Carnotaurus
Carnotaurus („Fleischfressender Stier“) ist eine Gattung theropoder Dinosaurier aus der Oberkreide Argentiniens. Bisher ist erst eine Art bekannt (Carnotaurus sastrei).
| Carnotaurus | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|

Skelettabguss von Carnotaurus im Chlupáč Museum in Prag | ||||||||||
| Zeitliches Auftreten | ||||||||||
| Oberkreide (Unteres Maastrichtium)[1] | ||||||||||
| 72 bis 69,9 Mio. Jahre | ||||||||||
| Fundorte | ||||||||||
| ||||||||||
| Systematik | ||||||||||
| ||||||||||
| Wissenschaftlicher Name | ||||||||||
| Carnotaurus | ||||||||||
| Bonaparte, 1985 | ||||||||||
| Art | ||||||||||
| ||||||||||

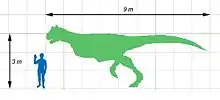
Charakteristisch für diesen zweibeinigen Fleischfresser waren ein sehr kurzer und tiefer Schädel sowie ein markantes Paar großer Stirnhörner. Bisher ist erst ein einziges Skelett gefunden worden, das jedoch bis auf die unteren Abschnitte der Beine und den Großteil des Schwanzes vollständig erhalten ist und sogar Abdrücke der Haut mit einschließt.
Carnotaurus wird zu den Abelisauridae gezählt, einer Gruppe mittelgroßer bis großer Theropoden, deren Fossilien in der Kreide Südamerikas, Afrikas und Indiens gefunden wurden. Bereits 1985 wissenschaftlich beschrieben, war Carnotaurus lange der einzige Vertreter der Abelisauridae, von dem ein nahezu vollständiges Skelett bekannt war. Verschiedene biomechanische Studien bieten Hinweise auf mögliche innerartliche Kämpfe, auf die Ernährung sowie auf die Laufgeschwindigkeit dieses Tieres.
Merkmale
Fund und Körpergröße
Das einzige Skelett ist im ursprünglichen anatomischen Verbund vorgefunden worden. Der mittlere und hintere Abschnitt des Schwanzes sowie die unteren Bereiche beider Hinterbeine fehlen, da sie bereits vor der Entdeckung des Skeletts durch Verwitterung zerstört wurden.[2] Das Skelett lag bei seiner Entdeckung auf der rechten Seite und zeigte einen über den Rücken gekrümmten Hals[3] – diese auch als death pose („Totenposition“) bezeichnete Haltung ist typisch für Fossilien langhälsiger Landwirbeltiere und vermutlich auf das postmortale Wirken eines vorgespannten, entlang der Wirbelsäule verlaufenden Bandes (das Ligamentum elasticum interlaminare) zurückzuführen.[4] Es handelt sich um ein erwachsenes Individuum, wie aus dem Verschmelzungsgrad der Knochennähte des Hirnschädels geschlossen werden kann.[5]
Das einzige bekannte Exemplar wird auf eine Körperlänge von 8 bis 9 Metern geschätzt.[6][7] Damit war Carnotaurus einer der größten bekannten Vertreter der Abelisauridae[8] – lediglich Ekrixinatosaurus und eventuell der nur durch einen Schädel bekannte Abelisaurus könnten ähnlich groß oder noch größer gewesen sein.[9][10][8] Gewichtsschätzungen variieren je nach Studie und verwendeter Methode. Dale Russell schätzt das Gewicht anhand der Länge des Oberschenkelknochens auf 1,35 Tonnen.[11] Gerardo Mazzetta und Kollegen (1998) berechnen das Gewicht mithilfe einer volumetrischen Methode auf 1,5 Tonnen, indem sie das Körpervolumen berechnen und dieses mit einer geschätzten mittleren Gewebedichte von 1000 kg pro m³ multiplizieren.[12] Gerardo Mazzetta und Kollegen (2004) konnten ein Gewicht von 2,1 Tonnen errechnen, basierend auf Längenmaßen verschiedener Gliedmaßenknochen.[13]
Schädel
Der Schädel ist fast vollständig überliefert und gut erhalten, obwohl er insbesondere im Schnauzenbereich seitlich zerdrückt ist. Gemessen von der Schnauzenspitze (Zwischenkieferbein) bis zur Hinterseite des Quadratbeins ist er 59,6 cm lang, und gemessen von der Oberseite des Hinterhauptbeins (Occipitalkamm) bis zur Unterseite des Jochbeins 42,5 cm hoch.[14] Damit war der Schädel im Verhältnis kürzer und tiefer als bei allen anderen bekannten Abelisauriden. Ein solches einzigartiges Merkmal wird auch als Autapomorphie bezeichnet und dient der Abgrenzung dieser Gattung von anderen Gattungen.[8] Mit Ausnahme des Hirnschädels zeigte der Schädel zahlreiche lose Knochennähte, was eine hohe Beweglichkeit der Schädelknochen zueinander (Schädelkinese) anzeigt[15]. Die Augen waren verhältnismäßig klein[16] und befanden sich im oberen Bereich der im Umriss schlüssellochartig geformten Augenhöhlen. Dieser obere Bereich war leicht nach vorne rotiert, was möglicherweise ein räumliches Sehen erlaubte[15][17]. Die Schnauze war relativ breit und gerundet, und nicht spitz zulaufend wie bei den meisten ursprünglichen Theropoden wie beispielsweise Ceratosaurus[18].

