Trachodon-Mumie
Die Trachodon-Mumie ist eine natürlich entstandene Mumie eines Dinosauriers. Sie wurde 1908 in Wyoming (USA) entdeckt und wird der Dinosaurier-Art Edmontosaurus annectens (ursprünglich Trachodon annectens) zugeschrieben, einem Vertreter der Hadrosauridae („Entenschnabelsaurier“). Die Trachodon-Mumie ist das erste von selten gefundenen Dinosaurier-Fossilien, das umfangreiche, den Körper umhüllende Hautabdrücke zeigt, was zur Bezeichnung „Dinosaurier-Mumie“ führte.[1] Die ungewöhnlich gute Erhaltung dieses detailreichen Fossils hat die wissenschaftlichen Vorstellungen zur Biologie der Hadrosauriden maßgeblich geprägt. Zwischen den Fingern der Mumie vorgefundene Hautabdrücke wurden als Schwimmhäute interpretiert, was als entscheidendes Argument für eine aquatische Lebensweise der Hadrosauriden herangezogen wurde. Heute gilt diese Theorie als widerlegt.
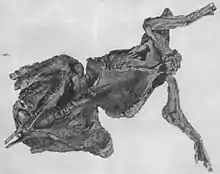
Die Mumie wurde von der Sternberg-Familie (Charles H. Sternberg und Söhne) entdeckt und geborgen, die für ihre zahlreichen Funde, insbesondere von Dinosaurier-Fossilien, berühmt ist. Heute ist die Mumie im American Museum of Natural History (AMNH) in New York ausgestellt, die Exemplarnummer lautet AMNH 5060.[2] 1910 entdeckte die Sternberg-Familie eine zweite Edmontosaurus-Mumie im selben Fundgebiet, die sie an das Senckenberg Naturmuseum in Frankfurt am Main (Senckenberg-Mumie) verkaufte, wo sie noch heute ausgestellt ist.[1]
Entdeckung


Die Trachodon-Mumie wurde 1908 von Charles Hazelius Sternberg und seinen drei Söhnen George, Charles und Levi entdeckt und geborgen. Sternberg war ein unabhängiger Fossilienjäger, der seine Funde verschiedenen Museen in Nordamerika und Europa zum Verkauf anbot. Seine Söhne arbeiteten anfangs als Assistenten ihres Vaters, stiegen später jedoch selbst zu anerkannten Paläontologen auf.[3] 1908 wollte Sternberg in ein neues Arbeitsgebiet vorstoßen, das Lance-Creek-Gebiet im östlichen Wyoming. Vorab bemühte er sich, Abnehmer für zu erwartende Fossilfunde zu finden, und schrieb an das British Museum of Natural History, er wisse, wo sich in Wyoming ein Fossil des gehörnten Dinosauriers Triceratops finden ließe. Das Museum willigte ein, es würde gute Fossilfunde aufkaufen, sollten welche gemacht werden.[4] Die Sternberg-Expedition brach zu Beginn des Frühjahrs von ihrem Familiensitz in Kansas auf,[4] das Lance-Creek-Gebiet erreichte sie im Juli.[5] Geplant war die Erkundung eines etwa 1000 Quadratmeilen (2590 km²) großen, unbewohnten Gebiets nördlich des North Platte River und südlich des Cheyenne River in Converse County (heute Niobrara County).[4][5] Die hier vorherrschenden Badlands schließen Gesteine der späten Oberkreide (Maastrichtium) auf, die heute als Lance-Formation bekannt sind. Das Gebiet wurde zuvor bereits intensiv von paläontologischen Expeditionen erkundet;[4] vor dem Start seiner Suche erfuhr Sternberg, dass das American Museum of Natural History bereits seit Jahren erfolglos in diesem Gebiet arbeitete.[5]
Die ersten Wochen der Suche blieben erfolglos.[5] Sternberg schrieb: “Day after day hoping against hope we struggled bravely on. Every night the boys gave answer to my anxious inquiry, What have you found? Nothing.” (deutsch: „Tag um Tag, hoffend gegen alle Hoffnung, mühten wir uns eifrig weiter ab. Jeden Abend gaben die Jungen Antwort auf meine bange Erkundigung, Was habt ihr gefunden? Nichts.“)[5] Ende August stieß Sternberg schließlich auf ein aus dem Gestein herauswitterndes Horn; die anschließende Grabung brachte einen 198 Zentimeter langen Triceratops-Schädel zu Tage.