Heteropterygidae
Die Heteropterygidae sind eine in Südostasien beheimatete Familie aus der Ordnung der Gespenstschrecken, zu welcher mehr als 130 beschriebene Arten gezählt werden (Stand Ende 2020).[1][2] Der für die Heteropterygidae stellenweise verwendete Name Riesengespenstschrecken ist irreführend, da er auch für andere nicht zu dieser Familie gehörende Arten verwendet wird. Außerdem erreichen lediglich einige Vertreter der Unterfamilie Heteropteryginae eine Größe, die diesen Namen rechtfertigen würde.
| Heteropterygidae | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Verschiedene Heteropterygidae Arten | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Heteropterygidae | ||||||||||||
| Kirby, 1896 | ||||||||||||
| Unterfamilien | ||||||||||||
Merkmale
Größe
Unter den Heteropterygidae finden sich sowohl sehr kleine, als auch sehr große und massige Arten. Dabei sind die Vertreter der Dataminae durchweg eher klein. So werden Planispectrum-Arten lediglich 1,9 bis 3,5 cm lang. Die größten Dataminae-Arten sind mit maximal 5,0 bis 6,0 cm Länge innerhalb der Gattungen Pylaemenes und Orestes zu finden. Während die Unterfamilie Obriminae mit knapp 3,0 cm bei Tisamenus hebardi und bis zu gut 13 cm Länge bei Trachyaretaon carmelae ein breites Spektrum an Größen zeigt, gelten die Heteropteryginae als eher groß. Zwar sind hier sehr große und massige Arten, wie die bis zu 17 Zentimeter lange Malaiische Riesengespenstschrecke (Heteropteryx dilatata) zu finden, aber auch kleine Arten wie die im männlichen Geschlecht nur 2,5 bis 3,7 cm lange Haaniella parva.[1][3][4]
Morphologie
Als gemeinsames autapomorphes Merkmal der Heteropterygidae gilt das Vorhandensein sensorischer Bereiche am Prosternum. Bei den Dataminae befindet sich sowohl ein Paar dieser sensorischen Felder auf dem Prosternum und mittig dahinter ein drittes auf dem sogenannten (Pro-)Furcasternit. Den Obriminae fehlt das hintere Feld, welches bei den Heteropteryginae vorhanden ist. Diesen wiederum fehlt das vordere Paar sensorischer Felder. Ein auffälliges morphologisches Merkmale ist der bei den Weibchen zur Mitte verbreiterte Hinterleib (Abdomen). Bei adulten Weibchen ist dieser Bereich nicht nur sehr breit, sondern durch die permanent und häufig in großer Zahl produzierten Eier auch deutlich erhöht. Bei den Weibchen der Unterfamilien Heteropteryginae und Obriminae endet das Abdomen in einem spitzen Legestachel, welcher den eigentlichen Ovipositor umgibt. Dieser wird ventral aus dem achten Sternit gebildet, welches hier als Subgenitalplatte,[5] oder auch Operculum bezeichnet wird. Dorsal wird der Legestachel bei den Heteropteryginae und den Obrimini aus dem als Supraanalplatte oder Epiproct bezeichneten elften Tergum gebildet. Bei den Hoplocloniini hingegen, wird dieser aus dem zehnten Tergum gebildet.[1][6] Die kleineren Männchen haben einen im Querschnitt rundlichen mittleren Abdomenbereich, welcher hier ganz im Gegensatz zu den Weibchen am dünnsten ist. Das verdickte Abdomenende wird bei ihnen ventral durch die Subgenitalplatte und dorsal vom achten, neunten und dem als Analsegment bezeichneten zehnten Tergum gebildet. Außer bei Miroceramia westwoodii und in reduzierter Form bei Pterobrimus depressus den jeweils einzigen Vertretern ihrer Gattungen finden sich bei den Vertretern der Obriminae und der Dataminae keine Flügel, während die Heteropteryginae Flügel haben.[7] Bei ihnen sind oft die Vorderflügel (Tegmina) und meist auch die Hinterflügel verkürzt, wie bei vielen Haaniella-Arten und den Weibchen der Malaiischen Riesengespenstschrecke (Heteropteryx dilatata). Bei einigen Haaniella-Männchen und denen von Heteropteryx dilatata bedecken die Tegmina fast das gesamte Abdomen. Die darunterliegenden, voll entwickelten Hinterflügel befähigen sie teilweise zu kurzen Flügen.
