Pachygenelus
Pachygenelus ist eine ausgestorbene Gattung der Tritheledontidae, eine Familie der Cynodontia und lebte im „unteren“ Unterjura (Hettangium – Sinemurium)[4] vor ca. 201,3 bis 190,8 Millionen Jahren. Entsprechende Fossilien wurden in der Karoo-Supergruppe in Südafrika[1] und der oberen Fundy Gruppe in Nova Scotia (Kanada)[2] gefunden. Die Zuordnung von Funden aus der Obertrias (Dockum Gruppe) von Texas zu Pachygenelus („Pachygenelus milleri“) durch Chatterjee, 1983[3] ist hingegen umstritten und zweifelhaft.[2][5]
| Pachygenelus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Zeichnung von Pachygenelus | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Hettangium bis Sinemurium | ||||||||||||
| 201,3 bis 190,8 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Pachygenelus | ||||||||||||
| Watson, 1913[1] | ||||||||||||
| Arten | ||||||||||||
Etymologie und Forschungsgeschichte
Der Gattungsname setzt sich zusammen aus dem altgriechischen παχύς (pachys), „dick“, „fett“ und der latinisierten Verkleinerungsform des altgriechischen γένῠς (genys), „(Unter-)Kiefer“. Pachygenelus lässt sich, sehr grob, also in etwa mit „Dickkieferchen“ übersetzen.
Die Erstbeschreibung von Gattung und Typusart erfolgte 1913 durch David Meredith Seares Watson. Watson hatte während eines Aufenthaltes in der damaligen Kapkolonie vom Landvermesser Maasdorp einige fossile Knochenreste erhalten und insbesondere die Bedeutung eines Unterkieferbruchstückes erkannt. Er ließ sich die Fundstelle zeigen und identifizierte die Fundschicht als „Red Beds“ innerhalb der Stormberg Gruppe.[1]
1929 stellte Broom Pachygenelus gemeinsam mit „Karoomys“ (= Cynognathus[6]) und Tritheledon in die Gruppe der Ictidosauria („Wieselechsen“).[7] 1932 erweiterte Broom die Gruppe um zwei weitere Funde die er zunächst neutral als „Ictidosaurier A“ und „Ictidosaurier B“ bezeichnete.[8] Crompton beschrieb diese beiden Fossilien 1958[9] bzw. 1963[10] als Diarthrognathus broomi. Hopson und Kitching widersprachen 1972 dieser Analyse und werteten Diarthrognathus broomi als Juniorsynonym zu Pachygenelus monus. Gleichzeitig wurde Brooms „Unterordnung“ Ictidosauria in die zuvor von Broom aufgestellte „Familie“ der Tritheledontidae[11] integriert bzw. mit dieser gleichgesetzt.[6] Erst 1980 gelang Gow durch eine Detailanalyse der Bezahnung von zuvor noch unbeschriebenem Museumsmaterial der Nachweis, dass es sich bei Pachygenelus monus und Diarthrognathus broomi eindeutig um zwei verschiedene Taxa handeln musste.[12]
1983 beschrieb Chatterjee eine zweite Art („Pachygenelus milleri“) aus der Dockum Gruppe von Texas.[3] Die Zuordnung des Fundes zur Gattung Pachygenelus gilt aufgrund des nur schlecht erhaltenen Fossilmaterials jedoch als umstritten. Zum Teil wird sogar angezweifelt, dass es sich überhaupt um die Überreste eines Vertreters der Eucynodontia handelt.[2]
1991 wurde durch Shubin et al. ein weiterer Vertreter der Gattung Pachygenelus aus der McCoy Brook Formation innerhalb der Fundy Gruppe im Südosten Kanadas beschrieben. Nachdem das kanadische Belegmaterial keinerlei Merkmale aufwies die eine eindeutige Unterscheidung zu Pachygenelus monus zugelassen hätten, verzichteten die Erstbeschreiber auf die Aufstellung einer neuen Art und wiesen den Funden im Sinne einer „offenen“ (d. h. nicht durch den ICZN geregelten) Nomenklatur[13] die Bezeichnung Pachygenelus cf. P. monus zu.[2] Die Abkürzung „cf.“ steht dabei für das lateinische „confer“ „vergleiche (mit)“.