Die Oberfläche des Gesichtsschädels war mit zahlreichen Unebenheiten wie kleinen Gruben und Fortsätzen texturiert. Diese Texturierung war auf dem paarigen Nasenbein besonders ausgeprägt und war in ähnlicher Form bei verschiedenen anderen Abelisauriden vorhanden. Dieses Merkmal könnte darauf hinweisen, dass die den Gesichtsschädel überdeckende Haut hornig war. Über den Augen befand sich ein Paar großer, sehr dicker Stirnhörner, die vom paarigen Stirnbein gebildet werden. Derartige Stirnhörner fanden sich bei keinem anderen Theropoden, weshalb dieses Merkmal zur Abgrenzung zu anderen Gattungen dient (Autapomorphie). Diese kurzen und auf der Oberseite flachen Hörner waren laterodorsal (zur Seite und nach oben) ausgerichtet und maßen eine Länge von knapp 15 cm[5]. Möglicherweise bildeten sie den knöchernen Ansatz eines Überzugs aus Hornsubstanz, wodurch die Hörner beim lebenden Tier deutlich länger gewesen sein könnten.[17]
Die Zähne erschienen relativ schlank[16] und waren länger als bei vielen anderen Abelisauriden, die typischerweise sehr kurze Zähne aufwiesen[18]. Auf jeder Seite des Oberkiefers saßen 12 Zähne[19]; im paarigen Zwischenkieferbein saßen wie bei anderen Abelisauriden je vier weitere Zähne. Auf jeder Seite des Unterkiefers saßen 15 Zähne.[20] Die Kiefer waren von der Seite betrachtet stärker gebogen als bei anderen Abelisauriden, wie die obere Kante des Unterkiefers zeigt. Der Oberkiefer erscheint am gefundenen Schädel jedoch stärker gekrümmt, als er beim lebenden Tier tatsächlich war, weil das Zwischenkieferbein als Folge der seitlichen Zerdrückung des Schädels nach oben auf das Nasenbein aufgeschoben ist.[8] Der Unterkiefer war schlank und fragil gebaut[16] – so war das Dentale (der zahntragende, vordere Knochen des Unterkiefers) durch lediglich zwei Kontaktpunkte mit den hinteren Kieferknochen verbunden.[20]
Rumpfskelett und Gliedmaßen
Die Wirbelsäule ist bis zum sechsten Schwanzwirbel vollständig erhalten und im ursprünglichen anatomischen Verbund vorgefunden worden. Die hinteren Schwanzwirbel fehlen, lediglich der vermutlich zwölfte Schwanzwirbel ist unvollständig erhalten.[20] Die Wirbelsäule besteht aus zehn Halswirbeln, zwölf Rückenwirbeln und sieben Kreuzbeinwirbeln. Die Halswirbel zeigten nur sehr kleine Dornfortsätze, während die beiden hinter den Dornfortsätzen befindlichen Fortsätze, die Epipophysen, besonders groß waren und die höchsten Punkte der Wirbel bildeten. Somit wurde die Halswirbelsäule von oben betrachtet nicht durch die einzelne Reihe von Dornfortsätzen, sondern durch eine Doppelreihe von großen Epipophysen dominiert. Die vergrößerten Epipophysen weisen darauf hin, dass die für seitliche Bewegungen des Halses zuständigen Muskeln kräftig entwickelt waren. Obwohl auch bei anderen Abelisauriden vorhanden, war dieses Merkmal bei Carnotaurus sehr stark ausgeprägt.[21][22] Die Schwanzwirbel wiesen eine für Theropoden sehr ungewöhnliche Morphologie auf: So waren die Querfortsätze (seitliche Auswüchse der Wirbelbögen) nicht horizontal ausgerichtet, sondern zeigten steil nach oben. In einigen Schwanzwirbeln waren diese Fortsätze höher als die zentralen Dornfortsätze. Die Querfortsätze waren nach hinten geneigt und zeigten ein nach vorne gerichtetes, halbmondförmiges Ende, das vermutlich mit dem Querfortsatz des vorangehenden Wirbels in Verbindung stand. Diese speziellen Anpassungen waren bei Carnotaurus besonders extrem ausgeprägt, fanden sich jedoch auch bei anderen südamerikanischen Abelisauriden (Aucasaurus, Ilokelesia und Skorpiovenator), während sie bei Abelisauriden aus Madagaskar und Indien fehlten. Möglicherweise deuten diese Anpassungen auf eine vergrößerte Muskulatur zum Antrieb der Beine (s. u.).[23]
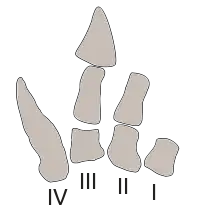
Die Arme waren lediglich als extrem zurückgebildete, verkürzte und vermutlich funktionslose[24] Reste ausgebildet. Bemerkenswert war außerdem der extrem verkürzte Unterarm, der lediglich einen Viertel der Länge des Oberarms ausmachte. Dieser Bauplan war für Vertreter der Abelisauridae typisch – im Vergleich mit verwandten Gattungen waren die Arme von Carnotaurus jedoch proportional noch kürzer und zudem robuster gebaut. Die Hände von Carnotaurus wurden im anatomischen Verbund vorgefunden, obwohl einige Knochen aus der ursprünglichen Position verschoben wurden oder fehlen. Die Hand war vierstrahlig und ebenfalls stark zurückgebildet. Wie bei Aucasaurus grenzten die Mittelhandknochen direkt an den