Kimberella
Kimberella ist eine ausgestorbene Tiergattung unsicherer systematischer Stellung, die vor ca. 558 bis 555 Millionen Jahren im Ediacarium lebte und der Ediacara-Fauna angehört. Bisher ist nur die Art K. quadrata in die Gattung gestellt worden.
| Kimberella | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Fossil von Kimberella | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Ediacarium | ||||||||||||
| 558 bis 555 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Kimberella | ||||||||||||
| Wade, 1972 | ||||||||||||
Die ersten Fossilien von Kimberella wurden 1959 in den Ediacara-Hügeln von Australien, der namensgebenden Lokalität des Ediacariums (635 bis 542 mya) gefunden. Eine genauere radiometrische Datierung des Stratum typicum für eine absolut-zeitliche Einordnung von Kimberella war nicht möglich. Doch 1997 wurden neuere und wesentlich besser erhaltene Fossilien in der Gegend des Weißen Meeres in Russland gefunden und 2007 gelang eine radiometrische Datierung der Fundschichten auf 558 bis 555 mya.[1]
Ursprünglich wurde Kimberella als Medusenstadium (Qualle) eines Hohltiers klassifiziert. Als 1997 die Bilateralsymmetrie und Spuren einer möglichen Raspelzunge (Radula) entdeckt wurden, erfolgte eine Einordnung als basaler (stammesgeschichtlich ursprünglicher) Vertreter der Weichtiere. Allerdings ist die Interpretation der Reste als Raspelzunge und damit die Einordnung an die Basis der Weichtiere umstritten geblieben. Einig sind sich die Paläontologen aber weiterhin in der Klassifikation von Kimberella als Angehöriger der Bilateria (Zweiseitentiere).
Verbreitung
Kimberella ist bisher nur von zwei Fundstellen bekannt: von den australischen Ediacara-Hügeln[2] und von der Ust’ Pinega-Formation am Weißen Meer in der Nähe der Mündung der Onega.
Die Kimberella-Fossilien vom Weißen Meer werden häufig zusammen mit Algen und den Ediacara-Fossilien Tribrachidium und Dickinsonia gefunden – mäandrierende Spurenfossilien, die evtl. von Kimberella selbst erzeugt wurden. Eine Schicht vulkanischer Asche, die zwischen Schichten mit Kimberella-Fossilien liegt, konnte radiometrisch anhand des Zerfalls von Uran in Zirkonen auf ein Alter von 558 bis 555,3 Millionen Jahre datiert werden.[3] Es sind Kimberella-Fossilien aus anderen Schichten bekannt, die jünger und älter als diese mithilfe der Ascheschicht sehr genau datierten Fossilien sind.[1] Die Fossilien von den Ediacara-Hügeln in Australien konnten im Gegensatz zu den Funden aus Russland bisher nicht exakt datiert werden.
Beschreibung
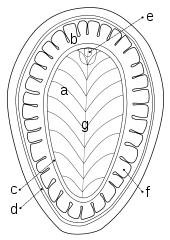
a) Riefen
b) Gekräuselter Saum
c) Innere Kante
d) Äußere Kante
e) Vorderer Hügel
f) Lobus
g) Mittlere Vertiefung oder Erhöhung
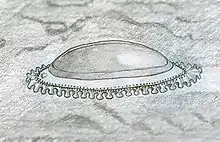
Bisher wurden über 1000 Kimberella-Exemplare aus allen Alters- bzw. Größengruppen in den feinkörnigen Sandsteinschichten der Ust'-Pinega-Formation am Weißen Meer gefunden.[1][4] Die große Anzahl der Fossilien und der gute Erhaltungszustand in dem feinkörnigen Sandstein ließen fundierte Rückschlüsse auf die äußere Morphologie, die interne Anatomie, den Bewegungsapparat und die Lebensweise dieser Gattung zu.[1]
Alle Größenstadien von Kimberella haben eine ovale bis birnenförmige Grundform, wobei größere Individuen eher länglich-oval bzw. länglich-birnenförmig sind. Die kleinsten gefundenen Fossilien sind nur zwei bis drei Millimeter lang. Das größte Individuum ist unvollständig und dürfte rekonstruiert eine Länge von 13 bis 15 cm erreicht haben.[1] In der statistischen Größenverteilung sind keine Maxima zu beobachten, die auf einen Geschlechtsdimorphismus oder eine genetisch fixierte Adultgröße hindeuten könnte. Auch sind keine ontogenetischen Änderungen in der Population zu beobachten; die kleinsten Exemplare sind einfach kleine Ausgaben der größten Exemplare. Lediglich der Wert für das Längen/Breiten-Verhältnis liegt bei größeren Exemplaren etwas höher.
