Lotka-Volterra-Gleichungen
Die Lotka-Volterra-Gleichungen (auch als Räuber-Beute-Gleichungen bekannt) sind ein System aus zwei nicht-linearen, gekoppelten Differentialgleichungen erster Ordnung. Sie beschreiben die Wechselwirkung von Räuber- und Beutepopulationen. Mit Räubern und Beute sind zwei Klassen von Lebewesen gemeint, deren eine sich von der anderen ernährt.[V 1] Aufgestellt wurden die Gleichungen 1925 von Alfred J. Lotka[1] und, unabhängig davon, 1926 von Vito Volterra.[2] Wesentliche Eigenschaften der Lösungen dieser Gleichungen sind als Lotka-Volterra-Regeln bekannt.
Die Ratengleichungen lauten

mit den Bezeichnungen[V 2]
| Anzahl der Beutelebewesen | zeitabhängig | |
| Reproduktionsrate der Beute ohne Störung und bei großem Nahrungsangebot | konstant | |
| Fressrate der Räuber pro Beutelebewesen = Sterberate der Beute pro Räuber | konstant | |
| Anzahl der Räuber | zeitabhängig | |
| Sterberate der Räuber, wenn keine Beute vorhanden ist | konstant | |
| Reproduktionsrate der Räuber pro Beutelebewesen | konstant |






Die Lotka-Volterra-Gleichungen sind eine wichtige Grundlage der Theoretischen Biologie, und darin insbesondere der Populationsdynamik. Bei den Räubern und der Beute muss es sich nicht unbedingt nur um Tiere oder einzelne Arten handeln; prinzipiell ist das Modell auf Gilden anwendbar – siehe z. B. Volterras Fischereidaten. Die Anwendbarkeit der Lotka-Volterra-Gleichungen hängt dabei davon ab, inwieweit die Begründung des mathematischen Modells im Einzelfall zutrifft.
Begründung des mathematischen Modells
Volterra begründet sein Gleichungssystem folgendermaßen:[V 3]
- Die Populationszahlen der Beute bzw. der Räuber seien mit bzw. bezeichnet.
- Die ungestörten Wachstumsraten pro Zeiteinheit seien und , wobei die Vorzeichen noch nicht fest liegen.
- Die (mittlere) Anzahl der Begegnungen zwischen Beute und Räuber pro Zeiteinheit ist mit einer positiven reellen Zahl , die innerhalb eines Biotops als konstant angenommen wird, aber im Allgemeinen vom Biotop abhängt.
- Eine genügend große Zahl Begegnungen haben im Mittel einen Effekt auf die Populationszahl . Bei den Beutelebewesen ist das ohne weiteres klar: Eine Begegnung mit einem Räuber führt mit einer gewissen Wahrscheinlichkeit dazu, dass die Beute gefressen wird. Dagegen ist die Auswirkung einer Begegnung auf die Zahl der Räuber nur indirekt, aber jedenfalls positiv; für die Modellbildung wird auch bei den Räubern eine sofortige Auswirkung auf die Populationszahl unterstellt.










Zusammengenommen führt das zu den Gleichungen

Division durch führt zu den Gleichungen

Setzt man

und führt den Grenzübergang durch, so erhält man die Lotka-Volterra-Gleichungen in der eingangs genannten Form.

Mathematische Behandlung
Natürlich war auch Volterra klar, dass die zeitabhängigen Populationszahlen und nur ganzzahlige Werte annehmen können und daher als Funktionen von entweder konstant oder nicht differenzierbar sind. Aber bei großen Populationszahlen ist der durch Übergang zum kontinuierlichen Modell gemachte relative Fehler gering. Der Vorteil der zweidimensionalen Lotka-Volterra-Gleichung ist jedoch, dass einige Aussagen mathematisch beweisbar sind, die einen interessanten Bezug zu realen Daten haben, wie weiter unten beschrieben ist.



Zur mathematischen Behandlung von Lotka-Volterra-Systemen benutzt man heute meist die etwas einfachere Notation[M 1]

wobei positive Konstanten sind und die Anzahl der Beutetiere und die Anzahl der Raubtiere (predators) bezeichnen.