[5][6] Kurz darauf entdeckte der älteste Sohn George aus dem Gestein herauswitternde Knochen, als er zusammen mit dem jüngsten Sohn Levi noch unerkundetes Gebiet ablief. Levi fand weitere Knochen in der unmittelbaren Nähe, die offenbar zum selben Skelett gehörten. Zu diesem Zeitpunkt hatte sich die Gruppe bis auf 65 Meilen (105 km) von ihrem Basislager entfernt, und die Nahrungsvorräte wurden knapp. Sternberg wies George und Levi an, vorsichtig den Sandstein oberhalb des Skeletts abzutragen, und brach mit Charles nach Lusk auf, um neue Vorräte einzukaufen und den Schädel an das British Museum zu verschicken. Am dritten Tag nach der Abreise des Vaters hatten George und Levi bereits erkannt, dass sie ein auf dem Rücken liegendes, offenbar vollständiges Exemplar vor sich hatten. Als George ein großes Stück Sandstein aus der Brustregion des Skeletts entnahm, entdeckte er zu seiner Überraschung einen perfekt erhaltenen Hautabdruck. 1930 schrieb George: “Imagine the feeling that crept over me when I realized that here for the first time a skeleton of a dinosaur had been discovered wrapped in its skin. That was a sleepless night for me.” (deutsch: „Stellen Sie sich das Gefühl vor, das mich überfiel, als ich erkannte, dass hier zum ersten Mal ein Dinosaurierskelett mitsamt seiner Haut entdeckt wurde. Das war eine schlaflose Nacht für mich.“)[3] Als Sternberg schließlich am fünften Tag zurückkehrte, waren George und Levi die Nahrungsmittel ausgegangen; die letzten beiden Tage mussten sie von ungesalzenen Kartoffeln leben. Nichtsdestotrotz hatten sie den aufliegenden Sandstein vollständig entfernt; die Grabung maß einen Umfang von 3,6 Meter Breite, 4,5 Meter Länge und 3 Meter Tiefe.[4][5]
Als Henry Fairfield Osborn, Leiter des American Museum of Natural History, von dem Fund erfuhr, schickte er unmittelbar einen Mitarbeiter nach Wyoming, um das Fossil für das Museum zu erwerben. Osborn wusste über die Vereinbarung zwischen Sternberg und dem British Museum, appellierte jedoch an Sternbergs Patriotismus und versprach eine dauerhafte Ausstellung des Fossils.[4] Schließlich kaufte das Museum die Mumie für 2.000 Dollar.[2] Im American Museum wurde die Freilegung der Hautabdrücke von dem Präparator Otto Falkenbach abgeschlossen,[7] anschließend wurde das Fossil von Osborn und Barnum Brown wissenschaftlich bearbeitet und schließlich im Museum ausgestellt. In der Ausstellung wird das von einer gläsernen Schutzhülle umfasste Exemplar in seiner ursprünglichen Fundlage gezeigt, auf die Rekonstruktion fehlender Teile wurde verzichtet.[2][4]
Bedeutung und systematische Einordnung


Der wissenschaftliche Wert der Mumie liegt in dem ungewöhnlich guten Erhaltungsgrad, dem Zusammenhang der Knochen in ihrem ursprünglichen anatomischen Verbund sowie in den umfangreichen Hautresten. Osborn schrieb 1911: “This truly wonderful specimen, therefore, nearly doubles our previous insight into the habits and life of a very remarkable group of reptiles.” (deutsch: „Dieses wirklich wundervolle Exemplar gewährt uns nahezu eine Verdopplung unserer bisherigen Einblicke in die Lebensgewohnheiten einer sehr bemerkenswerten Reptiliengruppe“)[8] Da Hautabdrücke von Hadrosauriden zuvor lediglich von wenigen Fragmenten bekannt waren, wurde die von Hautabdrücken eingehüllte Trachodon-Mumie zu einer paläontologischen Sensation.[3] Osborn schrieb jedoch 1912, dass das berühmte Typusexemplar von Trachodon mirabilis (AMNH 5730), das 1884 von Jacob Wortman gefunden wurde, ursprünglich ebenfalls von einer Hülle aus Hautabdrücken umgeben war. Diese Abdrücke wurden jedoch bei der Bergung des Skeletts größtenteils zerstört, weshalb nurmehr drei Fragmente aus der Schwanzregion vorliegen. Wie Osborn weiter anmerkte, ist es sehr schwierig, bei einer Ausgrabung die sehr dünne Schicht aus Hautabdrücken zu entdecken; so seien in den vorangegangenen Jahren möglicherweise viele Funde fossiler Haut verloren gegangen, weil sie weder erwartet noch erkannt worden waren.[7]