Unterarm; Handwurzelknochen fehlten gänzlich. Der vierte Mittelhandknochen war der größte Knochen der Hand und mehr als doppelt so groß wie der zweite Mittelhandknochen – im Unterschied zu dem nahe verwandten Aucasaurus, bei dem der erste Mittelhandknochen der größte war. Das untere Ende des vierten Mittelhandknochens war konisch geformt und zeigte keine Gelenkfläche, weshalb im vierten Handstrahl offensichtlich keine Fingerknochen vorhanden waren. Dem ersten Handstrahl fehlten vermutlich ebenfalls Fingerknochen, während im zweiten Handstrahl eine direkt an den zweiten Mittelhandknochen angrenzende Kralle und im dritten Handstrahl ein Fingerknochen sowie eine Kralle vorhanden waren.[25]
Die Beckenknochen waren insgesamt verhältnismäßig lang, schlank und gerade. Von den Hinterbeinen sind lediglich beide Oberschenkelknochen sowie jeweils das oberste Drittel beider Schienbeine erhalten geblieben, die unteren Bereiche der Beine und Füße fehlen. Die Beine waren verhältnismäßig lang und schlank; so maß der linke Oberschenkelknochen eine Länge von 103 cm, während der Knochenschaft nur durchschnittlich 11 cm im Durchmesser maß.[11]
Haut
Carnotaurus ist der erste Fund eines großen theropoden Dinosauriers, bei dem die Haut relativ vollständig überliefert ist.[3] So sind Hautabdrücke von verschiedenen Körperregionen von der rechten Seite des Skeletts überliefert, die teilweise direkt mit den Knochen assoziiert sind. Jeweils ein Fragment stammt vom Unterkiefer[3] und vom vorderen Abschnitt des Halses, während ein anderes Fragment dem Schultergürtel und zwei weitere Fragmente dem Brustkorb zuzurechnen sind. Das größte überlieferte Fragment stammt aus dem vorderen Bereich des Schwanzes.[26] Ursprünglich war die rechte Schädelseite ebenfalls mit verschiedenen Fragmenten von Hautabdrücken bedeckt, was sich allerdings erst nach der Präparation des Schädels herausstellte, in deren Folge diese Fragmente zum Großteil verlorengingen[3].
Die Haut unterschied sich in verschiedenen Körperregionen nur geringfügig. Sie bestand aus einem Mosaik aus polygonalen, sich nicht überlappenden Schuppen mit einem Durchmesser von etwa 5 mm, das von dünnen, parallelen Furchen unterbrochen wurde.[27] Im Unterschied zu anderen bekannten Hautabdrücken von Theropoden zeigten die von Carnotaurus konische, knopfartige Strukturen, die einen typischen Durchmesser von 4 bis 5 cm zeigten und einen flachen Kamm auf der Oberseite aufwiesen. Diese Strukturen waren mit einem Abstand von 8 bis 10 cm zueinander in unregelmäßigen Reihen entlang der Seiten des Tieres angeordnet.[26][3] Vermutlich bestanden diese Strukturen aus mehreren, zu einem Gesamtkomplex verdichteten Einzelschuppen. Es konnten keine Verknöcherungen in Verbindung mit diesen Strukturen nachgewiesen werden.[3]
Systematik
| Kladogramm, vereinfacht nach Canale und Kollegen, 2009:[28] | ||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||
| Systematische Position von Carnotaurus |
Carnotaurus ist ein Vertreter der Abelisauridae, einer auf die südlichen Landmassen (Gondwana) beschränkten Gruppe innerhalb der Ceratosauria. Carnotaurus gilt als der am stärksten spezialisierte und abgeleitete (fortgeschrittene) Vertreter dieser Gruppe: So waren verschiedene Merkmale dieser Gruppe, wie die Verkürzung des Schädels und die Verkürzung der Arme, bei Carnotaurus stärker ausgeprägt als bei jedem anderen Abelisauriden.[1]
Die Verwandtschaftsbeziehungen innerhalb der Abelisauridae sind stark umstritten. Als nächster Verwandter (Schwestergattung) von Carnotaurus gilt je nach Studie meist entweder Majungasaurus[29][30][31] oder Aucasaurus[32][33][34].[35] Diese Zweideutigkeit liegt vor allem darin begründet, dass der Schädel von Aucasaurus nur wenig bekannt ist.[35][1] Eine andere Studie vermutet, dass Carnotaurus weder mit Aucasaurus noch mit Majungasaurus näher verwandt ist, sondern die Schwestergattung von Ilokelesia bildet[35].
Carnotaurus ist die namensgebende Gattung zweier Untergruppen der Abelisauridae, die jedoch nicht von allen Forschern akzeptiert und verwendet werden: der Carnotaurinae und der Carnotaurini. Die Carnotaurinae sollen alle abgeleiteten Vertreter der Abelisauridae zusammenfassen, unter Ausschluss des basalen (ursprünglichen) Abelisaurus[36]. Die Gruppe Carnotaurini wird von denjenigen Forschern verwendet, die Aucasaurus als den nächsten Verwandten von Carnotaurus klassifizieren, und beschreibt die von diesen beiden Gattungen gebildete Klade[32].