Acanthotaxie
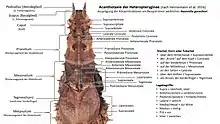
Der Körper kann mit zahlreichen spitzen Stacheln, mehr oder minder stumpfen Dornen oder Tuberkeln besetzt sein, die vor allem auf der Oberseite von Kopf und Thorax zu finden sind. Ihre Ausprägung ist sehr unterschiedlich und meist so artspezifisch, dass sie zur Bestimmung und Abgrenzung der Arten genutzt wird. Diese als Acanthotaxie bezeichnete Methode, wurde 1939 von James Abram Garfield Rehn & John W. H. Rehn für die Obriminae entwickelt,[8] 1998 und 2001 von Philip Edward Bragg für die Dataminae modifiziert und 2016 von Frank H. Hennemann et al. für die Heteropteryginae angepasst. Die jeweiligen Strukturen werden dabei nach ihrer Lage bezeichnet. Die Stacheln die sich auf dem Scheitel des schräg nach vorn unten gerichteten Kopfes befinden, wirken wie eine Krone und werden deshalb als Coronalen bezeichnet (Corona lat. für Kranz oder Krone). Die über der Basis der Antennen werden Supraantenalen, die auf dem Hinterhaupt Supraoccipitalen genannt (occiput lat. für Hinterhaupt, siehe: occipital). Stacheln auf dem Pro-, Meso- und Metanotum werden als Pronotalen, Mesonatalen beziehungsweise Metanotalen bezeichnet. Stacheln an den Meso- und Metapleuren werden allgemein als Lateralen bezeichnet und die Einzelstacheln über deren Coxen werden als Supracoxalen angesprochen.[1]
Verbreitungsgebiet
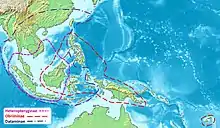
Das Zentrum des Verbreitungsgebietes der Heteropterygidae ist Borneo, wo Vertreter aller drei Unterfamilien vorkommen. Die Heteropteryginae haben das kleinste Verbreitungsgebiet. Es schließt neben Borneo nur den thailändischen Teil der malaiischen Halbinsel, Sumatra und Java ein. Auf dem kontinentalen Festland Asiens ist mit Haaniella gorochovi lediglich eine Hetropteryginae-Art im Süden Vietnams zu finden. Das Verbreitungsgebiet der Dataminae schließt die gesamte von den Heteropteryginae bewohnte Region ein. Darüber hinaus besiedeln sie auf dem asiatischen Festland das östliche Myanmar, Thailand, Laos, Kambodscha, Vietnam und Teile Südchinas. Östlich werden sowohl die Okinawa-Inseln, Taiwan, Palawan, die Molukkeninseln Buru und Seram, sowie sämtliche Kleinen Sundainseln und Sulawesi bewohnt. Das Verbreitungsgebiet der Obriminae erstreckt sich von Borneo aus nach Osten. Es schließt die Philippinen, Sulawesi, die meisten Molukkeninseln und Neuguinea ein. Noch weiter östlich befindet sich mit Viti Levu, der Hauptinsel der Fidschi-Gruppe das Verbreitungsgebiet von Pterobrimus depressus.[1][9]
Fortpflanzung und Lebenszyklus
Die adulten Weibchen der Heteropteryginae und der Obriminae legen ihre oft verhältnismäßig großen Eier mittels Legestachel fast immer einzeln, mehrere Zentimeter tief im Erdboden ab. Auch die Dataminae legen ihre Eier am oder im Boden ab. Bei Epidares und einigen Orestes-Arten wurde belegt, dass sie mit den Vorderbeinen eine Mulde im Boden vorbereiten, dann den Hinterleib schnell über den Vorderkörper klappen lassen, wobei sie ein Ei herauspressen. Dieses wird dabei nach vorn katapultiert, mit den Antennen gefangen und über diese in die Mulde abgerollt, wo es dann wiederum mittels der Vorderbeine mit Erde abgedeckt wird. Aus den Eiern schlüpfen nach zwei bis zwölf Monaten die Nymphen, welche selbst bei kleineren Arten mit mindestens zehn Millimeter Länge schon recht groß sind. Das Heranwachsen zur Imago dauert wiederum häufig bis zu einem Jahr. Neben Arten mit einer durchschnittlichen Lebenserwartung von mindestens sechs bis zwölf Monaten, gibt es viele die ein erstaunliches Alter erreichen. So konnte Oskar V. Conle bei einem Wildfangweibchen von Haaniella scabra ein Alter von mehr als fünf Jahren nachweisen. Auch von den Dataminae sind solche Lebensspannen belegt, etwa bei Dares verrucosus.[10]