In der ersten Hälfte der 1990er-Jahre erschien zudem eine ganze Serie von zusammenfassenden Arbeiten zur frühen Evolution der Säugetiere in denen auch Pachygenelus umfassende Aufmerksamkeit gewidmet wurde.[14][15][16]
2001 beschrieb Gow erstmals eindeutig Pachygenelus monus zuordenbare postkraniale Skelettelemente aus der oberen Elliot-Formation (Stormberg Gruppe) von Südafrika.[17]
2016 wurden am teilweise erhaltenen Schädel eines juvenilen Exemplars (BPI/1/5691) von P. monus computertomographische Scans durchgeführt, die insbesondere Hinweise auf Verlauf und Ausbildung des Nervus infraorbitalis lieferten.[18]
Fossilbeleg
Das zweifelhafte Taxon „Pachygenelus milleri“ wird hier in weiterer Folge nicht mehr berücksichtigt.
Pachygenelus monus
Der ursprünglich von Watson beschriebene Holotyp (BMNH R4091) umfasst das Bruchstück eines rechten Unterkieferastes mit den Wurzeln zweier Schneidezähne, eines Eckzahns und 5 Backenzähnen.[1] Dazu kommt noch weiteres fossiles Belegmaterial (Auswahl der am häufigsten in der Fachliteratur erwähnten Belegstücke):
- BPI 4381: Ein teilweise erhaltenes Schädelskelett[12], wahrscheinlich ident mit BP/1/4381 im Sinne von Abdala, 2007[19]
- SAM K1315: Teile des Ober- und Unterkiefers[12]
- SAM K1329: Ein teilweise erhaltenes Schädelskelett[12][14]
- SAM K1350: kleines, aber nahezu vollständig erhaltenes Schädelskelett[12][14]
- SAM K1394: Ein teilweise erhaltener Unterkiefer[12]
- BP/1/5623: Teile des Schädels und des postkranialen Skeletts mit Teilen von 19 mehr oder weniger artikulierten Wirbeln, mehreren Rippen, dem vollständigen Schultergürtel und einem Großteil der vorderen Gliedmaßen.[17]
- BPI/1/5691: Schädel eines juvenilen Exemplars.[18]
Pachygenelus cf. P. monus
Das kanadische Fossilmaterial umfasst mehrere, zum Teil sehr gut erhaltene, einzelne Schädelfragmente, darunter 2 Prämaxillare, 10 Maxillae und 6 Dentale mit Teilen der Bezahnung.[2]
Merkmale
Pachygenelus monus wurde von Watson auf Basis des Holotyps als „etwa von der Größe eines Foxterriers“ beschrieben.[1] Die bislang bekannten Skelettelemente des Schultergürtels und der Vordergliedmaßen zeigen keine anatomische Anpassung an eine grabende Lebensweise wie sie etwa von Vertretern der verwandten Tritylodontidae bekannt ist, sondern gleichen denen basaler Vertreter der Mammaliaformes wie etwa Morganucodon. Als typisches Reptilienmerkmal ist hingegen zwischen dem eigentlichen Schlüsselbein (Clavicula) und dem Brustbein (Sternum) noch ein, sehr robust ausgebildetes, T-förmiges „Zwischenschlüsselbein“ (Interclavicula) vorhanden.[17]
Kiefergelenk
Pachygenelus hatte ein für die Ictidosauria (Tritheledontidae) charakteristisches doppeltes Kiefergelenk mit einer Gelenksverbindung sowohl zwischen Os articulare und Os quadratum (primäres Kiefergelenk) als auch über die für Säugetiere typische Verbindung zwischen Dentale und dem Schuppenteil (Pars squamosa) des Schläfenbeins (sekundäres Kiefergelenk). Nur Säugetiere haben ein sekundäres Kiefergelenk, während alle anderen Tetrapoden ein primäres Kiefergelenk besitzen. Über den Grad der Ausbildung des sekundären Kiefergelenks bei Pachygenelus herrscht zunächst Uneinigkeit. Gow, 1980 zweifelte die Existenz einer Gelenksverbindung zwischen Dentale und Pars squamosa an.[12] Allin & Hopson, 1992[14] und Luo & Crompton, 1994[15] bestätigten jedoch den Kontakt zwischen Dentale und Pars squamosa, obschon Crompton, 1995 einräumt die Verbindung sei bei Pachygenelus möglicherweise noch nicht zu einem wohldefinierten Synovialgelenk ausgebildet gewesen.[16]
Gebiss
Sofern nicht anders angegeben erfolgt die Beschreibung für Pachygenelus monus nach Gow, 1980.[12]
Pachygenelus trägt in jedem Kieferast nur zwei Schneidezähne. Die unteren Schneidezähne sind nach vorne geneigt, die Abnutzungsfacetten liegen zungenseitig (lingual) bzw. an den Spitzen. Die oberen Schneidezähne sind ventral ausgerichtet und leicht nach hinten gekrümmt.