Die Trachodon-Mumie wird den Hadrosauridae („Entenschnabeldinosauriern“) zugeordnet, einer Gruppe innerhalb der Ornithischia („Vogelbeckensaurier“). Barnum Brown identifizierte die Mumie anfangs als Trachodon annectens. Der Gattung Trachodon wurden zu dieser Zeit nahezu sämtliche bekannten Hadrosauriden-Funde zugeordnet.[9] Ab 1942 wurde die Mumie unter der neuen Art Anatosaurus copei geführt,[10] welche 1990 in eine eigenständige Gattung, Anatotitan, ausgelagert wurde.[11] Heute wird Anatotitan copei von einem Großteil der Forscher als identisch mit Edmontosaurus annectens angesehen.[12] Den Hadrosauridae werden ein Großteil aller bekannten Dinosaurier-Hautabdrücke zugeschrieben; im Maastrichtium Nordamerikas werden Hadrosauriden-Fossilien 31 Mal häufiger mit Hautabdrücken vorgefunden als Fossilien anderer Gruppen.[13] Die Gründe für diese Verteilung sind unklar. Auf die Gattung Edmontosaurus entfallen 25 % aller bekannten Hautabdrücke von Hadrosauriden.[13]
Beschreibung und Interpretationen
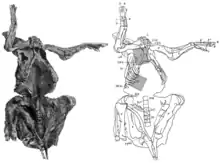
Die Mumie ist zu einem Großteil erhalten und die Knochen befinden sich noch in ihrem ursprünglichen anatomischen Verbund. Es fehlen die Hinterfüße, der hintere Abschnitt des Beckens sowie der Schwanz; diese Teile sind durch Erosion verloren gegangen, noch bevor das Fossil entdeckt wurde. Fossilien werden während der Fossilisierung sehr häufig flachgedrückt; die Trachodon-Mumie ist jedoch dreidimensional erhalten.[14] Das Exemplar liegt auf dem Rücken, Kopf und Hals sind nach unten und zur rechten Seite des Körpers gedreht. Die Knie sind hochgezogen, während die Vorderbeine ausgestreckt nach oben zeigen. Der Brustkorb ist offen.[7] Fast zwei Drittel der gesamten Hautfläche liegen als teilweise exzellent erhaltene Abdrücke vor,[7] was Osborn zu der Bezeichnung „Dinosaurier-Mumie“ verleitete.[8] Die Hautabdrücke sind eng an die Knochen gepresst und zwischen den Knochen in den Körper hineingezogen. Vermutlich war bei seiner Entdeckung das gesamte Skelett von Hautabdrücken umgeben, die jedoch bei der Herausarbeitung des Fossils aus dem Gestein teilweise zerstört wurden.[7] Die Hautabdrücke finden sich an den Vorderbeinen, Hals und Kehle, der Brustregion sowie an der rechten Seite des Rumpfs.[7] Sternberg wies außerdem auf die Erhaltung von Muskelabdrücken hin.[6] Carpenter (2007) gab an, dass möglicherweise auch Abdrücke von inneren Organen vorhanden sind; um diese Möglichkeit zu überprüfen, seien aber detaillierte Computertomographie- und Röntgenanalysen nötig.[14]
Haut

Die Haut erscheint im Verhältnis zur Körpergröße sehr dünn und filigran. Wie für Dinosaurier typisch besteht die Beschuppung aus sich nicht überlappenden Schuppen (Tuberkeln). Die Grundfläche wird von rundlichen „Grundtuberkeln“ mit einem Durchmesser von 1 bis 3 Millimetern gebildet, die von inselartigen Anhäufungen (Clustern) größerer, meist weniger als fünf Millimeter großer polygonaler „Pflastertuberkel“ unterbrochen werden. Die Cluster können aus 20 bis zu mehreren Hundert Pflastertuberkeln bestehen. Sie werden durch intermediäre Tuberkel gesäumt, die in Größe und Form zwischen Grundtuberkel und Pflastertuberkel vermitteln. Im Brust- und Bauchbereich sind die Cluster klein, oval geformt und in unregelmäßigen Längsreihen angeordnet. An den Seiten werden sie größer (5 bis 10 Zentimeter), ihre Form ist unregelmäßiger. Die größten Cluster finden sich oberhalb des Beckens und messen 50 Zentimeter im Durchmesser; vermutlich waren ähnlich große Cluster auf dem gesamten Rücken vorhanden.[7]
Muskeln und Gelenke waren generell mit kleineren Tuberkeln besetzt, um eine größere Flexibilität zu gewährleisten – größere Tuberkel finden sich in den Partien, die eng an die Knochen gepresst sind.[1][7] Die größten erhaltenen Schuppen finden sich auf der Außenseite der Arme; diese polygonalen Tuberkel erreichen einen Durchmesser von einem Zentimeter. Die Innenseite der Arme war dagegen vollständig von kleinen Tuberkeln bedeckt. Der Oberschenkel zeigt ebenfalls relativ kleine Tuberkel auf der Innenseite; von der Außenseite sind keine Abdrücke überliefert.[7] Hautabdrücke vom Schädel finden sich lediglich innerhalb des großen, die Nasenöffnungen beherbergenden Schädelfensters. Hier ist die Haut glatt und in Falten geworfen; Tuberkel fehlen gänzlich. Eine Vertiefung der Haut im vorderen Abschnitt des Schädelfensters könnte die Position der fleischigen Nasenöffnungen markieren.[10]