Abwehrverhalten
Typisch für die kurzflügeligen Vertreter der Unterfamilie der Heteropteryginae, ist ein ausgeprägtes Abwehrverhalten, bei welchem der Gegner zunächst durch eine mit den Flügeln erzeugte Abwehrstridulation angedroht wird. Langflügeligen Vertretern fehlt die Fähigkeit zu stridulieren. Sie öffnen an die Flügel um größer zu erscheinen. Schließlich werden der Hinterleib und die bedornten Hinterbeine angehoben, deren Schienen bei taktilen Reizen klappmesserartig gegen die Schenkel geschlagen werden, was ein Einklemmen des Gegners zum Ziel hat. Im Gegensatz dazu sind die weniger bewehrten Arten deutlich besser getarnt. Obriminae setzten auf Tarnung und halten sich an Borke oder Ästen der Bäume fest auf denen sie meist tagsüber versteckt sind. Die Vertreter der Dataminae haben die Mimese von Borke und kurzen Ästen nahezu perfektioniert. Werden sie berührt, reagieren die meisten Arten mit einer Schreckstarre, bei der Beine und Fühler längs der Körperachse gelegt werden, um so den Eindruck eines kurzen Stöckchens zu erzeugen. Eine ausgeprägte Autotomie der Beine, wie sie bei vielen anderen Gespenstschrecken bekannt ist, findet sich bei den Heteropterygidae nicht.
Systematik





Systematischer Hintergrund
Karl Brunner von Wattenwyl errichtete 1893 für die seinerzeit bereits beschriebenen Gattungen Obrimus, Hoploclonia, Tisamenus, Pylaemenes, Dares und Datames (heute Synonym zu Pylaemenes) die Obrimini (dort als Obrimi. abgekürzt) und stellte diese gemeinsam mit der Gattung Heteropteryx und den Cladomorphini (dort als Cladomorphi. abgekürzt) in die Familie Cladomorphidae (heute Synonym zu Cladomorphinae).[11] William Forsell Kirby errichtete 1896 für die Gattung Heteropteryx die Unterfamilie Heteropteryginae innerhalb der Familie Bacillidae, ohne auf die Obrimini aus Brunners Arbeit Bezug zu nehmen. Im Jahr 1904 schloss er alle damals bereits bekannten Gattungen der heutigen Heteropterygidae, sowie die Gattung Parectatosoma in diese Unterfamilie ein.[12] Josef Redtenbacher führte 1906 die Gattungen der heutigen Obriminae und Dataminae wie Brunner in der Tribus Obrimini, während er die Gattungen Heteropteryx, Leocrates (heute Synonym zu Heteropteryx) und wiederum Parectatosoma, sowie die neu errichtete Anisacantha in der Tribus Heteropterygini führt. Allerdings weist er nicht auf die bereits von Kirby und Brunner festgestellte Verwandtschaft beider Triben hin.[13] Lawrence Bruner erhob 1915 die Obrimini in den Rang einer Familie. Rehn & Rehn teilten 1939 die von ihnen nur noch als Unterfamilie angesprochenen Obriminae in die Triben Obrimini und Datamini auf.[8] Klaus Günther überführte 1953 diese beiden Triben in die Unterfamilie Heteropteryginae. Deren bisherige südostasiatische Gattungen stellte er in die Heteroperygini. Für die madagassischen Gattungen Parectatosoma und Anisacantha errichtete er die Anisacanthini.[14] Oliver Zompro erhob 2004 die Unterfamilie in den Rang der Familie Heteropterygidae. Von den vier enthaltenen Triben wurden drei in den Rang von Unterfamilien gestellt, während die madagassischen Arten in die neu errichtete Familie Anisacanthidae überführt wurden. Als Erstautor ist Kirby 1896 zu priorisieren.[1][15][16]