Die Anordnung der Eckzähnen ist dem gegenüber umgekehrt; die oberen Eckzähne sind leicht nach vorne geneigt, während die unteren gerade nach oben ausgerichtet sind. Bei den meisten Vertretern der Cynodontia finden sich im knöchernen Gaumen anterio-lingual zu den oberen Eckzähnen tiefe Gruben die bei geschlossenem Gebiss die unteren Eckzähne aufnehmen. Solche Gruben fehlen bei Pachygenelus. Stattdessen okkludieren die unteren Eckzähne, ähnlich wie bei Säugetieren, jedoch in geringfügig abweichender Position, mit den oberen Eckzähnen.
Die Anzahl der Backenzähne scheint innerhalb der Art variabel zwischen 7 und 10 pro Kieferast zu liegen.[17] Sie verfügen über jeweils nur eine Wurzel. Ober- und Unterkiefer tragen jeweils Backenzähne unterschiedlicher Form. Die Backenzähne des Oberkiefers weisen eine konische Hauptspitze und zwei mesial bzw. distal befindliche Nebenspitzen. Die Backenzähne des Unterkiefers sind in der Regel größer als die des Oberkiefers und seitlich abgeflacht. Die vorderen Backenzähne des Unterkiefers zeigen eine mesiale Hauptspitze und zwei distale Nebenspitzen. Die hinteren Backenzähne zeigen zusätzlich noch eine kleine, mesiale Nebenspitze vor der Hauptspitze und eine dritte distale Nebenspitze.
Der Zahnwechsel erfolgte permanent (wie bei Reptilien) und alternierend, d. h. neue Zähne brachen jeweils zwischen zwei voll ausgebildeten Zähnen hervor. Die oberen und unteren Backenzahnreihen okkludieren miteinander, allerdings nicht so wie bei den Säugetieren, sondern seitlich, so dass einfache Abnutzungsfacetten und entsprechende scharfe Schneidkanten des Zahnschmelzes bei den oberen Backenzähnen lingual und bei den unteren (bukkal) ausgebildet wurden.