Konstante Lösungen
Die konstanten Lösungen (auch Gleichgewichtspunkte oder kritische Punkte genannt) erhält man, indem man die rechten Seiten der Lotka-Volterra-Gleichungen gleich Null setzt:

Es gibt also genau zwei konstante Lösungen, nämlich den trivialen Gleichgewichtspunkt und den inneren Gleichgewichtspunkt


Ein erstes Integral
Eine Methode zum Auffinden nicht-konstanter Lösungen besteht darin, ein erstes Integral, also eine Invariante der Bewegung, zu suchen. Volterra findet eine solche auf folgendem Weg:[V 4] Multipliziert man die erste Grundgleichung mit und die zweite mit , und addiert anschließend die beiden Gleichungen, so verschwinden die Terme mit dem Produkt , und man erhält



- .

Durch Multiplikation der ersten Grundgleichung mit und der zweiten mit und anschließender Addition kommt man zu


- .


Subtraktion dieser beiden Gleichungen ergibt
- .

Durch Integration dieser letzten Gleichung erreicht man schließlich die Beziehung
- const.

Umgekehrt kann man die totale Ableitung der so definierten Funktion nach berechnen:

so gelangt man ebenfalls zu der Aussage, dass auf den Lösungen der Grundgleichungen konstant (invariant) ist; eine Lösung der Lotka-Volterra-Gleichung kann also ihre Niveaulinien von nicht verlassen.
Ein anderer Weg zum Auffinden einer Invarianten der Bewegung besteht darin, die Lotka-Volterra-Gleichungen mit Hilfe eines eulerschen Multiplikators in eine exakte Differentialgleichung umzuformen und diese dann zu integrieren.[W 1]
Stabilität
Da als erstes Integral auch eine Ljapunow-Funktion ist, und da am inneren Gleichgewichtspunkt ein striktes lokales Minimum besitzt, folgt aus dem ersten Kriterium von Ljapunow, dass dieser Gleichgewichtspunkt stabil ist.
Die Lotka-Volterra-Gesetze
Mit Hilfe des ersten Integrals beweist Volterra[V 5] drei mathematische Eigenschaften der Lösungen („Gesetze“) der Lotka-Volterra-Gleichungen, deren biologischen Interpretationen als Lotka-Volterra-Regeln Verbreitung gefunden haben.
Aus dem Randverhalten der Funktion kann man schließen, dass keine Trajektorie, die einen Punkt im ersten Quadranten

besitzt, diesen verlässt: der erste Quadrant ist invariant. Die Lotka-Volterra-Gesetze gelten allgemein für maximale Lösungen der Lotka-Volterra-Gleichungen in diesem Quadranten; stirbt eine der beiden Tierklassen aus, so wird dieser Quadrant verlassen und die Lotka-Volterra-Gesetze verlieren ihre Gültigkeit.
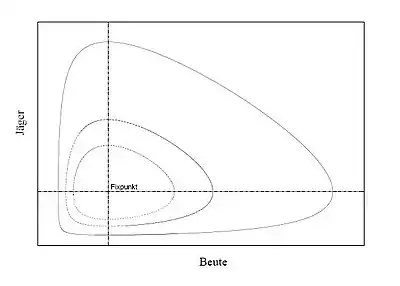
Periodizität
Da die Funktion im Quadranten strikt konvex ist und ihr Minimum im inneren Gleichgewichtspunkt annimmt, bilden die Niveaulinien von geschlossene Kurven im Phasenraum. Da jede Lösung in einer Niveauline von enthalten sein muss, folgt aus der Eindeutigkeit und einer Betrachtung des lokalen Richtungsfeldes die Periodizität der Lösungen.[V 6][W 2]

- Loi du cycle périodique. – Les fluctuations des deux espèces sont périodique.
- (Gesetz der periodischen Zykel: Die Schwankungen der beiden Klassen sind periodisch.)
Erhaltung der Mittelwerte
Aus der Periodizität der Lösungen folgt mit ein paar Zeilen Rechnung das
- Loi de la conservation des moyennes. – Les moyennes pendant une période des nombres des individus des deux espèces sont independent des conditions initiales, et égales aux nombres qui correspondent à l'ètat stationnaire, pour les valeurs données des «coefficients d’accroissement» , et des «coefficients de voracité» .