Innere Systematik
In einer umfangreichen Arbeit wurden 2016 von Hennemann et al. unter anderem die Triben innerhalb der Obriminae neu bearbeitet und die Tribus Eubulidini synonymisiert.[1] In der ersten Genanalyse zur Klärung der Phylogenie einer Phasmidenfamilie wurden von Sarah Bank et al. sieben mitochondriale Gene und Gene aus dem Zellkern untersucht um die Verwandtschaftsverhältnisse der äußeren und inneren Systematik zu klären. In deren Ergebnis wurde unter anderem auch die zweite von Zompro aufgestellte Tribus Miroceramiini und die 2016 von Hennemann et al. aufgestellte Tribus Tisamenini der Obriminae wieder eingezogen. Außerdem wurde die von der Morphologie des Legestachels her bekannte Sonderstellung der Gattung Hoploclonia auch phylogenetisch bestätigt und dem durch Errichtung einer entsprechenden Tribus Rechnung getragen. Die Datamini wurden als die ursprünglichere Klade identifiziert. In den drei übrigen Triben hat sich der sekundäre Ovipositor dreimal unabhängig voneinander entwickelt hat. Die Vertreter Heteropterigini bilden zwar eine gemeinsame Klade, aber die Gattung Heteropteryx ist phylogenetisch mitten in mehreren Linien von aktuell in Haaniella geführten Arten zu platzieren. Dem folgend müsste entweder Haaniella in mehrere Gattungen aufgespalten werden oder zugunsten der früher beschrieben Gattung Heteropteryx eingezogen werden. Letzteres erscheint aufgrund fehlender, größerer autapomorpher Unterschiede wahrscheinlicher. Die Verwandtschaftsverhältnisse der untersuchten Heteropterygidae-Gattungen sind in folgendem Kladogramm dargestellt:[9]

| Heteropterygidae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Die drei Unterfamilien und deren Triben sind hier bis zur Gattungsebene dargestellt:[1][2]
- Dataminae Rehn, J. A. G. & Rehn, J. W. H., 1939
- Datamini Rehn, J. A. G. & Rehn, J. W. H., 1939
- Dares Stål, 1875
- Epidares Redtenbacher, 1906
- Hainanphasma Ho, 2013
- Microrestes Bresseel & Constant, 2020
- Orestes Redtenbacher, 1906
- Planispectrum Rehn, J. A. G. & Rehn, J. W. H., 1939
- Pylaemenes Stål, 1875
- Spinodares Bragg, 1998
- Datamini Rehn, J. A. G. & Rehn, J. W. H., 1939
- Heteropteryginae Kirby, 1896
- Heteropterygini Kirby, 1904
- Haaniella Kirby, 1904
- Heteropteryx Gray, G. R., 1835
- Heteropterygini Kirby, 1904
- Obriminae Brunner von Wattenwyl, 1893
(Syn. = Therameninae Karny, 1923)- Obrimini Brunner von Wattenwyl, 1893
(Syn. = Eubulidini Zompro, 2004)
(Syn. = Miroceramiini Zompro, 2004)
(Syn. = Tisamenini Hennemann, Conle, Brock & Seow-Choen, 2016)- Aretaon Rehn, J. A. G. & Rehn, J. W. H., 1939
- Brasidas Rehn, J. A. G. & Rehn, J. W. H., 1939
- Eubulides Stål, 1877
- Euobrimus Rehn, J. A. G. & Rehn, J. W. H., 1939
- Heterocopus Redtenbacher, 1906
- Mearnsiana Rehn, J. A. G. & Rehn, J. W. H., 1939
- Miroceramia Günther, 1934
- Obrimus Stål, 1875
- Pterobrimus Redtenbacher, 1906
- Stenobrimus Redtenbacher, 1906
- Sungaya Zompro, 1996
- Theramenes Stål, 1875
- Tisamenus Stål, 1875
- Trachyaretaon Rehn, J. A. G. & Rehn, J. W. H., 1939
- Hoplocloniini Bank et al., 2021
- Hoploclonia Stål, 1875
- Obrimini Brunner von Wattenwyl, 1893
Äußere Systematik