Ein weiteres Merkmal von Pachygenelus, das auch bei Säugetieren vorkommt, ist der plesiomorphe, prismatische Zahnschmelz.[20]
Fell und Schnurrhaare
Computertomographische Scans am Schädel eines juvenilen Exemplars (BPI/1/5691) zeigen, dass Pachygenelus, analog zu den Säugetieren, im Oberkiefer bereits einen kurzen, vollständig knochenummantelten Unteraugenkanal (Canalis infraorbitalis) für den Nervus infraorbitalis aufwies. Die äußere Öffnung des Unteraugenkanals (Foramen infraorbitale) über die in weiterer Folge die Haut des gesamten vorderen und oberen Gesichts sensorisch versorgt wird, weist ähnliche Dimensionen auf wie bei modernen Kleinsäugern. Bei letzteren korreliert die Größe der Öffnung insbesondere mit der Anzahl der Vibrissen und der Befund lässt den Schluss zu, dass bei Pachygenelus zumindest die anatomischen Voraussetzungen für Schnurrhaare vorhanden waren und das Tier solche mit hoher Wahrscheinlichkeit auch besessen hat.[18]
Ein Parietalforamen ist, ebenfalls analog zu modernen Säugetieren, bei Pachygenelus nicht vorhanden. Ausbildung oder Fehlen dieser Schädelöffnung wird im Wesentlichen durch das homöotische Gen Msx2 gesteuert, das bei Säugetieren ebenfalls für die Ausbildung des Fellkleides und der Milchdrüsen verantwortlich ist. Der Interpretation von Benoit et al., 2016 folgend, kann demnach davon ausgegangen werden, dass bei Pachygenelus ebenso bereits die genetischen Voraussetzungen für eine Fellbedeckung und die Ausbildung von Milchdrüsen vorhanden waren.[18]
Systematik
Über die systematische Stellung von Pachygenelus herrscht in Fachkreisen keine Einigkeit. Allgemein anerkannt ist die Bedeutung des relativ gut bekannten und erforschten Taxons für das Verständnis der frühen Entwicklungsgeschichte der Mammaliamorpha. Die folgende Zusammenstellung ist als Kurzübersicht der wesentlichsten phylogenetischen Hypothesen zu Pachygenelus zu lesen und in keinster Weise vollständig.
„Ictidosauria“ vs. Tritheledontidae vs. Pachygenelidae/Pachygenelinae
Weitgehend einig ist man sich dahingehend, dass Pachygenelus eine Gattung der Tritheledontidae repräsentiert. Diese Gruppe, zuweilen auch als „Tritheledonta“ bzw. (fälschlich) als „Trithelodonta“ oder „Trithelodontidae“ bezeichnet, wurde von Hopson & Kitching, 1972 mit den Ictidosauria gleichgesetzt und gemeinsam mit den Tritylodontidae in die Gruppe der Cynodontia gestellt.[6]
Bonaparte, 1980 schlug vor die „Pachygenelidae“ (Pachygenelus und Chaliminia) als eigenständige Klade von den Tritheledontidae (Diarthrognathus und Tritheledon) abzutrennen. „Pachygenelidae“ und Tritheledontidae sollten dem Vorschlag entsprechend eine übergeordnete Klade „Ictidosauria“ bilden.[21] Dieser Vorschlag wurde zwar allgemein zur Kenntnis genommen, stieß bei anderen Fachautoren jedoch auf nur sehr geringe Zustimmung.
| |||||||||||||||||||||||||||||||||||||||||||||
| Systematische Stellung von Pachygenelus innerhalb der „Ictidosauria“ nach Martinelli & Rougier, 2007.[22] |
Martinelli & Rougier definieren die Ictidosauria 2007 als „der letzte gemeinsame Vorfahre von Riograndia guaibensis und Pachygenelus monus und alle seine Nachfahren“. Die Tritheledontidae sehen sie als Teilgruppe der „Ictidosauria“, definiert als „der letzte gemeinsame Vorfahre von Irajatherium hernandezi und Pachygenelus monus und alle seine Nachfahren“. Für die Tritheledontidae werden nochmals zwei Untergruppen, die Chalimininae (Chaliminia + Elliotherium) und die Pachygenelinae (Pachygenelus + Diarthrognathus + Tritheledon) eingeführt und dem Taxon Irajatherium gegenübergestellt.[22]
