Gap Junction
Gap Junctions sind Ansammlungen („plaques“ oder „cluster“) von Zell-Zell-Kanälen, die die Zellmembranen zweier benachbarter Zellen durchqueren. Sie verbinden das Cytoplasma benachbarter Zellen direkt miteinander. Dabei werden die Membranen der Zellen in einem Abstand von nur 2 bis 4 Nanometer zueinander fixiert, lassen zwischen sich aber eine elektronenmikroskopisch erkennbare Lücke (engl.: gap) frei (im Gegensatz zu den Tight Junctions). Die Kanäle der Gap Junction werden aus zwei Halbkanälen (Hemichannels, Connexone) gebildet, von denen jede Zelle jeweils einen beisteuert. Jedes Connexon besteht aus Proteinkomplexen aus (in der Regel) sechs membrandurchspannenden Proteinen, die sich in einer sechseckigen Anordnung so zusammenlagern, dass in der Mitte zwischen ihnen eine Pore frei bleibt. Zwei solche Proteinkomplexe der benachbarten Zellmembranen zweier Zellen lagern sich schließlich so zusammen, dass sie gemeinsam einen Kanal bilden.
| Übergeordnet |
| Zell-Zell-Kontakt |
| Untergeordnet |
| Connexon Pannexon |
| Gene Ontology |
|---|
| QuickGO |
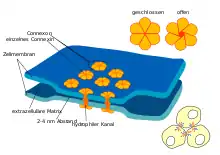
Aufgabe der Gap Junctions ist die Kommunikation (Austausch von Signalen) zwischen benachbarten Zellen. Dabei werden Ionen oder kleine Moleküle durch Diffusion direkt von einer Zelle auf die Nachbarzelle übertragen. Dies können z. B. als Signalgeber dienende Kalium- oder Calcium-Ionen, sekundäre Botenstoffe (Second Messenger) wie cAMP, cGMP oder IP3, oder kleine Stoffwechselprodukte (Metabolite) wie Glucose sein. Je nach dem speziellen Aufbau des Zellkanals ist der Übergang mehr oder weniger selektiv, er kann durch Signalgeber wie z. B. das Membranpotential gesteuert werden. Einige Typen von Kanälen leiten auch bestimmte Stoffe selektiv nur in eine Richtung weiter.
Die Kanäle der Gap Junctions werden bei verschiedenen Tierarten von zwei verschiedenen Proteinfamilien aufgebaut. Ausschließlich bei den Chordatieren (Wirbeltiere und Verwandte) bestehen sie aus Connexinen genannten Proteinen. Bei allen anderen Gewebetieren erfüllt diese Aufgabe eine andere Proteinfamilie, die Innexine genannt werden. Beide sind funktional ähnlich aufgebaut, besitzen aber keine homologen Basensequenzen, sind also nicht nahe miteinander verwandt. Inzwischen wurde bei den Wirbeltieren eine Proteinfamilie neu entdeckt, die Pannexine, die in ihrer Sequenz größere Ähnlichkeit mit den Innexinen aufweisen, also vermutlich homolog zu ihnen sind. Allerdings sind Pannexine äußerstenfalls sehr selten an der Bildung von Gap Junctions beteiligt (wahrscheinlich sogar überhaupt nicht), sie erfüllen andere Aufgaben in der Zelle. Weitere verwandte, membrandurchspannende Proteine sind die an der Bildung der Tight Junctions beteiligten Claudine und Occludin.
Bei Pflanzen erfüllen die Plasmodesmata ähnliche Aufgaben, unterscheiden sich vom Aufbau jedoch deutlich von den aus Innexinen und Connexinen aufgebauten Kanälen.
Forschungsgeschichte
Die ersten Hinweise auf die Zellkopplung stammten von elektrophysiologischen Untersuchungen an spezifischen Paaren interagierender Nervenzellen im Rückenmark des Flusskrebses. Die Messung der elektrischen Kopplung dieser benachbarten Riesenneuronen wurde zuerst von Fursphan und Potter 1959 durchgeführt (Fursphan und Potter 1959).