In der klassischen Taxonomie wurden die Heteropterygidae der umstrittenen Teilordnung der Areolatae zugeordnet. Bereits in dieser wurde sie mit den Familien Bacillidae und Anisacanthidae in der Überfamilie Bacilloidea geführt.[2] Wie genetische Untersuchungen zeigten handelt es sich bei den Vertretern der Bacilloidea nicht um eine einheitliche Klade. Lediglich die Verwandtschaft mit den Vertreter der madagassischen Anisacanthidae, die schon von Kirby 1904 bezüglich der Gattung Parectatosoma und 1906 von Redtenbacher ebenfalls an Parectatosoma und der neu errichteten Gattung Anisacantha gemutmaßt wurde, konnte bestätigt werden. Die Einordnung der Heteropterygidae in bisher genetisch untersuchte Vertreter verschiedener Taxa wird in folgendem Kladogramm dargestellt:[9]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Weblinks
Quellen
- Frank H. Hennemann, Oskar V. Conle, Paul D. Brock & Francis Seow-Choen: Zootaxa 4159 (1): Revision of the Oriental subfamiliy Heteropteryginae Kirby, 1896, with a re-arrangement of the family Heteropterygidae and the descriptions of five new species of Haaniella Kirby, 1904. (Phasmatodea: Areolatae: Heteropterygidae), Magnolia Press, Auckland, New Zealand 2016, ISSN 1175-5326
- Paul D. Brock, Thies H. Büscher & Edward W. Baker: Phasmida Species File Online. Version 5.0./5.0 (abgerufen am 21. März 2021)
- Phasmid Study Group Culture List (engl.)
- Oliver Zompro: Zwergformen der Phasmatodea - die Kleinsten unter den Riesen, Arthropoda 16 (3) November 2008, Sungaya-Verlag Kiel. ISSN 0943-7274
- Ingo Fritzsche: Stabschrecken - Carausius, Sipyloidea & Co., Natur und Tier Verlag, Münster 2007, ISBN 978-3-937285-84-9
- Christoph Seiler, Sven Bradler, Rainer Koch: Phasmiden – Pflege und Zucht von Gespenstschrecken, Stabschrecken und Wandelnden Blättern im Terrarium. bede, Ruhmannsfelden 2000, ISBN 3-933646-89-8
- Oliver Zompro: Stabschrecken, Gespenstschrecken, Wandelnde Blätter - Phasmidensystematik im Überblick I., Arthropoda 17 (1) April 2009, Sungaya-Verlag Kiel. ISSN 0943-7274
- James Abram Garfield Rehn & John W. H. Rehn: Proceedings of The Academy of Natural Sciences (Vol. 90, 1938), Philadelphia 1939, S. 389 ff.
- Sarah Bank, Thomas R. Buckley, Thies H. Büscher, Joachim Bresseel, Jérôme Constant, Mayk de Haan, Daniel Dittmar, Holger Dräger, Rafhia S. Kahar, Albert Kang, Bruno Kneubühler, Shelley Langton-Myers & Sven Bradler: Reconstructing the nonadaptive radiation of an ancient lineage of ground-dwelling stick insects (Phasmatodea: Heteropterygidae), Systematic Entomology (2021), DOI: 10.1111/syen.12472
- Phasmatodeaseite von Frank H. Hennemann, Oskar V. Conle, Bruno Kneubühler und Pablo Valero
- Karl Brunner von Wattenwyl: Annali del Museo Civico di Storia Naturale Giacomo Doria. Genova (2) 13 (33):101, 1893, S. 98
- William Forsell Kirby: A synonymic catalogue of Orthoptera. 1. Orthoptera Euplexoptera, Cursoria et Gressoria. (Forficulidae, Hemimeridae, Blattidae, Mantidae, Phasmidae) 1904, S. 396–400
- Joseph Redtenbacher: Die Insektenfamilie der Phasmiden. Vol. 1. Phasmidae Areolatae. Verlag Wilhelm Engelmann, Leipzig 1906, S. 36–57 & 162–172
- Klaus Günther: Über die taxonomische Gliederung und die geographische Verbreitung der Insektenordnung der Phasmatodea, Beiträge zur Entomologie, Band 3, 1953, Nr. 5, S. 541–563
- Oliver Zompro: Beiträge zur Kenntnis philippinischer Phasmiden I. Entomologische Zeitschrift (1996) 106 (4), S. 160–164.
- Oliver Zompro: Revision of the genera of the Areolatae, including the status of Timema and Agathemera (Insecta, Phasmatodea), Goecke & Evers, Keltern-Weiler 2004, S. 191–240, ISBN 978-3931374396