Die Unterteilung der Tritheledontidae durch Martinelli & Rougier, 2007 weicht damit nicht nur signifikant vom ursprünglichen Vorschlag durch Bonaparte, 1980 ab, die Autoren verwenden für die Klade um Pachygenelus auch die Bezeichnung „Pachygenelinae“ anstatt des von Bonaparte vorgeschlagenen Namens „Pachygenelidae“. 2017 übernehmen Bonaparte & Crompton zwar diese veränderte Unterteilung, benutzen jedoch wieder die ursprüngliche Bezeichnung „Pachygenelidae“.[23]
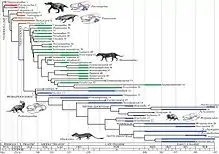
Systematische Stellung zu den Säugetieren (Mammalia)
In Bezug auf die verwandtschaftlichen Beziehungen zwischen Mammalia, Tritylodontidae und Tritheledontidae (und damit auch Pachygenelus) existieren im Wesentlichen zwei unterschiedliche Lehrmeinungen:[4]
Die eine „Schule“ vertritt die „klassische“ Lehrmeinung nach der die Tritheledontidae das am nächsten verwandte Schwester-Taxon der frühen Mammalia darstellen.[15][25][26][23]
Die zweite „Schule“ sieht in den Tritylodontidae das am nächsten verwandte Schwester-Taxon der frühen Mammalia und interpretiert die Tritheledontidae als Nebenzweig der sich bereits früher von der gemeinsamen Entwicklungslinie abgespalten hat.[27][28][29][24]
Palökologie
Für Pachygenelus wird eine weitgehend carnivore Ernährungsweise angenommen.[16] Obwohl anatomische Hinweise auf eine grabende Lebensweise, wie bei Vertretern der Tritylodontidae, nicht erkennbar sind,[17] wird Pachygenelus zumindest von einigen Autoren, mit Vorbehalten, als möglicher Verursacher von fossilen Grabbauten aus der oberen Elliot-Formation Südafrikas in Betracht gezogen.[30]
Die Fossilien der südafrikanischen Art Pachygenelus monus stammen überwiegend aus der oberen Elliot-Formation innerhalb der Stormberg-Gruppe. Die obere Elliot-Formation wird von Sedimenten eines fluvio-lakustrinen Ablagerungsmilieus unter semiariden klimatischen Bedingungen, mit nur saisonal wasserführenden, mäandrierenden Flüssen, deren Schwemmkegeln und flachen, playa-artigen Seen, durchsetzt mit äolischen Dünensedimenten aufgebaut.[31] Die artenarme Paläoflora der oberen Elliot-Formation enthält Vertreter der Bennettitales (Otozamites), Schachtelhalmen, Koniferen (Sphenolepidium) und weiteren verholzenden Nacktsamigen Pflanzen wie etwa Podocarpoxylon oder Agathoxylon.[32]
Pachygenelus monus ist ein Vertreter der sogenannten „Massospondylus Range Zone“[33] (MRZ), einer Faunen-Assoziation, die für die obere Elliot-Formation charakteristisch ist. Namensgebend für die MRZ ist Massospondylus aus der Gruppe der Sauropodomorpha, dessen stratigraphisches Auftreten innerhalb der Elliot-Formation sich weitgehend mit dem von Pachygenelus monus deckt. Weitere Mitglieder dieser Faunen-Assoziation, und damit Zeitgenossen die sich mit Massospondylus und Pachygenelus monus denselben Lebensraum geteilt haben, sind mindestens drei weitere Vertreter der Sauropodomorpha (Aardonyx, Pulanesaura und Arcusaurus)[34], die Theropoden Coelophysis rhodesiensis (=Megapnosaurus rhodesiensis) und Dracovenator[35], sowie mehrere Vertreter der Ornithischia wie etwa Heterodontosaurus, Lycorhinus, Lesothosaurus oder Stormbergia.[36]
Neben Pachygenelus monus treten Diarthrognathus und die Typusgattung Tritheledon als weitere Vertreter der Tritheledontidae innerhalb der oberen Elliot-Formation in Erscheinung. Die Tritylodontidae sind ebenfalls durch ihre Typusgattung Tritylodon vertreten;[4] die basalen Mammaliaformes hingegen mit Erythrotherium und Megazostrodon.[37] Fossile Überreste von stereospondylen Amphibien,[38] von Vertretern der Crocodylomorpha wie etwa Sphenosuchus, Litargosuchus, Protosuchus oder Orthosuchus[39] und von der Schildkröte Australochelys africanus[40] ergänzen das Bild der zeitgenössischen lokalen Wirbeltierfauna.