Hautkamm
Oberhalb der Halswirbelsäule ist ein 25 Zentimeter langer und 7 bis 8 Zentimeter tiefer Hautabdruck erhalten. Osborn interpretierte diesen Bereich als Teil eines ornamentalen Hautkamms, der sich entlang der Mittellinie von Hals und Rücken erstreckte. Oberhalb der Wirbelgelenke sei der Kamm in Falten geschlagen, um die Beweglichkeit des Halses zu gewährleisten, was den Kamm wie eine Krause erscheinen ließe. Zwischen den gefalteten Bereichen des Kamms findet sich jeweils ein großer, ovaler Cluster aus Pflastertuberkeln. Osborn bemerkte, dass der obere Rand des Kamms bei der Bergung der Mumie zerstört worden ist, weshalb sich die Höhe des Kamms nicht mehr feststellen lässt. Vermutlich sei der Kamm nach oben durch mindestens eine weitere Reihe von Clustern erweitert gewesen. Stephen Czerkas (1997) argumentierte, dass der Kamm beim lebenden Tier nicht gefaltet gewesen sei; die Faltung sei eine Folge der Mumifikation und durch vertrocknete Nackenbänder hervorgerufen worden.[15] Ein 1984 von John Horner beschriebenes Edmontosaurus-Fossil zeigt, dass der obere Rand des Hautkamms zumindest im Schwanzbereich in regelmäßigen, rechteckigen Lappen segmentiert war.[16]
Hand
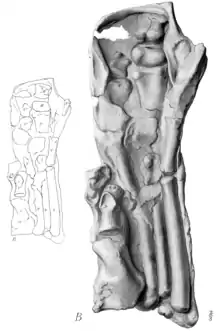
Die Trachodon-Mumie ermöglichte die erste akkurate Rekonstruktion des Handskeletts eines Hadrosauriden. Barnum Brown (1912) zeigte, dass die Handwurzel nicht aus zwei vollständigen Reihen verknöcherter Handwurzelknochen bestand, wie Othniel Charles Marsh in seiner verbreiteten Rekonstruktion von 1892 angenommen hatte, sondern dass lediglich zwei verknöcherte Handwurzelknochen vorhanden waren. Diese Elemente liegen in der Mumie direkt übereinander und oberhalb des dritten Mittelhandknochens. Wahrscheinlich handelt es sich dabei um die ursprüngliche Lebendposition, da die Knochen in beiden Händen in derselben Position erhalten sind.[17][18] Weiter wies Brown darauf hin, dass der erste Finger fehlte und die Finger 2, 3, 4 und 5 aus je drei Fingergliedern bestanden. Marsh hatte den ersten Finger als reduziertes Element mit nur zwei Fingergliedern rekonstruiert, der fünfte Finger war in seiner Rekonstruktion nicht vorhanden.[18]
Die Finger der Trachodon-Mumie sind durch eine Hülle aus Hautabdrücken teilweise miteinander verbunden. Osborn argumentierte 1912, dass es sich bei dieser Hauthülle um Schwimmhäute gehandelt und die Vordergliedmaße als Paddel fungiert habe, was ein deutlicher Hinweis auf eine aquatische Lebensweise von Trachodon und vermutlich anderer Vertreter der Trachodontidae (= Hadrosauridae) sei. Die Schwimmhäute hätten nicht nur die Finger miteinander verbunden, sondern sich bis fünf Zentimeter über die Fingerspitzen hinaus erstreckt. Beachtenswert seien außerdem das Fehlen deutlich ausgeprägter Hufe und großer fleischiger Fußpolster an den Vordergliedmaßen – Merkmale, wie sie bei einem primär landlebenden Tier zu erwarten seien.[7] Mit der Senckenberg-Mumie wurde 1910 ein weiteres Trachodon-Exemplar mit schwimmhautartigen Strukturen entdeckt. Während die Hypothese einer aquatischen Lebensweise zuvor als umstritten und spekulativ galt, etablierte sie sich mit der Entdeckung dieser beiden Mumien zur unbestrittenen Lehrmeinung.[19][20][21] Charles H. Sternberg schrieb 1917: “I was reluctently giving up Marsh's and Cope’s ideas; they believed these dinosaurs lived on land, feeding off the tender foliage of trees […] Entirely different views are held now […] These [the duck-bills] lived in the water instead on land, and consequently they had thin skin and strong paddles, or rather webbed feet.” (deutsch: „Ich hatte Marshs und Copes Ideen nur widerwillig aufgegeben; sie haben geglaubt, dass diese Dinosaurier auf dem Land lebten und sich von dem weichen Laub von Bäumen ernährt haben […] Heute werden komplett andere Meinungen vertreten […] Die Entenschnabeldinosaurier lebten im Wasser anstatt auf dem Land, und hatten konsequenterweise dünne Haut und starke Paddel, oder vielmehr mit Schwimmhäuten versehene Füße.“)[22]