Geprägt wurde der Begriff der Gap Junctions durch Jean-Paul Revel und Morris Karnovsky 1967, die als erste zeigten, dass sich im elektronenmikroskopischen Bild der Abstand zwischen zwei benachbarten Plasmamembranen im Bereich der Gap Junctions von 20–30 nm auf 2–4 nm verengt, wodurch der optische Eindruck einer Lücke (engl.: gap) in der Kontinuität benachbarter Plasmamembranen entsteht.
Aufbau


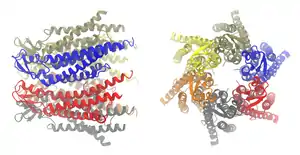
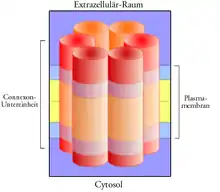
Sechs Connexine (mit je vier Transmembran-Regionen) lagern sich zum sogenannten Connexon (oder Halbkanal) zusammen. Ein Connexon kann homomer (Zusammensetzung aus einer Art von Connexinen) oder heteromer (aus verschiedenen Connexinen) aufgebaut werden. Je nach Zusammensetzung der Connexone kann die Permeabilität des Kanals variieren.
Je ein Halbkanal verbindet sich mit einem ihm gegenüberliegenden Halbkanal der Nachbarzelle zu einer durchgehenden Pore (interzellulärer Kanal, Gap Junction). Der interzelluläre Kanal kann homotypisch (aus zwei gleichen Connexonen) oder heterotypisch (aus unterschiedlichen Connexonen) aufgebaut werden, wobei nicht alle Connexone gleich gut zusammenlagern. Die Pore hat einen Durchmesser von 1,5 bis 2 nm und lässt deshalb Moleküle oder Ionen von maximal ≈1000 Dalton relativer Molekülmasse passieren. Der Aufbau einer Gap Junction (früher: Nexus) kann innerhalb weniger Sekunden erfolgen, wenn zwei Zellen miteinander in Kontakt treten.
Eine Connexon-Untereinheit hat einen Durchmesser von 2,5 nm und ist 7,5 nm lang. Sie ragt 0,7 nm in das Zytosol und 1,7 nm in den Extrazellularraum.
Die Connexone sind in der Biomembran in Feldern in einem regelmäßigen hexagonalen Muster (Abstand der Kanal-Mittelpunkte 8,5 nm) mit einer Dichte von einigen wenigen bis zu 28.000 Kanälen pro Quadratmikrometer angeordnet, sie bilden sogenannte Plaques.
Gap Junctions unterscheiden sich von anderen Kanalsystemen der Zelle:
- Sie durchziehen zwei benachbarte Membranen (statt nur einer).
- Sie verbinden Zytosol mit Zytosol (statt Zytosol mit Extrazellulärraum oder Organellinnenraum).
- Die Connexine werden von zwei verschiedenen Zellen synthetisiert (statt nur von einer).
- In der Regel sind sie im Ruhezustand geöffnet und schließen nur, wenn bestimmte Bedingungen eintreten (siehe weiter unten)
Vorkommen und Funktion
Gap Junctions treten erst bei den Eumetazoa auf. Während im Embryonalstadium Gap Junctions weit verbreitet sind, kommen sie beim Adulten vor allem im Herzmuskel, in Epithel- und Gliazellen sowie in der Retina vor.
Die allgemeinen Funktionen der GJs sind
- direkte elektrische Kommunikation zwischen Zellen (wobei verschiedene Connexine verschiedene Leitfähigkeiten bedingen)
- direkte chemische Kommunikation zwischen Zellen über second messenger (z. Bsp. IP3, Ca2+) (wobei verschiedene Connexine unterschiedliche Selektivität für kleine Moleküle aufweisen)
- Austausch von Molekülen bis ≈1 kDa zwischen Zellen (wobei verschiedene Connexine GJs mit unterschiedlichen Durchmessern bilden können und unterschiedliche Präferenzen für geladene Teilchen besitzen)
- verhindern, dass Moleküle oder Ladungen beim Austausch in den Extrazellularraum verloren gehen.