Über die Fauna der Wirbellosen ist dagegen nur wenig bekannt. Aus den hangendsten Anteilen der oberen Elliot-Formation sind aus den Ablagerungen nur zeitweilig wasserführender Seen die Schalen von Conchostracen belegt[41] und Xing et al. beschrieben 2015 Bohrspuren an einem Dinosaurierknochen die sie dem Ichnogenus Cubiculum zuwiesen. Diese Spuren sind wahrscheinlich auf die Larven aasfressender Käfer zurückzuführen.[42]
Einzelnachweise
- D. M. S. Watson: On a new Cynodont from the Stormberg In: The Geological Magazine - New Series, Dec. V, Vol. X, No. IV, S. 145–148, 1913. (Digitalisat)
- N. H. Shubin, A. W. Crompton, H.-D. Sues & P. E. Olsen: New Fossil Evidence on the Sister-Group of Mammals and Early Mesozoic Faunal Distributions. In: Science, Vol. 251, S. 1063–1065, 1991. (Digitalisat)
- S. Chatterjee: An Ictidosaur Fossil from North America. In: Science, Vol. 220, No. 4602, S. 1151–1153, 1983. (Abstract)
- C. A. Sidor & P. J. Hancox: Elliotherium kersteni, a new Tritheledontid from the Lower Elliot Formation (Upper Triassic) of South Africa. In: Journal of Paleontology, Vol. 80, No. 2, S. 333–342, 2006 (Digitalisat)
- J. W. Martz, B. Mueller, S. J. Nesbitt, M. R. Stocker, W. G. Parker, M. Atanassov, N. Fraser, J. Weinbaum & J. R. Lehane: A taxonomic and biostratigraphic re-evaluation of the Post Quarry vertebrate assemblage from the Cooper Canyon Formation (Dockum Group, Upper Triassic) of southern Garza County, western Texas. In: Earth and Environmental Science Transactions of the Royal Society of Edinburgh, Vol. 103, S. 1–26, 2013. (Digitalisat)
- J. A. Hopson & J.W. Kitching: A Revised Classification of Cynodonts (Reptilia; Therapsida). In: Palaeontologia Africana, Vol. 14, S. 71–85, 1972. (Digitalisat)
- R. Broom: On some recent new Light on the Origin of Mammals. In: Proceedings of the Linnean Society of New South Wales, Vol. 54, S. 688–694, 1929. (Digitalisat)
- R. Broom: The mammal-like reptiles of South Africa and the origin of mammals. 376 S., H. F. & G. Witherby, London, 1932.
- A. W. Crompton: The cranial morphology of a new genus and species of ictidosauran. In: Proceedings of the Zoological Society of London, Vol. 130, S. 183–216, 1958.
- A. W. Crompton: On the lower jaw of Diarthrognathus and the origin of the mammalian lower jaw. With appendix: on the statics of jaw musculature by D. G. Parkyn. In: Proceedings of the Zoological Society of London, Vol. 140, S. 697–753, 1963. (Digitalisat)
- R. Broom: On a New Type of Cynodont from the Stormberg. In: Annals of the South African Museum, Vol. 7, S. 334–336, 1912. (Digitalisat)
- C. E. Gow: The dentitions of the Tritheledontidae (Therapsida: Cynodontia). In: Proceedings of the Royal Society of London, Series B, Vol. 208, S. 461–481, 1980. (Digitalisat)
- S. C. Matthews: Notes on Open Nomenclature and on Synonymy Lists, In: Palaeontology, Vol. 16, Part 4, S. 713–719, 1973. (Digitalisat)
- E. F. Allin & J. A. Hopson, J.A.: Evolution of the Auditory System in Synapsida („Mammal-Like Reptiles“ and Primitive Mammals) as Seen in the Fossil Record. In: O. B. Webster, R. A. Fay & A. N. Popper (Hrsg.): The Evolutionary Biology of Hearing. S. 587–614, Springer Verlag, New York, 1992. (Digitalisat)