Erst 1964 meldete John H. Ostrom Zweifel an: Dieser Forscher konnte zeigen, dass sich Hadrosauriden nicht wie zuvor angenommen von weichen Wasserpflanzen ernährten, sondern dass ihr elaborierter Kauapparat zur Zerkleinerung widerstandsfähigen Pflanzenmaterials wie Koniferen ausgelegt war. Die Skelettanatomie würde auf eine hochgradig spezialisierte, zweibeinige Fortbewegung an Land hindeuten. Ostrom bemerkte, dass Hadrosauriden keine Hautknochenplatten oder ähnliche Strukturen zur Verteidigung gegen Räuber zeigten, wie sie bei vielen anderen pflanzenfressenden Dinosauriern zu finden sind. Somit dienten die Schwimmhäute möglicherweise dazu, bei Gefahr eine Flucht ins Wasser zu ermöglichen.[21] Robert Bakker (1986) argumentierte schließlich, dass das Tier keine Schwimmhäute besessen habe und die zwischen den Fingern gespannten Hautreste die Überbleibsel eines fleischigen Handpolsters darstellen, das im Zuge der Mumifikation ausgetrocknet und abgeflacht wurde. Ganz ähnliche, von Fußpolstern herrührende Hautstrukturen fänden sich an heutigen mumifizierten Kamelkadavern. Außerdem seien die Finger kurz und könnten kaum voneinander abgespreizt werden, was sie fundamental von den langen, abgespreizten Zehen heutiger paddelnder Tiere wie beispielsweise Enten unterscheide. Bakker resümiert: “Far from being the best, the duckbills must have been the clumsiest and slowest swimmers in all the Dinosauria.” (deutsch: „Die Entenschnabeldinosaurier waren keinesfalls die besten, sondern die schwerfälligsten und langsamsten Schwimmer innerhalb der gesamten Dinosauria.“)[20] Heute gilt die Schwimmhaut-Hypothese als widerlegt.[19][20]
Verknöcherte Sehnen und Mageninhalt

Charles H. Sternberg stellte 1909 fest, dass entlang der Rückenwirbelsäule hunderte bleistiftdicker verknöcherter Sehnen erhalten geblieben sind. Sternberg spekulierte, dass diese Sehnen der Verteidigung dienten und von den Krallen räuberischer Dinosaurier wie dem zeitgenössischen Tyrannosaurus nicht durchdrungen werden konnten.[5] Heute ist bekannt, dass diese Sehnen die Wirbelsäule versteiften, wahrscheinlich um beim Gehen auftretenden Biegekräften auf die Wirbelsäule entgegenzuwirken, und dass auch ein Großteil des in der Trachodon-Mumie nicht erhaltenen Schwanzes durch derartige Sehnen versteift war.[1]
Sternberg berichtete außerdem von verkohlten Nahrungsresten, die in der Magenregion der Trachodon-Mumie entdeckt wurden.[6][23] Eine Analyse dieser Reste wurde bis heute nicht durchgeführt.[24] Kräusel (1922) untersuchte eine Konkretion von braunem Pflanzenmaterial in der Senckenberg-Mumie, die vor allem aus Nadeln und Ästen von Koniferen bestand. Bei beiden Mumien kann allerdings nicht ausgeschlossen werden, dass das Pflanzenmaterial erst nach dem Tod des Tieres in die Bauchhöhle eingewaschen worden ist.[24]
Farbmuster