Die Exemplare waren achsensymmetrisch (bilateral) und besaßen rückenseitig (dorsal) eine einzelne Schalenplatte, welche sich über die gesamte Körperlänge erstreckte. Bei größeren Exemplaren konnte diese rückenseitige Schalenplatte bis zu 15 Zentimeter lang, fünf bis sieben Zentimeter breit und drei bis vier Zentimeter dick werden.[1] Die Schalenplatte war zwar fest, aber trotzdem noch flexibel und scheint aus keinem mineralisierten Material bestanden zu haben. Da sie mit steigendem Alter vermutlich immer dicker wurde, wurde sie auch immer steifer.[1] Am einen Ende der Schalenplatte hatte diese eine höckerförmige Struktur, die vermutlich das vordere Ende des Tieres kennzeichnete.[1][5] Manche Exemplare zeigen feine, streifenförmige Vertiefungen über die gesamte Länge der Schalenplatte, die vertikal zur Körperachse stehen. Sie werden als Ansatzstellen für die Muskulatur interpretiert. Ähnliche streifenförmige Vertiefungen wurden auch an den Enden einiger Schalenplatten entdeckt. Hier könnten die Rückziehmuskeln (Retraktormuskeln) für das Einziehen des Fußes angesetzt haben.[1]
Der Organismus hatte einen erhöhten „Grat“ längs der Körperachse und seitlich (lateral) keine glatte, sondern eine leicht gewellte Oberfläche. Kimberella zeigt zwar keine sichtbare Segmentierung, aber trotzdem eine Art Modularisierung. Jedes Modul beinhaltete ein gut ausgebildetes Muskelband, die vertikal zur Körperachse von der höchsten Stelle (Schalenplatte) bis zum muskulösen Fuß angelegt waren. Ebenso kleinere im Fuß befindliche Muskeln, die ebenfalls vertikal zur Körperachse von vorne bis zum hinteren Teil des Körpers angelegt waren. Die Kombinierung der Bewegung der beiden Muskeltypen ermöglichte Kimberella zu kriechen, in dem es den Fuß wellenförmig bewegte.[1][5]
Der Körper hatte einen gekräuselten Randsaum. Es wird vermutet, dass dieser ein Teil des Atmungsapparats gewesen sein könnte und ähnlich funktionierte wie heutige Kiemen. Die Tatsache, dass dieser Saum sehr stark ausgedehnt war, lässt vermuten, dass es sich entweder um ein nicht sehr effizientes Atmungssystem handelte oder dass er doch eine andere Funktion hatte.[1]
Lebensweise

Die Fazies der Ust' Pinega-Formation und die Einbettung der Kimberella-Fossilien aus der Gegend des Weißen Meeres lassen schließen, dass die Tiere auf dem Meeresgrund lebten.[5] Kimberella lebte in seichtem bis etwa zehn Meter tiefen Wasser, vor allem in seichten Rinnen auf schlammigen Untergrund und teilte den Lebensraum mit photosynthetisch aktiven Organismen.[1] Kimberella wurde zusammen mit anderen Vertretern der Ediacara-Fauna wie Yorgia, Dickinsonia, Tribrachidium und Charniodiscus gefunden. Dies legt nahe, dass diese Formen im gleichen Lebensraum gelebt haben.[1] Kimberella graste vermutlich mikrobakterielle Matten ab, ohne sich auf eine spezielle Nahrungsquelle spezialisiert zu haben.[1] Während des Fressens kroch sie vermutlich rückwärts und graste die Mikrobenmatten flächig ab, wie die erhaltenen Weidespuren zeigen.[1] Wie sich Kimberella fortpflanzte, ist nicht bekannt.[1]
Die Sedimentoberfläche, auf der Kimberella lebte, wurde gelegentlich durch feinkörnigen Sand verschüttet, z. B. durch Sturmereignisse oder die Änderung der Strömung. Als Antwort auf diesen Stress zogen sich die Tiere ähnlich der Reaktion vieler Weichtiere unter ihre Schalenplatte zurück. Es ist eher unwahrscheinlich, dass sie schnell genug waren, um einem solchen Ereignis zu entkommen.[1] Manche Exemplare überlebten die Verschüttung und konnten sich selbständig aus dem Sand ausgraben wie Fluchtspuren zeigen. Vor allem aber jüngere Tiere blieben trotz Fluchtversuchen im Sediment gefangen und wurden so am Ende eines wenige Zentimeter langen Tunnels fossilisiert.[1]