- Z. Luo & A. W. Crompton: Transformation of the Quadrate (Incus) through the Transition from Non-Mammalian Cynodonts to Mammals. In: Journal of Vertebrate Paleontology, Vol. 14, No. 3, S. 341–374, 1994. (Digitalisat)
- A. W. Crompton: Masticatory function in nonmammalian cynodonts and early mammals. In: J. Thomason (Hrsg.): Functional Morphology in Vertebrate Paleontology, S. 55–75, Cambridge University Press, 1995. (Digitalisat)
- C. E. Gow: A Partial Skeleton of the Tritheledontid Pachygenelus (Therapsida: Cynodontia) In: Palaeontologia Africana, Vol. 37, S. 93–97, 2001. (Digitalisat)
- J. Benoit, P. R. Manger & B. S. Rubidge: Palaeoneurological clues to the evolution of defining mammalian soft tissue traits. In: Nature – Scientific Reports, Vol. 6, No. 25604, S. 1–10, 2016 doi:10.1038/srep25604
- F. Abdala: Redescription of Platycraniellus elegans (Therapsida, Cynodontia) from the Lower Triassic of South Africa, and the Cladistic Relationships of Eutheriodonts. In: Palaeontology, Vol. 50, Part 3, S. 591–618, 2007. (Digitalisat)
- C. B. Wood, E. R. Dumont & A. W. Crompton: New studies of enamel microstructure in Mesozoic Mammals: a review of enamel prisms as a mammalian synapomorphy. In: Journal of Mammalian Evolution, Vol. 6, No. 2, S. 177–213, 1999. (abrufbar)
- J. F. Bonaparte: El primer ictidosaurio (Reptilia-Therapsida) de América del Sur, Chaliminia musteloides, del Triásico Superior de La Rioja, Argentina. In: Actas II Congreso Argentino de Paleontología y Bioestratigrafía y I Congreso Latinoamericano de Paleontología, Vol. 1, S. 123–133, 1980.
- A. G. Martinelli & G. W. Rougier: On Chaliminia musteloides (Eucynodontia: Tritheledontidae) from the Late Triassic of Argentina, and a Phylogeny of Ictidosauria In: Journal of Vertebrate Paleontology, Vol. 27, No. 2, S. 442–460, 2007. (Digitalisat)
- J. F. Bonaparte & A. W. Crompton: Origin and relationships of the Ictidosauria to non-mammalian cynodonts and mammals. In: Historical Biology, Vol. 30, Issue 1–2, S. 174–182, 2018. Online veröffentlicht am 23.06.2017 doi:10.1080/08912963.2017.1329911 (pdf)
- M. Ruta, J. Botha-Brink, S. A. Mitchell & M. J. Benton: The radiation of cynodonts and the ground plan of mammalian morphological diversity In: Proceedings of the Royal Society of London B: Biological Sciences, Vol. 280, Issue 1769, 2013. doi:10.1098/rspb.2013.1865
- J. A. Hopson & J. W. Kitching: A Probainognathian Cynodont from South Africa and the Phylogeny of Nonmammalian Cynodonts. In: Bulletin of the Museum of Comparative Zoology at Harvard College, Vol. 156, No. 1, S. 5–35, 2001. (Digitalisat)
- B. S. Rubidge & Ch. A. Sidor: Evolutionary Patterns Among Permo-Triassic Therapsids. In: Annual Review of Ecology and Systematics, Vol. 32, S. 449–480, 2001. (Digitalisat)
- T. Rowe: Definition, Diagnosis, and Origin of Mammalia. In: Journal of Vertebrate Paleontology, Vol. 8, No. 3, S. 241–264, 1988. (Digitalisat)
- T. Rowe: Phylogenetic Systematics and the Early History of Mammals. In: F. S. Szalay, M. J. Novacek & M. C. McKenna (Hrsg.): Mammal Phylogeny – Mesozoic Differentiation, Multituberculates, Monotremes, Early Therians, and Marsupials. S. 129–145, Springer Verlag, 1993 (Digitalisat)
- R. N. Martinez, C. L. May & C. A. Forster: A new carnivorous cynodont from the Ischigualasto Formation (Late Triassic, Argentina), with comments on eucynodont phylogeny In: Journal of Vertebrate Paleontology, Vol. 16, No. 2, S. 271–284, 1996. (Digitalisat)
