cellular complexity with ancient duplications, in: Nature Ecology & Evolution, 26. Oktober 2020, doi:10.1038/s41559-020-01320-z, dazu:
Timeline Unveiled for One of the Most Important and Puzzling Events in the Evolution of Life, auf: SciTechDaily vom 4. November 2020, Quelle: Institute for Research in Biomedicine (IRB Barcelona) – ‚Bakterien‘ ist hier im Sinn von ‚Prokaryoten‘ zu verstehen. - Die Bezeichnung ‚Bakterien‘ ist nicht ganz korrekt, bei den betrachteten Mikroben handelt es sich um Archaeen oder (nach Ansicht mancher Forscher) jedenfalls von den Bakterien verschiedene Proto-Eukaryonten.