Forschungsgeschichte und Namensgebung
Das Skelett (Holotyp, Exemplarnummer MACN-CH 894) wurde 1984 von einer Expedition unter der Leitung von José Bonaparte geborgen.[1] Diese Expedition war die achte in Patagonien durchgeführte paläontologische Expedition eines von der National Geographic Society finanzierten Projekts über die jurassische und kreidezeitliche terrestrische Wirbeltierfauna Südamerikas.[2] Der Fundort befindet sich auf dem Land der Farm „Pocho Sastre“ nahe Bajada Moreno im Departamento Telsen in der argentinischen Provinz Chubut. Die Fossilien waren in einem sehr harten Gestein – einer Konkretion aus Hämatit – eingebettet, was ihre Präparation verkomplizierte und verlängerte.[16][37] Heute werden sie in der Sammlung des Museo Argentino de Ciencias Naturales Bernardino Rivadavia in Buenos Aires aufbewahrt.[17]
José Bonaparte veröffentlichte 1985 eine vorläufige Beschreibung des Schädels und benannte die neue Art und Gattung Carnotaurus sastrei.[37] Der Name Carnotaurus (lat. carno – „Fleisch“; lat. taurus – „Stier“) bedeutet so viel wie „Fleischfressender Stier“ und deutet auf die ungewöhnlichen Stirnhörner dieses Fleischfressers.[38] Im Jahr 1990 veröffentlichten José Bonaparte zusammen mit Fernando Novas und Rodolfo Coria eine umfangreichere Beschreibung des Schädels sowie des Postcranialskeletts.[2] Jüngere, auf diese Gattung fokussierte Studien behandeln die Biomechanik und stellen paläobiologische Implikationen auf oder beschreiben Skelettpartien wie den Hirnschädel[39] oder die Arme,[25] auf deren Anatomie in den vorangegangenen Beschreibungen nicht oder nur unzureichend eingegangen wurde. Carnotaurus war bis zu der Entdeckung von Aucasaurus der am vollständigsten überlieferte Vertreter der Abelisauridae. Heute sind mit Aucasaurus, Majungasaurus und Skorpiovenator weitere, gut erhaltene Abelisauriden-Funde bekannt, die es ermöglichen, verschiedene Merkmale des Carnotaurus-Skeletts, deren Interpretation bislang umstritten war, neu zu untersuchen.[8]
Paläohabitat
Obwohl der Fundort ursprünglich dem obersten Bereich der Gorro-Frigio-Formation zugeschrieben wurde, die an der Wende von Unter- und Oberkreide (Albium oder Cenomanium) zur Ablagerung kam,[17] wird er heute dem deutlich jüngeren unteren Bereich der La-Colonia-Formation zugeschrieben, der erst am Ende der Oberkreide im Unteren Maastrichtium ablagerte.[21][1]
Die La-Colonia-Formation ist an den Südhängen des Nordpatagonischen Massivs (Comarca Norpatagónica) weiträumig aufgeschlossen und besteht aus kontinentalen bis randmarinen Sedimentgesteinen. Der östliche Teil der Formation kann in drei Faziesassoziationen unterteilt werden. Der Großteil der Wirbeltierfossilien der Formation einschließlich des Carnotaurus-Skeletts stammen aus der zweiten Faziesassoziation:[40] Funde schließen sowohl terrestrische als auch aquatische Wirbeltiere mit ein und umfassen die Überreste von Lungenfischen aus der Gruppe der Ceratodontidae, Krokodile, Schildkröten, Plesiosaurier, Eidechsen, Schlangen, Dinosaurier und Säugetiere[41]. Der Ablagerungsraum (Paläoenvironment) der zweiten Faziesassoziation wird als Ästuar, als Watt oder als Küstenebene interpretiert. Das Klima zur Zeit der Ablagerung war vermutlich saisonal geprägt, wobei sich Trockenzeiten mit Regenzeiten abwechselten.[40]
Paläobiologie
Funktion der Stirnhörner und der Haut

Carnotaurus ist der einzige bekannte Theropode mit großen Stirnhörnern. Verschiedene Hypothesen versuchen die Funktion dieser Stirnhörner zu klären. Häufig diskutiert werden mögliche Funktionen als Waffe oder als Stoßdämpfer bei innerartlichen Kämpfen; die Hörner könnten jedoch auch eine visuelle Funktion gehabt und der Zurschaustellung gedient haben.[42] Gerardo Mazzetta und Kollegen (1998) schlagen alternativ eine Funktion als Jagdwaffe vor. So könnten die Hörner zum Töten oder Verletzen von kleinen Beutetieren verwendet worden sein. Eine solche Funktion von Hörnern ist aus dem Tierreich allerdings bislang nicht beschrieben worden.[43]
Bereits Gregory Paul (1988) vermutet, dass die Stirnhörner bei Kämpfen mit Artgenossen verwendet wurden. Wie dieser Autor angibt, könnten die stark verkleinerten Augenhöhlen das Verletzungsrisiko der Augen bei derartigen Kämpfen verringert haben.[16] Daniel Chure (1998) hält dagegen, dass die Stirnhörner sehr breit waren und eine flache Oberseite zeigen, was darauf hinweist, dass mögliche Kämpfe vor allem durch Stoßen mit der Schädeloberseite ausgetragen worden wären. Außerdem merkt er an, dass die verkleinerten Augenhöhlen nicht mit Kämpfen in Verbindung gebracht werden könnten – so würden andere Großtheropoden ohne Hörner, wie Abelisaurus, Acrocanthosaurus und Carcharodontosaurus, ebenfalls verkleinerte Augenhöhlen aufweisen.[44]