Erhaltung
Die Erhaltung der Kimberella-Fossilien wurde einerseits durch die schnelle Sedimentation ermöglicht, welche die toten Körper rasch vom Kontakt mit dem Meerwasser und auch Aasfressern abschnitt, andererseits auch eine rasche Verfestigung (Lithifizierung) der darunter und darüber liegenden Sedimente.[1] Die Lithifizierung der Sedimente wurde möglicherweise durch die Verwesungsprodukte anderer Organismen beschleunigt. Einige Wissenschaftler nehmen sogar an, dass auch der Schleim des Tieres seine eigene Fossilierung mitbeeinflusst haben könnte.[1] Dagegen spricht, dass sich der Schleim wahrscheinlich sehr schnell verflüchtigte und so keine Rolle bei der Fossilisierung spielen konnte.[6] Die rasche Verfestigung der Sedimente führte dazu, dass die dreidimensionale Form des Tieres bei der Fossilisierung erhalten geblieben ist.[1]
Systematik
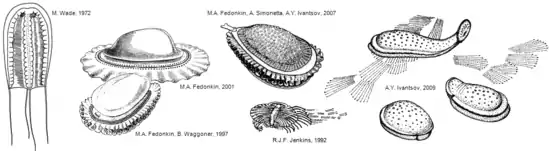
nach Mikhail A. Fedonkin 1997, 2001
Das erste Kimberella-Fossil wurde 1959 in Australien entdeckt und 1966 von Martin Glaessner und Mary Wade als Kimberia erstmals wissenschaftlich beschrieben und benannt. Der Name ehrt John Kimber, einen Lehrer und Sammler, der 1964 bei einer Expedition ins Zentrum Australiens ums Leben kam. Ursprünglich wurde das Fossil als Medusenstadium eines Hohltiers klassifiziert.[7] 1972 benannte Mary Wade die Gattung in Kimberella um, da der Name Kimberia bereits vergeben war. Kimberella wurde nun als Würfelqualle (Cubozoa) interpretiert. Diese systematische Einordnung blieb solange bestehen, bis die Fossilien in der Region des Weißen Meeres gefunden wurden.[8] Mikhail A. Fedonkin und Benjamin M. Waggoner beschrieben 1997 Kimberella auf der Basis dieses umfangreichen Materials neu. Sie stellten zweifelsfrei fest, dass Kimberella bilateralsymmetrisch war. Die Systematisierung als Medusenstadium (Qualle) eines Nesseltiers oder Rippenqualle war damit widerlegt, da zum Grundplan dieser Gruppen ein radialsymmetrischer Bauplan gehört. Fedonkin und Waggoner beobachteten eine dorsale, nicht mineralisierte, flexible Schalenplatte, unter die sich das Tier möglicherweise zurückziehen konnte. Die Spur, die ein Exemplar von Kimberella auf der Sedimentoberfläche hinterlassen hat, kurz bevor es verschüttet wurde, weist Kratzspuren auf. Diese Kratzspuren wurden von diesen Autoren als Weidespuren interpretiert, die entstanden, als das Exemplar den mikrobakteriellen Rasen auf der Sedimentoberfläche mit einer Raspelzunge (Radula) abweidete. Kimberella wird deshalb von diesen Autoren als einschaliges Weichtier oder als ein sehr nahe verwandtes Tier betrachtet.[5] Bei den Fossilien von Kimberella wurde aber die typische Raspelzunge eines modernen Weichtiers bisher nicht nachgewiesen. Allerdings hat die Raspelzunge (Radula) der Weichtiere aufgrund ihres ausschließlich organischen Aufbaus ein sehr geringes Fossilisationspotenzial, und bisher sind nur sehr wenige fossilisierte Radulae überhaupt gefunden worden (z. B. Ammoniten, aber bisher noch keine Schneckenradulae!). Deshalb ist dieser negative Befund kein Argument, dass Kimberella keine Raspelzunge gehabt hat.
Die Interpretation von Kimberella als Weichtier ist nicht unwidersprochen geblieben. Kritiker halten dagegen, dass die wenigen verfügbaren Fakten über Kimberella es nicht erlauben, sie systematisch zu den Weichtieren oder mit diesen verwandten Tieren zu stellen. Die Kratzspuren könnten auch von anderen Körperstrukturen verursacht worden sein. Man müsse sogar die Einordnung als Zweiseitentier in Zweifel ziehen (vgl. Diskussion in Butterfield[9]).
