Myelinscheide
Die Myelinscheide (auch Markscheide, zu altgriechisch μυελός myelos, deutsch ‚Mark‘[1]) ist eine lipidreiche Schicht, die die Axone mancher Nervenzellen von Wirbeltieren umgibt. Sie besteht aus Myelin, das von Gliazellen (Oligodendrozyten im ZNS, Schwann-Zellen im PNS) gebildet wird und die Axone spiralförmig umwickelt. Die Myelinscheide dient der Senkung von Membranleitwert und Membrankapazität und ermöglicht damit die besonders schnelle saltatorische Erregungsleitung.
| Aufbau einer Nervenzelle |
|---|
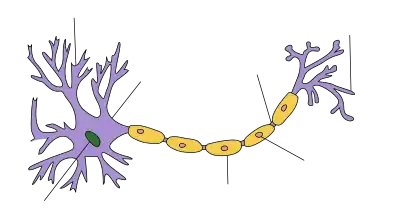
Bildung
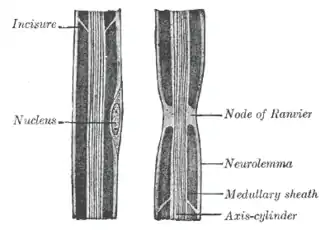
Im ZNS gehen vom Soma eines Oligodendrozyten Fortsätze aus, die jeweils einen Teilabschnitt eines Axons umhüllen. Ein Oligodendrozyt kann so an den Umhüllungen von etwa 10–50 Axonen beteiligt sein.[2] Eine Schwann-Zelle im PNS umwickelt je einen Teilabschnitt eines Axons, wobei viele Schwann-Zellen hintereinander liegen, wenn der lange Neurit umhüllt und so zum Axon wird.
Bei der Bildung von Myelinscheiden, der Myelogenese oder Markreifung, wachsen die Schwann-Zellen mehrmals um den Nervenzellfortsatz herum und wickeln ihn derart bis zu fünfzigmal mit doppelten Lagen ihrer Zellmembran ein.[3] Dabei kommen die Zellmembranschichten unmittelbar aneinander zu liegen. Zwischen zwei Membraninnenseiten ist normalerweise kein Zytoplasma mehr vorhanden, sondern sie sind miteinander zur Hauptlinie oder inneren Anlagerungslinie verschmolzen, wo sich das basische Myelinprotein befindet.[2] Nicht verschmolzen sind die Zellmembranen an den Stellen der sogenannten Schmidt-Lantermann-Einkerbungen oder Myelininzisuren. Hier bestehen für den Stoffaustausch Zytoplasmabrücken, die sich als lichtmikroskopisch sichtbarer schmaler, schräg verlaufender Streifen durch alle Myelinschichten der Myelinscheide ziehen und miteinander verbunden sind.
Funktion
Nervenfasern mit stark ausgebildeten Myelinscheiden werden als markhaltig oder markreich bezeichnet und damit unterschieden von markarmen Fasern mit geringer ausgebildeter Myelinscheide und marklosen Fasern ohne Myelinscheide. Eine starke Myelinscheide schützt den umhüllten Neuriten mechanisch und isoliert ihn elektrisch vom umgebenden Milieu; der größere Abstand zwischen intrazellulärer und extrazellulärer Flüssigkeit senkt zudem die spezifische Membrankapazität.
Aktive Erregungsleitung ist langsamer als passive (rein elektrotonische) Erregungsleitung. Bei der saltatorischen Erregungsleitung werden Aktionspotentiale nur noch an den sogenannten Ranvier-Schnürringen aufgebaut, wo zwischen zwei umhüllenden Gliazellen die dicke Myelinscheide des Axons unterbrochen ist. Der jeweils circa 0,2 bis 1,5 Millimeter lange Axonabschnitt zwischen zwei solchen Schnürringen wird als Internodium bezeichnet. In diesen internodalen Segmenten verringert die isolierende Myelinscheide Leckströme durch die Membran, weshalb die elektrotonische Ausbreitung der Depolarisation verlustarm über diese weite Strecke möglich ist (größere Membranlängskonstante). Die verringerte Membrankapazität beschleunigt dabei die Umladung der Membran (kleinere Membranzeitkonstante).
Erkrankungen
Bei der Multiplen Sklerose und anderen demyelinisierenden Erkrankungen wird ebendiese Myelinhülle der Nervenzellfortsätze streckenweise zerstört – im Fall der MS durch Autoantikörper wie Anti-MOG-Antikörper und Anti-MBP-Antikörper. Dieser im Erkrankungsschub ablaufende Prozess wird als eine entzündliche Demyelinisation bezeichnet.
Einzelnachweise
- Renate Wahrig-Burfeind (Hrsg.): Wahrig. Illustriertes Wörterbuch der deutschen Sprache. ADAC-Verlag, München 2004, ISBN 3-577-10051-6, S. 590.
- Ulrich Welsch: Sobotta Lehrbuch Histologie. Zytologie, Histologie, mikroskopische Anatomie. 2., völlig überarbeitete Auflage. Elsevier, Urban & Fischer, München u. a. 2006, ISBN 3-437-44430-1, S. 189.
- Ulrich Welsch: Sobotta Lehrbuch Histologie. Zytologie, Histologie, mikroskopische Anatomie. 2., völlig überarbeitete Auflage. Elsevier, Urban & Fischer, München u. a. 2006, ISBN 3-437-44430-1, S. 186–188.