Beispiele für die Funktion der GJs:
- In schwach durchbluteten Geweben (zum Beispiel Augenlinse und Knochen) dienen sie dem Nahtransport von Nährstoffen: Rand-Zellen nehmen die Nährstoffe auf und geben sie über die Gap Junctions an ihre Nachbarzellen bis ins unterversorgte Zentrum weiter.
- In Drüsen wie Leber und Bauchspeicheldrüse helfen sie bei der Sekretion.
- Im Herzmuskel und im Nervensystem sind sie an der schnellen Weiterleitung von Aktionspotentialen beteiligt.
- Gap Junctions scheinen auch an der Kontrolle des Zellwachstums beteiligt zu sein (zum Beispiel während der Embryonalentwicklung).
- Eines der Connexin-Gene wurde als Tumorsuppressor-Gen identifiziert.
Die Poren durch eine Gap Junction können sehr schnell geschlossen werden, wenn bestimmte Faktoren eintreten, die auf eine Schädigung der benachbarten Zelle hindeuten. Dadurch wird die geschädigte Zelle von ihren Nachbarn abgekoppelt, sodass die gesunden Nachbarzellen in ihrer Zellchemie unbeeinflusst bleiben. Das Schließen wird durch eine hohe cytosolische Kalziumionenkonzentration oder einen niedrigen cytosolischen pH-Wert (also bei hoher Protonenkonzentration) ausgelöst. Beides sind Zeichen für baldigen Zelltod der Nachbarzelle.
Seit einigen Jahren ist bekannt, dass durch alle drei Proteinfamilien (Connexine, Innexine und Pannexine) gebildete Halbkanäle sich nicht immer zu Gap Junctions zusammenschließen müssen. In zahlreichen Zellen und Zelltypen erfüllen sie zusätzliche Funktionen als einfache Membrankanäle ohne Koppelung an andere Zellen. Wahrscheinlich kommen die Pannexine ausschließlich in dieser Form vor.
Gap Junctions als elektrische Synapsen
Gap Junctions fungieren in Neuronen, in der Retina und im Herzen, aber auch bei Invertebraten als spannungsgesteuerte, transmitterfreie Synapsen. Sie werden auch als Elektrische Synapsen bezeichnet. Sie ermöglichen eine schnelle und synchrone Ausbreitung von Aktionspotentialen. In den Glanzstreifen zwischen den Herzmuskelzellen können sie eine Fläche bis zu einem Quadratmikrometer bedecken. Die Leitfähigkeit der Gap Junctions variiert mit der Zusammensetzung aus verschiedenen Connexinen. An Neuronen treten sie nicht so zahlreich auf wie chemische Synapsen, sie wurden aber auch bei Gliazellen gefunden, deren Beteiligung am neuronalen Geschehen über die Versorgung der Nervenzellen hinaus gerade erforscht wird. Die Hauptaufgabe der elektrischen Synapsen scheint die Synchronisierung von Nervenzellgruppen zu sein, die als Oszillatoren und Rhythmusgeber dienen. Möglicherweise spielen sie auch bei epileptischen Anfällen eine Rolle.
Funktionsweise der elektrischen Synapse
Die Depolarisation der präsynaptischen Zelle führt zu einem Potentialgefälle zwischen beiden durch Gap Junctions verbundenen Zellen, so dass Kationen von der präsynaptischen Zelle in Richtung postsynaptische Zelle fließen und Anionen von der post- zur präsynaptischen Zelle.