Gerardo Mazzetta und Kollegen (1998) kommen zu dem Ergebnis, dass die Halsmuskulatur die bei frontalen Zusammenstößen der Schädel auftretenden Kräfte absorbieren konnte. So war die Halsmuskulatur stark genug, um einen Stoß abzufedern, der entsteht, wenn zwei Tiere mit einer Laufgeschwindigkeit von je 5,7 m/s mit den Schädeln zusammenstoßen. Ein ähnlicher Wert wird für das rezente Breitmaulnashorn angenommen.[43]
Eine jüngere Studie von Gerardo Mazzetta und Kollegen (2009) untersucht mithilfe einer an einem digitalen dreidimensionalen Schädelmodell durchgeführten Finite-Elemente-Analyse, wie sich der Schädel beim Beißen sowie bei möglichen Kämpfen mechanisch verhält. Laut dieser Studie war der Schädel nicht in der Lage, heftigen frontalen Stößen zu widerstehen, konnte aber auf den Hörnern lastende Kompressionskräfte effektiv verteilen, ohne das Gehirn zu beeinträchtigen. Die Forscher vermuten deshalb, dass Kämpfe nicht durch Ringen, sondern vor allem durch Schieben mit den Hörnern ausgetragen wurden, ähnlich wie bei rezenten Rindern wie den Bisons.[42]
Stephen Czerkas (1997) vermutet, dass die konischen Hautstrukturen, die in Reihen die Seiten des Tieres bedeckten, zum Schutz bei Kämpfen gegen Artgenossen und anderen Theropoden dienten. Dieser Autor verweist auf den Hals der Grünen Leguane (Iguana), der ähnliche Strukturen zeigt, die begrenzten Schutz bei Revierkämpfen bieten.[3]
Funktion der Kiefer und Schädelkinese
Gerardo Mazzetta und Kollegen (1998, 2009) rekonstruieren die Kiefermuskulatur von Carnotaurus und kommen zu dem Schluss, dass der Biss von Carnotaurus im Vergleich mit anderen Theropoden wie Ceratosaurus oder Allosaurus weniger kraftvoll, aber schneller war.[43][42] Daraus schließen diese Forscher, dass Carnotaurus hauptsächlich relativ kleine Beutetiere jagte. So zeigen Studien an rezenten Krokodilen, dass die Fähigkeit zum schnellen Zupacken für das Jagen kleiner Beutetiere wichtiger ist als eine hohe Bisskraft. Gerardo Mazzetta und Kollegen (2009) kommen außerdem zu dem Schluss, dass der Schädel darauf ausgelegt war, großen Belastungen zu widerstehen, wie sie beispielsweise beim Zerren großer Beutestücke auftreten. Diese Forscher vermuten daher, dass Carnotaurus auch zum Jagen größerer Beutetiere in der Lage war.[42]
Forscher um François Therrien (2005) widersprechen den Ergebnissen von Gerardo Mazzetta und Kollegen. In ihrer Studie über die Biomechanik des Unterkiefers verschiedener Theropoden kommen diese Forscher zu dem Schluss, dass die Kiefer von den Abelisauriden Majungasaurus und Carnotaurus Analogien mit denen des Komodowarans aufwiesen. So würde die Stärke der Unterkiefer bzw. die Biegefestigkeit nach vorne hin linear abnehmen – dieses Muster finde sich bei Tieren, deren Kiefer nicht als Präzisionswerkzeug zum Fangen von kleinen Beutetieren geeignet sind. Die Forscher vermuten, dass die Kiefer von Carnotaurus und Majungasaurus auf das Zufügen von Schlitzwunden ausgelegt gewesen seien. Außerdem kommen diese Forscher zu dem Ergebnis, dass Carnotaurus einen kraftvollen Biss zeigte – so sei die Bisskraft doppelt so groß wie die des Mississippi-Alligators. Die Forscher schließen, dass Carnotaurus bevorzugt große Beutetiere jagte, die er möglicherweise aus dem Hinterhalt angriff.[45]
Robert Bakker (1998) vermutet ebenfalls, dass Carnotaurus an das Jagen sehr großer Beutetiere, insbesondere Sauropoden, angepasst war. Dieser Forscher erkennt in verschiedenen Merkmalen, wie der kurzen Schnauze, den relativ kleinen Zähnen und dem stark ausgeprägten Hinterhauptbein, konvergente Entwicklungen zu Allosaurus, welcher ebenfalls an das Jagen von Sauropoden angepasst gewesen sei. Robert Bakker zufolge könnten die Oberkiefer von Allosaurus und Carnotaurus wie eine gesägte Keule zum Zufügen von Wunden verwendet worden sein, wobei große Sauropoden mit wiederholten Attacken geschwächt worden seien.[46]