Osborn beobachtete 1912 dass die Pflastertuberkel-Cluster auf der Oberseite des Rumpfes und der Gliedmaßen deutlich zahlreicher waren als auf der Unterseite. Folglich würden sie in den Bereichen dominieren, die der Sonne ausgesetzt waren; bei vielen heute lebenden Reptilien seien diese sonnenexponierten Bereiche mit den meisten Pigmenten besetzt. Aus diesen Beobachtungen hypothetisierte Osborn einen Zusammenhang zwischen Pigmentierung und Beschuppung: So könnten die Pflastertuberkel-Cluster dunkler gefärbte Bereiche auf einer hellen Grundfläche repräsentiert haben. Die unregelmäßige Verteilung der Cluster würde auf ein unregelmäßiges Farbmuster weisen. Das elaborierteste Farbmuster sei auf dem Hautkamm des Halses zu finden. Allerdings räumte Osborn ein, dass bei heutigen Echsen die Verteilung der Pigmente vollständig unabhängig von der Art der Beschuppung ist.[7] Heutige Forscher stellten fest, dass Aussagen über das Farbmuster nicht möglich sind.[19]
Taphonomie
Verschiedene Autoren beschäftigten sich mit der Frage, wie das Tier zu Tode gekommen ist und welche Umstände zu der außergewöhnlich guten Erhaltung geführt haben. Charles H. Sternberg (1909)[5][6] und Charles M. Sternberg (1970)[25] gingen von einem Tod im Wasser aus. Die sich nach dem Tod in der Bauchregion sammelnden Gase hätten den Kadaver aufschwimmen lassen, wobei der Bauch nach oben zeigte und der Kopf unter der Schulter zu liegen kam. Am Einbettungsort angelangt, sei der Kadaver auf dem Rücken liegend von Sedimenten zugeschüttet worden. Die Haut sei durch die Auflast der Sedimente[25] oder durch das Entweichen der Gase[5] in die Körperhöhle hineingezogen. Osborn schlug 1911 ein anderes Szenario vor: So sei das Tier einen natürlichen Tod gestorben, und der Kadaver hätte für lange Zeit der Sonne ausgesetzt in einem trockengefallenen Flussbett gelegen, unbeeinträchtigt von Aasfressern. Muskeln und Eingeweide seien vollständig ausgetrocknet und damit geschrumpft, wodurch die hart und ledrig gewordene Haut einsank und schließlich eng an den Knochen haftete. Die so entstandene natürliche Mumie sei am Ende der Trockenzeit von einer plötzlichen Flut erfasst, eine Strecke transportiert und am Einbettungsort sehr schnell von Sedimenten eingedeckt worden. Die feine Korngröße des Sediments (feiner Flusssand mit ausreichendem Tonanteil) hätte zu dem perfekten Abdruck der filigranen Hautstrukturen geführt, noch bevor die gehärtete Haut aufweichen konnte.[8] Heute gilt Osborns Hypothese als die wahrscheinlichste.[1][14]

Über die Todesursache des Tieres lässt sich lediglich spekulieren. Kenneth Carpenter (2007) hielt Verhungern während einer Dürre für die wahrscheinlichste Todesursache. So gleiche die Mumie mit ihrer um die Knochen versunkenen Haut Kadavern heutiger Tiere, wie sie während Dürren gefunden werden. Außerdem sei der Kadaver nicht von Aasfressern beeinträchtigt worden. Ein solcher Umstand sei während heutiger Dürren zu beobachten, wenn Trockenheit Aasfresser aus dem betroffenen Gebiet fernhält, oder eine derart große Anzahl von Kadavern anfällt, dass sie von den vorhandenen Aasfressern nicht mehr bewältigt werden kann. Schließlich bemerkt Carpenter, dass heutige große Paarhufer während Dürren eng an Wasser gebunden sind, um Überhitzung zu vermeiden. Die wichtigste Todesursache dieser Tiere während Dürren sei Verhungern, nicht Verdursten. Die Trachodon-Mumie sei in Flusssedimenten entdeckt worden; das Tier müsse folglich in der Nähe eines Flusses oder zumindest eines ausgetrockneten Flussbetts gestorben sein.[14]
Die Sedimente des Fundorts wurden durch einen mäandrierenden Fluss abgelagert. Ein solcher Fluss ändert beständig seinen Lauf, indem er das Ufer an der Außenseite der Flussschlaufen (Prallhang) abträgt und Sediment auf der Innenseite (Gleithang) ablagert. Der Gleithang bildet eine charakteristische Sedimentabfolge aus, die Gleithangsequenz. Carpenter folgert anhand eines während der Grabung gemachten Fotos, dass das Fossil innerhalb einer Gleithangsequenz entdeckt wurde, der Kadaver also auf einem Gleithang eingebettet wurde. Carpenter vermutete, dass der Kadaver während Flutereignissen nach Ende einer Dürre eingebettet wurde. Ein bedeutender Faktor zur schnellen Einbettung liegt laut Carpenter darin begründet, dass der Kadaver als Hindernis in der Strömung auftritt: Beim Umlaufen des Kadavers erhöht sich die Strömungsgeschwindigkeit des Wassers, wodurch Sediment abgetragen wird. Der Kadaver würde in die so geschaffene Mulde sukzessive einsinken. Das für die weitere Einbettung nötige Sediment sei flussaufwärts durch kollabierende Prallhänge in den Fluss gelangt, worauf der hohe Tongehalt der Sandsteine deuten würde. Die Sedimentfracht des Flusses hätte sich insbesondere bei ausklingender Flut abgelagert, wenn die Strömungsgeschwindigkeit und damit das Transportvermögen des Flusses abnimmt. Somit sei der Kadaver innerhalb von Stunden oder höchstens weniger Tage eingebettet worden.[14]
Was nach der Einbettung geschah, ist nur schwer rekonstruierbar, vor allem auch, weil heute keine Proben des Sandsteins mehr existieren, der die Mumie unmittelbar umgab. Wahrscheinlich hat zunächst die langsame bakterielle Verwesung des nun wieder feuchten Kadavers eingesetzt. Carpenter betont, dass die exzellente Erhaltung des Fossils erst durch bakteriell gebildete Minerale möglich wurde (Biomineralisation). Diese Minerale hätten den die Mumie umgebenden tonreichen Sand verfestigt, was die ungewöhnliche dreidimensionale Erhaltung ermöglichte. An der ursprünglichen Kontaktfläche von Sand und Kadaver sei somit zugleich auch die Oberflächenstruktur der Haut als Abdruck erhalten worden. Außerdem habe die Mineralisierung die Knochen in ihrer anatomischen Lage verankert und damit verhindert, dass sie bei fortschreitender Verwesung des Weichgewebes auseinanderfielen. Wichtige seinerzeit gebildete Minerale seien das Kalziumkarbonat Kalzit sowie das Eisenkarbonat Siderit, welches – zu Limonit oxidiert – für die rostige Farbe der Mumie verantwortlich sei. Nach der mehr oder weniger vollständigen Zersetzung des Weichgewebes sei im zementierten Sediment ein Hohlraum zurückgeblieben, der nachfolgend mit Sand verfüllt worden sei. Dabei seien auch die Hautabdrücke an der Innenwand des Hohlraumes ausgefüllt worden.[14] Im Zuge der normalen Gesteinsbildung ist schließlich auch dieser Sand zementiert worden. Somit ist die Weichkörperüberlieferung der Trachodon-Mumie zu einem gewissen Grad mit der Bildung von Steinkernen bei wirbellosen Meerestieren mit Außenskelett vergleichbar.
Literatur
- Kenneth Carpenter: How to Make a Fossil: Part 2 – Dinosaur Mummies and Other Soft Tissue. In: The Journal of Paleontological Science. 2007, S. 5–14.
- Phillip Lars Manning: Grave secrets of dinosaurs: soft tissues and hard science. National Geographic, Washington, D. C. 2008, Chapter four: Dinosaur Mummies.
- Henry Fairfield Osborn: Integument of the iguanodont dinosaur Trachodon. In: Memoirs of the American Museum of Natural History. Band 1, 1912, S. 30–54.
- Henry Fairfield Osborn: A Dinosaur Mummy. In: The American Museum Journal. Band 11. New York 1911, S. 7–11.
- Katherine Rogers: The Sternberg Fossil Hunters: A Dinosaur Dynasty. Mountain Press Publishing Company, 1999, ISBN 0-87842-404-0, S. 106–121.
- Charles H. Sternberg: The Life of a Fossil Hunter. Henry Holt and Company, New York 1909, S. 270–277.
Einzelnachweise
- Phillip Lars Manning: Grave secrets of dinosaurs: soft tissues and hard science. National Geographic, Washington, D. C. 2008, Chapter four: Dinosaur Mummies.
- Mark A. Norell, Eugène S. Gaffney, Lowell Dingus: Discovering Dinosaurs: Evolution, Extinction, and the Lessons of Pre-History. University of California Press, 2000, ISBN 0-520-22501-5, Edmontosaurus annectens, S. 154–155.
- Edwin Harris Colbert: The Great Dinosaur Hunters and Their Discoveries. Courier Dover Publications, 1984, ISBN 0-486-24701-5, S. 184, 195–197.
- Katherine Rogers: The Sternberg Fossil Hunters: A Dinosaur Dynasty. Mountain Press Publishing Company, 1999, ISBN 0-87842-404-0, S. 107–108, 110–115.
- Charles H. Sternberg: The Life of a Fossil Hunter. Henry Holt and Company, New York 1909, S. 270–277.