Wird der Schwellenwert an der postsynaptischen Membran überschritten, folgt hier ein Aktionspotential, und das Signal kann praktisch ohne Zeitverzögerung (10−5 s) weitergeleitet werden (macht Synchronisation vieler Zellen z. B. im Herzmuskel aufgrund geringer Zeitverzögerung möglich!).
Vergleich zwischen elektrischer und chemischer Synapse
Neben der viel geringeren Zeitverzögerung unterscheiden sich elektrische Synapsen von chemischen Synapsen auch darin, dass bei diesen die Erregungsübertragung i. d. R. in beide Richtungen erfolgen kann.
Allerdings können auch Gap Junctions einiger Zellen in ihrer Stromrichtung reguliert werden, entweder von Ca2+ abhängig oder Membranpotential-abhängig. Allerdings hat die Anwendung von Gap Junctions im Körper auch einige Nachteile: Eine direkte Erregungsübertragung auf weit entfernte Zellen ist nicht möglich, vor allem aber kann eine Erregung nicht zur Hemmung einer anderen Zelle genutzt werden. Elektrische Synapsen haben im ZNS der Säugetiere geringere Bedeutung im Vergleich zu chemischen Synapsen.
| elektrische Synapse | chemische Synapse mit direkter Signalübertragung | chemische Synapse mit indirekter Signalübertragung | |
|---|---|---|---|
| Spaltbreite | 3,5 nm | 30 bis 50 nm | |
| Kanalproteine | Gap Junction-Kanäle in prä- und postsynaptischer Membran | postsynaptische, transmittergesteuerte Ionenkanäle | Rezeptoren des Second-Messenger-Systems (G-Proteine) |
| Synaptische Verzögerung | keine | 0,1 bis 0,5 ms | mehr als 10 ms |
| Transmitter | — | Acetylcholin, Gamma-Aminobuttersäure (GABA), Glycin, Glutamat, Aspartat | Noradrenalin, Dopamin, Serotonin, Acetylcholin, Neuropeptide |
| Vorkommen | Herzmuskel, ZNS, auch bei Wirbellosen | Motorische Endplatten, zentrales und peripheres Nervensystem | zentrales und peripheres Nervensystem |
Quellen
- Revel J.P., Karnovsky M.J., Hexagonal array of subunits in intercellular junctions of the mouse heart and liver, J. Cell Biol. 1967; 33: C7-C12.
- Daisuke Fushiki, Yasuo Hamada, Ryoichi Yoshimura, Yasuhisa Endo (2010): Phylogenetic and bioinformatic analysis of gap junction-related proteins, innexins, pannexins and connexins. In: Biomedical Research Vol. 31 No. 2: 133-142. doi:10.2220/biomedres.31.133
- Daniel A. Goodenough & David L. Paul (2009): Gap Junctions. Cold Spring Harbour Perspectives in Biology 2009;1:a002576 doi:10.1101/cshperspect.a002576
- Gülistan Mese, Gabriele Richard, Thomas W. White (2007): Gap Junctions: Basic Structure and Function. In: Journal of Investigative Dermatology Volume 127: 2516-2524. doi:10.1038/sj.jid.5700770
- Silvia Penuela, Ruchi Gehi, Dale W. Laird (2013): The biochemistry and function of pannexin channels. In: Biochimica et Biophysica Acta 1828: 15–22. doi:10.1016/j.bbamem.2012.01.017
- Alberto E. Pereda, Sebastian Curti, Gregory Hoge, Roger Cachope, Carmen E. Flores, John E. Rash (2013): Gap junction-mediated electrical transmission: Regulatory mechanisms and plasticity. In: Biochimica et Biophysica Acta Biomembranes Volume 1828, Issue 1: 134–146 doi:10.1016/j.bbamem.2012.05.026
- Eliana Scemes, David C. Spray, Paolo Meda (2009): Connexins, pannexins, innexins: novel roles of “hemi-channels”. In: Pflügers Arch. 457 (6): 1207–1226. doi:10.1007/s00424-008-0591-5