Gerardo Mazzetta und Kollegen (1998) vermuten, dass die Schädelkinese – die Beweglichkeit der Schädelknochen zueinander – ausgeprägter war als bei jedem anderen bekannten Dinosaurier. Die Forscher folgern, dass Carnotaurus kleinere Beutetiere möglicherweise als Ganzes verschluckt haben könnte. Im Gegensatz zu dem Bau des übrigen Schädels ist das hintere Schädeldach fest mit dem Hirnschädel verschmolzen, was zu einer erhöhten Stabilität dieses Bereichs führt. Die Forscher vermuten, dass dieser stabile Bereich zum Aufnehmen der durch die Hörner übertragenden Kräfte diente, wie sie bei möglichen Kämpfen auftreten würden.[15]
Laufgeschwindigkeit und Wendefähigkeit
Gerardo Mazzetta und Kollegen (1998, 1999) schätzen, inwieweit Carnotaurus zum schnellen Laufen fähig war, basierend auf der Widerstandsfähigkeit der Beinknochen gegen beim Laufen auftretende Biegemomente, die aus verschiedenen Maßen des Oberschenkelknochens und dem vermuteten Körpergewicht berechnet wird. Beinknochen werden durch Biegemomente, wie sie besonders beim schnellen Laufen auftreten, stark beansprucht. Die Forscher kommen zu dem Ergebnis, dass Carnotaurus ein schneller Läufer war, der besser an ein schnelles Laufen angepasst war als ein Mensch, aber deutlich weniger als ein Afrikanischer Strauß.[47][48]
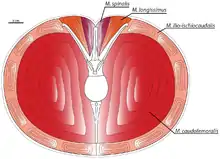
Die Schwanzwirbel zeigen steil nach oben gerichtete Querfortsätze. Scott Persons und Philip Currie (2011) geben an, dass dieses Merkmal den Platz vergrößerte, der für den Musculus caudofemoralis zur Verfügung stand. Der M. caudofemoralis war der für die Fortbewegung wichtigste Muskel und setzt am Vierten Trochanter des Oberschenkelknochens an; eine Kontraktion dieses Muskels zieht das Hinterbein nach hinten. Die Forscher kalkulieren den vergrößerten M. caudofemoralis von Carnotaurus auf ein Gewicht 111–137 kg pro Bein, somit war dieser Muskel bei Carnotaurus größer als bei jedem anderen auf dieses Merkmal hin untersuchten Theropoden. Basierend auf diesen Ergebnissen vermuten diese Forscher, dass Carnotaurus ein schneller Sprinter war und möglicherweise zu den schnellsten Großtheropoden zählte.[23]
Scott Persons und Philip Currie (2011) stellen außerdem fest, dass sich durch die nach oben gerichteten Querfortsätze der zur Verfügung stehende Platz für die epaxialen, entlang der Dornfortsätze verlaufenden Muskeln, den Musculus longissimus und den Musculus spinalis, verringerte. Diese beiden Muskel waren jedoch für die Stabilität als auch für die horizontale und vertikale Beweglichkeit des Schwanzes verantwortlich. Die verlängerten Querfortsätze waren an ihren oberen Enden miteinander artikuliert, was laut diesen Forschern die Stabilität des Schwanzes trotz der verringerten Epaxialmuskulatur sicherstellte. Dies schränkte jedoch die Beweglichkeit des Schwanzes weiter ein, und damit die Wendefähigkeit des Tieres: So verhielt sich die Hüfte und der Schwanz beim Wenden als eine Einheit, während andere Theropoden zuerst die Hüfte, und nachträglich den Schwanz wenden konnten.[23]
Weblinks
Belege
Hauptliteratur
- José F. Bonaparte, Fernando E. Novas, Rodolfo A. Coria: Carnotaurus sastrei Bonaparte, the horned, lightly built carnosaur from the Middle Cretaceous of Patagonia (= Contributions in Science. Nr. 416, ISSN 0459-8113). Natural History Museum of Los Angeles County, Los Angeles CA 1990, Digitalisat (PDF; 4,99 MB).
- Stephen A. Czerkas, Sylvia J. Czerkas: The Integument and Life Restoration of Carnotaurus. In: Donald L. Wolberg, Edmund Stump, Gary Rosenberg (Hrsg.): Dinofest International. Proceedings of a Symposium held at Arizona State University. Academy of Natural Sciences, Philadelphia PA 1997, ISBN 0-935868-94-1, S. 155–158.
- Gerardo V. Mazzetta, Richard A. Fariña, Sergio F. Vizcaíno: On the Paleobiology of the South American horned Theropod Carnotaurus Sastrei Bonaparte. In: Gaia. Revista de Geociências. Bd. 15, 1998, ISSN 0871-5424, S. 185–192.
- Gerardo V. Mazzetta, Adrián P. Cisilino, R. Ernesto Blanco, Néstor Calvo: Cranial mechanics and functional interpretation of the horned carnivorous dinosaur Carnotaurus sastrei. In: Journal of Vertebrate Paleontology. Bd. 29, Nr. 3, 2009, ISSN 0272-4634, S. 822–830, doi:10.1671/039.029.0313.
- Javier Ruiz, Angélica Torices, Humberto Serrano, Valle López: The hand structure of Carnotaurus sastrei (Theropoda, Abelisauridae): implications for hand diversity and evolution in abelisaurids. In: Palaeontology. Bd. 54, Nr. 6, 2011, ISSN 0031-0239, S. 1271–1277, doi:10.1111/j.1475-4983.2011.01091.x.
- W. Scott Persons, Philip J. Currie: Dinosaur Speed Demon: The Caudal Musculature of Carnotaurus sastrei and Implications for the Evolution of South American Abelisaurids. In: PLoS ONE. Bd. 6, Nr. 10, 2011, e25763, doi:10.1371/journal.pone.0025763.