Heute gilt als nahezu gesichert, dass Kimberella, möglicherweise nach Vernanimalcula,[10] das älteste je dokumentierte triploblastische Zweiseitentier ist.[11] Die Interpretation als Weichtier ist dagegen weiter mit Zweifeln behaftet.
Bedeutung für die Stammesgeschichte der Tiere
Die systematische Stellung von Kimberella ist wichtig für das Verständnis der Kambrischen Radiation. Die Theorie der Kambrischen Explosion geht davon aus, dass die grundlegenden Baupläne der heutigen Arten zwischen 543 Mio. Jahre und 518 Mio. Jahre entstanden sind.[12] Die Interpretation von Kimberella als ein Zweiseitentier würde damit die Diversifikation der Tierüberstämme bereits in das Präkambrium verschieben. Sollte es sich bei Kimberella tatsächlich um ein Weichtier und somit um ein Urmundtier handeln, dann hätten sich nicht nur die Tierüberstämme der Urmünder (Protostomia) und die Neumünder (Deuterostomia) bereits einige Zeit vor 555 Millionen Jahre und damit dem Beginn des Kambriums (Grenze Ediacarium/Kambrium: 542 Millionen Jahre) evolutionär auseinanderentwickelt, sondern auch die Tierstämme der Weichtiere und der Ringelwürmer/Gliederfüßer, die derzeit als Schwestergruppen interpretiert werden. Die Existenz des einen Tierstamms (Weichtiere) beweist damit auch die Existenz der anderen Schwestergruppe (Ringelwürmer/Gliederfüßer), auch wenn bisher noch keine sicheren Fossilien dieses anderen Tierstammes gefunden wurden (terminus post quem non).
Einzelnachweise
- M. A. Fedonkin, A. Simonetta, A. Y. Ivantsov: New data on Kimberella, the Vendian mollusc-like organism (White sea region, Russia): palaeoecological and evolutionary implications. In: Patricia Vickers-Rich, Patricia Komarower: The Rise and Fall of the Ediacaran Biota. (= Special publications. 286). Geological Society, London 2007, ISBN 978-1-86239-233-5, S. 157–179. doi:10.1144/SP286.12
- M. Glaessner, B. Daily: The Geology and Late Precambrian Fauna of the Ediacara Fossil Reserve. (PDF; 69,4 MB). In: Records of the South Australian Museum. 13, 1959, S. 369–401.
- M. W. Martin, D. V. Grazhdankin, S. A. Bowring, D. A. D. Evans, M. A. Fedonkin, J. L. Kirschvink: Age of Neoproterozoic Bilaterian Body and Trace Fossils, White Sea, Russia: Implications for Metazoan Evolution. In: Science. Band 288, Nr. 5467, 5. Mai 2000, S. 841. doi:10.1126/science.288.5467.841. PMID 10797002. Abgerufen 10. Mai 2007.
- A. Yu. Ivantsov: New reconstruction of Kimberella, problematic Vendian metazoan. In: Paleontological Journal. Band 43, Nr. 6, 2009, S. 601–611. doi:10.1134/S003103010906001X.
- M. A. Fedonkin, B. M. Waggoner: The Late Precambrian fossil Kimberella is a mollusc-like bilaterian organism. In: Nature. Band 388, Nr. 6645, 1997, S. 388. doi:10.1038/42242
- P. R. Getty: Producing And Preserving Climactichnites. Philadelphia Annual Meeting, 2006. (online), abgerufen am 2. Juni 2008.
- M. F. Glaessner, M. Wade: The late Precambrian fossils from Ediacara, South Australia. In: Palaeontology. Band 9, Nr. 4, 1966, S. 599 (palass.org [PDF]).
- M. Wade: Hydrozoa and Scyphozoa and other medusoids from the Precambrian Ediacara fauna, South Australia. In: Palaeontology. Ausgabe 15, 1972, S. 197–225.
- N. J. Butterfield: Hooking some stem-group "worms": fossil lophotrochozoans in the Burgess Shale. In: Bioessays. Band 28, Nr. 12, 2006, S. 1161–1166. doi:10.1002/bies.20507
- J. Y. Chen, D. J. Bottjer, P. Oliveri, S. Q. Dornbos, F. Gao, S. Ruffins, H. Chi, C. W. Li, E. H. Davidson: Small bilaterian fossils from 40 to 55 million years before the Cambrian. In: Science. 305, 2004, S. 218–222.
- Douglas H. Erwin, Eric H. Davidson: The last common bilaterian ancestor. In: Development. Ausgabe 129, 2002, S. 3021–3032. (online)
- R. Cowen: History of Life. 3. Auflage. Blackwell Science, 2000, ISBN 0-632-04444-6, S. 63.