- E. M. Bordy, L. Sciscio, F. Abdala, B. W. McPhee & J. N. Choiniere: First Lower Jurassic vertebrate burrow from southern Africa (upper Elliot Formation, Karoo Basin, South Africa) In: Palaeogeography, Palaeoclimatology, Palaeoecology, Vol. 468, S. 362–372, 2017. (Abstract)
- L. Sciscio, E. M. Bordy, M. Abrahams, F. Knoll & B. W. McPhee: The first megatheropod tracks from the Lower Jurassic upper Elliot Formation, Karoo Basin, Lesotho. In: PLoS ONE, Vol. 12, No. 10: e0185941, 2017. doi:10.1371/journal.pone.0185941
- L. Sciscio, F. Knoll, E. M. Bordy, M. O. de Kock & R. Redelstorff: Digital reconstruction of the mandible of an adult Lesothosaurus diagnosticus with insight into the tooth replacement process and diet In: PeerJ, 5:e3054, 2017. doi:10.7717/peerj.3054
- J. W. Kitching & M. A. Raath: Fossils from the Elliot and Clarens Formations (Karoo Sequence) of the Northeastern Cape, Orange Free State and Lesotho, and a suggested Biozonation based on Tetrapods In: Palaeontologia Africana, Vol. 25, S. 111–125, 1984. (abrufbar)
- B. W. McPhee, E. M. Bordy, L. Sciscio & J. N. Choiniere: The sauropodomorph biostratigraphy of the Elliot Formation of southern Africa: Tracking the evolution of Sauropodomorpha across the Triassic–Jurassic boundary In: Acta Palaeontologica Polonica, Vol. 62, No. 3, S. 441–465, 2017. (pdf)
- A. M. Yates: A new theropod dinosaur from the Early Jurassic of South Africa and its implications for the early evolution of theropods. In: Palaeontologia Africana, Vol. 41, S. 105–122, 2005. (Digitalisat)
- R. J. Butler: The `fabrosaurid´ ornithischian dinosaurs of the Upper Elliot Formation (Lower Jurassic) of South Africa and Lesotho. In: Zoological Journal of the Linnean Society, Vol. 145, Issue 2, S. 175–218, 2005. doi:10.1111/j.1096-3642.2005.00182.x
- C. E. Gow: A new Skull of Megazostrodon (Mammalia, Triconodonta) from the Elliot Formation (Lower Jurassic) of Southern Africa. In: Palaeontologia Africana, Vol. 26, No. 2, S. 13–23, 1986. (Digitalisat)
- A. Warren & R. Damiani: Stereospondyl Amphibians from the Elliot Formation of South Africa. In: Palaeontologia Africana, Vol. 35, S. 45–54, 1999. (Digitalisat)
- R. B. Irmis, S. J. Nesbitt & H.-D. Sues: Early Crocodylomorpha In: S. J. Nesbitt, J. B. Desojo & R. B. Irmis (Hrsg.): Anatomy, Phylogeny and Palaeobiology of Early Archosaurs and their Kin, Geological Society, London, Special Publications, Vol. 379, S. 275–302, 2013 (Digitalisat)
- E. S. Gaffney & J. W. Kitching: The Morphology and Relationships of Australochelys, an Early Jurassic Turtle from South Africa. In: American Museum Novitates, No. 3130, 29 S., American Museum of Natural History, New York, 1995. (abrufbar)
- E. M. Bordy, P. Eriksson: Lithostratigraphy of the Elliot Formation (Karoo Supergroup), South Africa. In: South African Journal of Geology, Vol. 118, No. 3, S. 311–318, 2015. (Digitalisat)
- L. Xing, A. H. Parkinson, H. Ran, C. A. Pirrone, E. M. Roberts, J. Zhang, M. E. Burns, T. Wang, J. Choiniere: The earliest fossil evidence of bone boring by terrestrial invertebrates, examples from China and South Africa. In: Historical Biology. 10 S., 2015. doi:10.1080/08912936.2015.1111884.