Ergänzende Literatur
- Robert T. Bakker: Brontosaur Killers: Late Jurassic Allosaurids as Sabre-Tooth Cat Analogues. In: Gaia. Revista de Geociências. Bd. 15, 1998, S. 145–158, Digitalisat (PDF; 4,95 MB).
- José F. Bonaparte: A horned Cretaceous carnosaur from Patagonia. In: National Geographic Research. Bd. 1, 1985, ISSN 8755-724X, S. 149–151.
- José F. Bonaparte: Cretaceous tetrapods of Argentina. In: Münchner geowissenschaftliche Abhandlungen. Reihe A: Geologie und Paläontologie. Bd. 30, 1996, ISSN 0177-0950, S. 73–130.
- Jorge O. Calvo, David Rubilar-Rogers, Karen Moreno: A new Abelisauridae (Dinosauria: Theropoda) from northwest Patagonia. In: Ameghiniana. Bd. 41, Nr. 4, 2004, ISSN 0002-7014, S. 555–563, Digitalisat (PDF; 939,68 kB).
- Juan I. Canale, Carlos A. Scanferla, Federico L. Agnolin, Fernando E. Novas: New carnivorous dinosaur from the Late Cretaceous of NW Patagonia and the evolution of abelisaurid theropods. In: Die Naturwissenschaften. Bd. 96, Nr. 3, 2009, S. 409–414, doi:10.1007/s00114-008-0487-4.
- Ariana Paulina Carabajal: The braincase anatomy of Carnotaurus sastrei (Theropoda: Abelisauridae) from the Upper Cretaceous of Patagonia. In: Journal of Vertebrate Paleontology. Bd. 31, Nr. 2, 2011, S. 378–386, doi:10.1080/02724634.2011.550354.
- Matthew T. Carrano, Scott D. Sampson: The Phylogeny of Ceratosauria (Dinosauria: Theropoda). In: Journal of Systematic Palaeontology. Bd. 6, Nr. 2, 2008, ISSN 1477-2019, S. 183–236, doi:10.1017/S1477201907002246.
- Daniel Chure: On the Orbit of Theropod Dinosaurs. In: Gaia. Revista de Geociências. Bd. 15, 1998, S. 233–240, Digitalisat (PDF; 2,53 MB).
- Rodolfo A. Coria, Luis M. Chiappe, Lowell Dingus: A new close relative of Carnotaurus sastrei Bonaparte 1985 (Theropoda: Abelisauridae) from the Late Cretaceous of Patagonia. In: Journal of Vertebrate Paleontology. Bd. 22, Bd. 2, 2002, S. 460–465, doi:10.1671/0272-4634(2002)022[0460:ANCROC]2.0.CO;2.
- Martín D. Ezcurra, Federico L. Agnolin, Fernando E. Novas: An abelisauroid dinosaur with a non-atrophied manus from the Late Cretaceous Pari Aike Formation of southern Patagonia. In: Zootaxa. Nr. 2450, 2010, S. 1–25, Digitalisat (PDF; 815,22 kB).
- Gerardo V. Mazzetta, Richard A. Fariña: XIV jornadas Argentinas de paleontologia de vertebrados. Estimacion de la capacidad atlética de Amargasaurus cazaui Salgado y Bonaparte, 1991, y Carnotaurus sastrei Bonaparte, 1985 (Saurischia, Sauropoda-Theropoda). In: Ameghiniana. Bd. 36, Nr. 1, 1999, S. 105–106.
- Gerardo V. Mazzetta, Per Christiansen, Richard A. Fariña: Giants and Bizarres: Body Size of Some Southern South American Cretaceous Dinosaurs. In: Historical Biology. Bd. 16, Nr. 2/4, 2004, ISSN 0891-2963, S. 71–83, doi:10.1080/08912960410001715132, Digitalisat (PDF; 5747,66 kB).
- Fernando E. Novas: The age of dinosaurs in South America. Indiana University Press, Bloomington IN 2009, ISBN 978-0-253-35289-7.
- Rosendo Pascual, Francisco J. Goin, Pablo González, Alberto Ardolino, Pablo F. Puerta: A highly derived docodont from the Patagonian Late Cretaceous: evolutionary implications for Gondwanan mammals. In: Geodiversitas. Bd. 22, Nr. 3, 2000, ISSN 1280-9659, S. 395–414.
- Gregory S. Paul: Ceratosaurs. In: Gregory S. Paul: Predatory Dinosaurs of the World. A complete and illustrated Guide. Simon and Schuster, New York NY u. a. 1988, ISBN 0-671-61946-2.
- Achim G. Reisdorf, Michael Wuttke: Re-evaluating Moodie's Opisthotonic-Posture Hypothesis in Fossil Vertebrates. Part I: Reptiles—the taphonomy of the bipedal dinosaurs Compsognathus longipes and Juravenator starki from the Solnhofen Archipelago (Jurassic, Germany). In: Palaeobiodiversity and Palaeoenvironments. Band 92, Nr. 1, 2012, ISSN 1867-1594, S. 119–168, doi:10.1007/s12549-011-0068-y.