- Charles H. Sternberg: Expedition to the Laramie Beds of Converse County, Wyoming. In: Transactions of the Kansas Academy of Science. Band 22, 31. Dezember 1909, S. 113–116.
- Henry Fairfield Osborn: Integument of the iguanodont dinosaur Trachodon. In: Memoirs of the American Museum of Natural History. Band 1, 1912, S. 33–35, 46–54.
- Henry Fairfield Osborn: A Dinosaur Mummy. In: The American Museum Journal. Band 11. New York 1911, S. 7–11.
- Benjamin S. Creisler: Deciphering duckbills: a history in nomenclature. In: Kenneth Carpenter (Hrsg.): Horns and Beaks. Ceratopsian and Ornithopod Dinosaurs. Indiana University Press, Bloomington 2007, S. 196, 198.
- Richard Swann Lull, Nelda E. Wright: Hadrosaurian Dinosaurs of North America. In: Geological Society of America Special Papers. Nr. 40, 1942, S. 113.
- Ralph E. Chapman, Michael K. Brett-Surman: Morphometric observations on hadrosaurid ornithopods. In: Kenneth Carpenter, Philip J. Currie (Hrsg.): Dinosaur Systematics: Perspectives and Approaches. Cambridge University Press, Cambridge 1990, ISBN 0-521-43810-1, S. 177.
- John R. Horner, David B. Weishampel, Catherine A. Forster: Hadrosauridae. In: David B. Weishampel, Halszka Osmólska, Peter Dodson (Hrsg.): The Dinosauria. 2. Auflage. University of California Press, Berkeley 2004, ISBN 0-520-24209-2, S. 460.
- Matt Davis: Census of dinosaur skin reveals lithology may not be the most important factor in increased preservation of hadrosaurid skin. In: Acta Paleontologica Polonica. Band 59, Nr. 3, 2014, S. 601–605, doi:10.4202/app.2012.0077.
- Kenneth Carpenter: How to Make a Fossil: Part 2 – Dinosaur Mummies and Other Soft Tissue. In: The Journal of Paleontological Science. 2007, S. 5–14.
- Stephen A. Czerkas: Skin. In: Philip J. Currie, Kevin Padian (Hrsg.): Encyclopedia of Dinosaurs. Academic Press, San Diego, 1997, S. 671–672.
- John R. Horner: A "Segmented" Epidermal Tail Frill in a Species of Hadrosaurian Dinosaur. In: Journal of Paleontology. Band 58, Nr. 1, Januar 1984, S. 270–271.
- Michael K. Brett-Surman, Jonathan R. Wagner: Discussion of Character Analysis of the Appendicular Anatomy in Campanian and Maastrichtian North American Hadrosaurids – Variation and Ontogeny. In: Kenneth Carpenter (Hrsg.): Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Indiana University Press, Bloomington, Indiana 2007, ISBN 0-253-34817-X, S. 143–144.
- Barnum Brown: The osteology of the manus in the family Trachodontidae. In: Bulletin of the American Museum of Natural History. Band 31, 1912, S. 105–108.
- Catherine A. Forster: Hadrosauridae. In: Philip J. Currie, Kevin Padian (Hrsg.): Encyclopedia of Dinosaurs. Academic Press, San Diego, 1997, S. 294, 297.
- Robert T. Bakker: The dinosaur heresies. William Morrow and Company, Inc., New York 1986, ISBN 0-688-04287-2, S. 146–159.
- John H. Ostrom: A reconsideration of the paleoecology of hadrosaurian dinosaurs. In: American Journal of Science. Band 262, Oktober 1964, S. 975–977, 995–996.
- Charles H. Sternberg: Hunting dinosaurs in the bad lands of the Red Deer River, Alberta, Canada: a sequel to The life of a fossil hunter. The world company press, Lawrence, Kansas 1917, S. 7–8.
- Charles H. Sternberg: A New Trachodon from the Laramie Beds of Converse County, Wyo. In: Science. Band 29, Nr. 749, 1909, S. 753–754.
- Philip J. Currie, Eva B. Koppelhus, A. Fazal Muhammad: "Stomach" contents of a Hadrosaur from the Dinosaur Park Formation (Campanian, Upper Cretaceous) of Alberta, Canada. In: Ailing Sun, Yuanqing Wang (Hrsg.): Sixth Symposium on Mesozoic Terrestrial Ecosystems and Biota, Short Papers. China Ocean Press, Peking 1995, S. 111–114.
- Charles M. Sternberg: Comments on Dinosaurian Preservation in the Cretaceous of Alberta and Wyoming. In: Publications in Palaeontology. Nr. 4. Ottawa 1970, S. 3–4.