- (Gesetz der Erhaltung der Mittelwerte: Die zeitlichen Mittelwerte über eine Periode der Individuenzahlen der beiden Klassen hängen nicht von den Anfangsbedingungen ab, und sind gleich den Anzahlen des Gleichgewichtszustands, für die gegebenen „Wachstumskoeffizienten“ , und die „Fresskoeffizienten“ .)
Das bedeutet, die zeitlichen Mittelwerte erfüllen die Gleichungen

Auf den ersten Blick verwirrend ist hier, dass der Mittelwert der Beutetierpopulation nur von Sterbe- und Fressrate der Raubtierpopulation und nicht von der Reproduktionsrate der Beutetiere abhängt. Dagegen ist der Mittelwert der Raubtierpopulation nur von Reproduktions- und Sterberate der Beutetierpopulation und nicht von Fress- und Sterberate der Raubtiere abhängig. Dabei ist die gleichgewichtige Anzahl der Beute umso höher, je ungünstiger die Parameter für die Raubtiere sind. Die gleichgewichtige Anzahl der Räuber ist dagegen umso höher, je günstiger die Parameter für die Beutetiere sind.
Verständlich wird diese Eigenschaft des Lotka-Volterra-Modells, wenn man sich die in der mathematischen Begründung zur Anwendung kommende Modellbildung ansieht: die Kontrolle über die Populationszahl der einen Tierklasse obliegt hier ausschließlich der jeweils anderen Klasse.
Störung der Mittelwerte
Das wegen seiner biologischen Interpretation interessanteste dieser Gesetze ist das
- Loi de la perturbation des moyennes. – Si l’on détruit les deux espèces uniformément et proportionnellement aux nombres de leurs individus (assez peu pour que les fluctuations subsistent), la moyenne du nombres des individus de l’espèce dévorée croît et celles de l’espèce devorant diminue.
- (Gesetz der Störung der Mittelwerte: Werden die beiden Tierklassen gleichmäßig und proportional zur Populationsgröße dezimiert, und ist die Rate der Dezimierung der Beute kleiner als deren Reproduktionsrate, so wächst der Mittelwert der Beutepopulation, und der Mittelwert der Räuberpopulation sinkt.)
Tatsächlich beweist Volterra eine quantitative Version: Ist die Zerstörungsrate der Beutelebewesen, und die Zerstörungsrate der Räuber, so sind die Mittelwerte für die Lösungen der gestörten Lotka-Volterra-Gleichungen


Das bedeutet: die über eine Lotka-Volterra-Periode gemittelte Anzahl der Beutelebewesen steigt genau dann an, wenn die Räuber dezimiert werden – ziemlich unabhängig von einer Dezimierung der Beute, solange diese nicht ausgerottet wird. Umgekehrt sinkt die mittlere Anzahl der Räuber immer dann, wenn die Beutelebewesen dezimiert werden, und dieses Absinken hängt nicht davon ab, wie stark die Räuber zusätzlich dezimiert werden (solange diese nicht ausgerottet werden).
Erweiterungen
In der theoretischen Ökologie bilden die Lotka-Volterra-Gleichungen den Ausgangspunkt zur Entwicklung komplexerer Modelle, von denen einige bereits in Volterras Buch beschrieben sind.
Intraspezifische Konkurrenzterme
Eine erste Erweiterung der Lotka-Volterra-Gleichungen entsteht durch Subtraktion von Termen proportional zu bzw. , die die intraspezifische Konkurrenz modellieren.[W 3] Es gibt verschiedene Möglichkeiten, die Form und der neu hinzugekommenen Terme zu begründen:




- Mit den empirischen Untersuchungen zur Bevölkerungsentwicklung nach Pierre-François Verhulst, siehe logistische Gleichung.
- Durch die Annahme, die (ungestörte) Wachstumsrate einer Population sei proportional zur Differenz zwischen einer Kapazitätsgrenze und der tatsächlichen Populationszahl.
- Durch eine Analyse des Einflusses von intraspezifischen Begegnungen auf die Populationszahl ähnlich Volterras Begründung des Terms zur Modellierungen der interspezifischen Konkurrenz: Eine intraspezifische Begegnung ist mit einer gewissen Wahrscheinlichkeit eine Konkurrenz um eine Ressource, bei der ein Individuum den Kürzeren zieht.
Die daraus entstehenden Lotka-Volterra-Konkurrenzgleichungen der Theoretischen Biologie sind ein klassischer Ansatz zur Beschreibung der Dynamik einer stark vereinfachten Biozönose, bestehend aus einer nachwachsenden Ressource und mindestens 2 darum konkurrierender Arten:

- ,


wobei a,b exponentielle Wachstumsraten sind und die m (mortality rate) Sterberaten darstellen. Die zu einem Zeitpunkt verfügbare Ressourcenmenge wird angenommen als:

Damit ergibt sich:


durch Ausmultiplizieren und Ersetzung der Koeffizienten






kommt man zu Gleichungen der Form

die wiederum zwei Gleichgewichtslagen zulassen: den trivialen Gleichgewichtspunkt , und den inneren Gleichgewichtspunkt , der durch ein lineares Gleichungssystem gegeben ist:


Durch Lösen dieses Gleichungssystems findet man den Gleichgewichtspunkt

der unter der Bedingung im ersten Quadranten liegt.

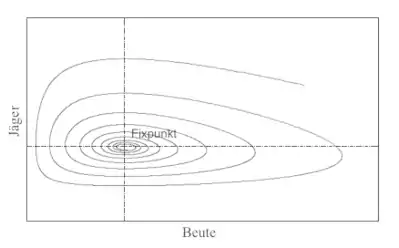
Zu diesem erweiterten Lotka-Volterra-Systems gibt es auch eine Ljapunow-Funktion:

mit der die Voraussetzungen des Zweiten Kriteriums von Ljapunow für den Gleichgewichtspunkt erfüllt sind. Daraus folgt, dass dieser Gleichgewichtspunkt jetzt asymptotisch stabil ist.[W 4]
Mehr als zwei Klassen von Lebewesen
Ein großer Teil von Volterras Buch bezieht sich auf Erweiterungen seines Systems auf mehr als zwei Klassen von Lebewesen, die in unterschiedlichen Weisen miteinander interagieren.
Anwendungen
Fischereidaten
In der Einleitung zu Volterras Buch[V 7] findet sich eine Tabelle, die zu den Jahren 1905 und 1910–1923 und zu drei Fischereihäfen jeweils den prozentualen Anteil der Knorpelfische (Sélaciens), also insbesondere der Haie, am gesamten Fischfang des Fischereihafens enthält:
| 1905 | 1910 | 1911 | 1912 | 1913 | 1914 | 1915 | 1916 | 1917 | 1918 | 1919 | 1920 | 1921 | 1922 | 1923 | |
| Trieste | – | 5,7 | 8,8 | 9,5 | 15,7 | 14,6 | 7,6 | 16,2 | 15,4 | – | 19,9 | 15,8 | 13,3 | 10,7 | 10,2 |
| Fiume | – | – | – | – | - | 11,9 | 21,4 | 22,1 | 21,2 | 36,4 | 27,3 | 16 | 15,9 | 14,8 | 10,7 |
| Venise | 21,8 | – | – | – | - | – | – | - | – | – | 30,9 | 25,3 | 25,9 | 26,8 | 26,6 |
- «Cela prouve pendant la période 1915–1920, où la pêche était moins intense à cause de la guerre, un accroissement relatif de la classe des Sélaciens qui, particulièrement voraces, se nourissent d’autres poissons. Les statistique inclinent donc à penser qu’une diminution dans l’intensité de la destruction favorise les espèces les plus voraces.»[V 8]
Diese Statistiken zeigen in den Jahren 1915 bis 1920, als der Fischfang im Mittelmeer wegen des Ersten Weltkriegs weniger intensiv war, einen erhöhten Anteil an Raubfischen, der dann mit der Intensivierung der Fischerei nach 1920 wieder zurückgeht. Das dritte Lotka-Volterra-Gesetz, die Verschiebung der Mittelwerte, bietet hierfür eine plausible Erklärung.
Medizinische Epidemiologie
In der Theoretischen Biologie sowie in der medizinischen Epidemiologie finden Modelle vom Lotka-Volterra-Typ zur Beschreibung der Ausbreitungsprozesse von Krankheiten Verwendung. Einige Beispiele finden sich in SI-Modell, SIR-Modell und SIS-Modell.
Wirtschaftswissenschaften
Dem Goodwin-Modell zur Erklärung von Konjunkturschwankungen liegen Lotka-Volterra-Gleichungen zugrunde, wobei der Lohnquote die Rolle des Räubers und der Beschäftigungsquote die Rolle der Beute zukommt.
Gerold Blümle entwickelte ein Konjunkturmodell, in dem (mathematisch) der Investitionsquote die Rolle der Raubtiere zukommt, und der Streuung oder Varianz der Gewinne die Rolle der Beutetiere.[3] Bei Frank Schohl kommt der Varianz der Renditenänderungen der Unternehmen die Rolle der Raubtiere, der Varianz der Angebotsänderungen der Unternehmen die Rolle der Beutetiere zu.[4]
Literatur
- Alfred J. Lotka: Analytical Theory of Biological Populations. Plenum Press, New York NY u. a. 1998, ISBN 0-306-45927-2, englische Übersetzung der beiden Bände
- Théorie analytique des associations biologiques (= Exposés de biométrie et de statistique biologique. Bd. 4 = Actualités scientifiques et industrielles. Bd. 187). Première partie: Principes. Hermann, Paris 1934.
- Théorie analytique des associations biologiques. Deuxième partie: Analyse démographique avec application particulière à l’espèce humaine (= Exposés de biométrie et de statistique biologique. Bd. 12 = Actualités scientifiques et industrielles. Bd. 780). Hermann, Paris 1939.
Einzelnachweise
- Elements of Physical Biology. 1925, S. 115
- Variazioni e fluttuazioni del numero d’individui in specie animali conviventi. In: Mem. R. Accad. Naz. dei Lincei. Ser. VI, vol. 2, 31-113.
- Gerold Blümle: Wachstum und Konjunktur bei Differenzgewinnen - Ein Schumpeter-Modell der wirtschaftlichen Entwicklung. In: H. J. Ramser, Hajo Riese (Hrsg.) Beiträge zur angewandten Wirtschaftsforschung. Gottfried Bombach zum 70. Geburtstag. Berlin 1989, S. 13–37. Dargestellt auch in Frank Schohl: Die markttheoretische Erklärung der Konjunktur. Schriften zur angewandten Wirtschaftsforschung. Tübingen 1999.
- Frank Schohl: Die markttheoretische Erklärung der Konjunktur. Schriften zur angewandten Wirtschaftsforschung. Tübingen 1999, S. 232.
- Vito Volterra: Leçons sur la Théorie Mathématique de la Lutte pour la Vie. Gauthier-Villars, 1931; autorisierter Nachdruck: Éditions Jaques Gabay, 1990, ISBN 2-87647-066-7
- chap. I, sec. II. Deux espèces dont l’une dévore l’autre
- S. 14
- S. 14f
- S. 15
- S. 15–27
- S. 15–19
- S. 2 ff.
- S. 2
- James D. Murray: Mathematical Biology I: An Introduction. 3. Auflage. Springer, 2008, ISBN 978-0-387-95223-9
- S. 79
- Günther J. Wirsching: Gewöhnliche Differentialgleichungen. Eine Einführung mit Beispielen, Aufgaben und Musterlösungen. Teubner, Wiesbaden 2006, ISBN 978-3-519-00515-5
- Beispiel 4.2, S. 65 ff.
- S. 67–70
- S. 80 ff.
- S. 82