- Scott D. Sampson, Lawrence M. Witmer: Craniofacial Anatomy of Majungasaurus Crenatissimus (Theropoda: Abelisauridae) From the Late Cretaceous of Madagascar. Scott D. Sampson, David W. Krause (Hrsg.): Majungasaurus crenatissimus (Theropoda:Abelisauridae) from the late Cretaceous of Madagascar (= Journal of Vertebrate Paleontology. Bd. 27, Supplement to Nr. 2, 2007 = Society of Vertebrate Paleontology. Memoir. Nr. 8). Society of Vertebrate Paleontology, Northbrook IL 2007, S. 32–104, doi:10.1671/0272-4634(2007)27[32:CAOMCT]2.0.CO;2.
- Philip Senter: Vestigial skeletal structures in dinosaurs. In: Journal of Zoology. Bd. 280, Nr. 1, 2010, ISSN 0022-5460, S. 60–71, doi:10.1111/j.1469-7998.2009.00640.x.
- Paul C. Sereno, Jeffrey A. Wilson, Jack L. Conrad: New dinosaurs link southern landmasses in the Mid–Cretaceous. In: Proceedings of the Royal Society. Series B: Biological Sciences. Bd. 271, Nr. 1546, 2004, ISSN 0950-1193, S. 1325–1330, doi:10.1098/rspb.2004.2692.
- Juliana Sterli, Marcelo S. De La Fuente: A new turtle from the La Colonia Formation (Campanian–Maastrichtian), Patagonia, Argentina, with remarks on the evolution of the vertebral column in turtles. In: Palaeontology. Bd. 54, Nr. 1, 2011, S. 63–78, doi:10.1111/j.1475-4983.2010.01002.x.
- François Therrien, Donald M. Henderson, Christopher B. Ruff: Bite Me: Biomechanical Models of Theropod Mandibles and Implications for Feeding Behavior. In: Kenneth Carpenter (Hrsg.): The carnivorous dinosaurs. Indiana University Press, Bloomington IN u. a. 2005, ISBN 0-253-34539-1, S. 179–237.
- Ronald S. Tykoski, Timothy Rowe: Ceratosauria. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 47–70.
- Rubén D. Juárez Valieri, Juan D. Porfiri, Jorge O. Calvo: New Information on Ekrixinatosaurus novasi Calvo et al 2004, a giant and massively-constructed Abelisauroid from the „Middle Cretaceous“ of Patagonia. In: Jorge Calvo, Juan Porfiri, Bernardo González Riga, Domenica Dos Santos (Hrsg.): Paleontología y Dinosaurios desde América Latina. (= Serie Documentos y Testimonios. Aportes. Nr. 24). EDIUNC, Mendoza 2011, ISBN 978-950-39-0265-3, S. 161–169, Digitalisat (PDF; 643,51 kB).
- Jeffrey A. Wilson, Paul C. Sereno, Suresh Srivastava, Devendra K. Bhatt, Ashu Khosla, Ashok Sahni: A new abelisaurid (Dinosauria, Theropoda) from the Lameta Formation (Cretaceous, Maastrichtian) of India (= The University of Michigan. Contributions from the Museum of Paleontology. Bd. 31, Nr. 1, 2003, ISSN 0097-3556). The University of Michigan – Museum of Paleontology, Ann Arbor MI 2003, online.
Einzelnachweise
- Novas 2009, S. 276–279
- Bonaparte et al. 1990, S. 2
- Czerkas 1997
- Reisdorf und Wuttke 2012, S. 159
- Carabajal 2011, S. 379
- Bonaparte et al. 1990, S. 38
- Valieri und Porfiri 2011, S. 162
- Carrano und Sampson, S. 191
- Calvo et al. 2004, S. 556
- Valieri und Porfiri 2011, S. 163
- Bonaparte et al. 1990, S. 28–31
- Mazzetta et al. 1998, S. 187
- Mazzetta et al. 2004, S. 79
- Bonaparte et al. 1990, S. 8
- Mazzetta et al. 1998, S. 188–191
- Paul 1988, S. 284–285
- Bonaparte et al. 1990, S. 3–5
- Sampson und Witmer 2007, S. 95–96
- Novas 2009, S. 255
- Bonaparte et al. 1990, S. 6–16
- Bonaparte 1996, S. 89
- Novas 2009, S. 257
- Persons und Currie 2011, S. 1–7
- Senter 2010, S. 64
- Ruiz et al. 2011
- Bonaparte et al. 1990, S. 32
- Novas 2009, S. 264–266
- Canale et al. 2009
- Sereno et al. 2004
- Tykoski und Rowe 2004, S. 65
- Wilson et al. 2003, S. 25
- Coria et al. 2002, S. 460
- Canale et al. 2009, S. 4
- Ezcurra et al. 2010, S. 14
- Carrano und Sampson, S. 200, 202
- Paul Sereno: Carnotaurinae. (Nicht mehr online verfügbar.) In: Taxon Search. Archiviert vom Original am 16. Mai 2012; abgerufen am 4. August 2014.
- Bonaparte 1985
- Ben Creisler: Dinosauria Translation and Pronunciation Guide. Archiviert vom Original am 11. November 2011; abgerufen am 4. August 2014.
- Carabajal 2011
- Pascual et al. 2000, S. 398–400
- Sterli et al. 2011, S. 64–65
- Mazzetta et al. 2009
- Mazzetta et al. 1998, S. 190–191
- Chure 1998, S. 237
- Therrien et al. 2005, S. 179–198, 228
- Bakker 1998, S. 152–157
- Mazzetta und Fariña 1999
- Mazzetta et al. 1998, S. 186, 190