Heleosaurus
Heleosaurus ist eine ausgestorbene Gattung varanopider Synapsiden aus dem mittleren Perm von Südafrika. Fossilien des bis zu einen halben Meter langen Tieres stammen mit hoher Wahrscheinlichkeit aus der Tapinocephalus-Assemblage-Zone der Beaufort-Gruppe des südafrikanischen Karoo-Hauptbeckens.[1] Es existiert nur eine einzige Art, Heleosaurus scholtzi, die 1907 von dem südafrikanischen Paläontologen Robert Broom erstbeschrieben wurde.[2] Als einem der wenigen Vertreter der südafrikanischen Varanopiden kommt Heleosaurus eine wichtige Rolle bei der Bestimmung der Verwandtschaftsverhältnisse innerhalb des Monophylums der Varanopiden zu. Wie alle Varanopiden war Heleosaurus ein Fleischfresser.[1]
| Heleosaurus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
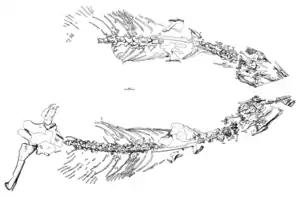
Zeichnung des Holotyps von Heleosaurus scholtzi | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Mittelperm | ||||||||||||
| 265,1 bis 259,9 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Heleosaurus | ||||||||||||
| Broom, 1907 | ||||||||||||
| Art | ||||||||||||
| ||||||||||||
Klassifikation
Heleosaurus scholtzi wurde im Jahr 1907 von Robert Broom als frühes diapsides Reptil beschrieben. Broom stellte die Art zusammen mit äußerlich ähnlichen Taxa zu den permischen und triassischen Eosuchia, von denen man annahm, dass sie den evolutionären Grundstock für lebende diapside Reptilien wie Brückenechsen, Eidechsen, Schlangen und Krokodile und deren fossile Vorfahren bildeten. Lange Zeit galt Heleosaurus als der Familie der Younginidae zugehörig. Dabei wurden die integrierte Panzerung, die s-förmige, diapsidenähnliche Krümmung des Femurs und die Zähnelung (Serration) der Zähne als Gründe für diese Einordnung angeführt.[3]
Nachdem bereits in den 1980er Jahren Zweifel an dieser Sicht aufgekommen waren, führte ein besseres Verständnis von Anatomie und Evolution der Varanopiden zu einer Neubewertung der taxonomischen Einordnung von Heleosaurus.[1]
Die Exemplare von Helosaurus stammen aus der Tapinocephalus-Assemblage-Zone der oberen Abrahamskraal-Formation des südafrikanischen Karoo-Beckens, die auf ein Alter von ca. 270 Millionen Jahren datiert wird.[1][4][5]
Beschreibung
Die einzige Art Heleosaurus scholtzi unterschied sich von anderen Varanopiden durch charakteristische Ornamentierung des angularen und surangularen Unterkieferknochens. Trotz der Serration der marginalen Zähne, die lippen- und zungenseitig (labiolingual) gestaucht erscheinen, waren die größten nicht verlängert wie bei anderen Varanopiden. Die bauchseitige Oberfläche jedes Rückenwirbels besaß einen querverlaufenden, breiten Kamm, welcher von einer längsverlaufenden Kerbe unterteilt wurde. Dieses Merkmal unterscheidet Heleosaurus von Elliotsmithia.[1]
Einzelne Exemplare von Heleosaurus besaßen eine Kopfrumpflänge von ca. 30 Zentimetern. Inklusive Schwanz maß Heleosaurus ungefähr 50 Zentimeter. Die Schnauze war niedrig und angeschrägt, das Jochbein (Os jugale) und das Os quadratojugale waren mit kleinen, gequetschten Höckern oder Knöllchen besetzt. Dieses Merkmal teilte Heleosaurus mit der nahe verwandten Gattung Mesenosaurus. Wie bei Mesenosaurus saß ein besonders großes Tuberkel auf der unterseitigen Kante des Os jugale auf der Höhe des postorbitalen Fortsatzes. Das Os quadratojugale war verlängert und reichte bis weit unter das Temporalfenster, so dass es beinahe die Maxilla berührte. Eine schmale Einkerbung entlang dem Saum für die Anheftung des Schuppenbeins. Dies deutet darauf hin, dass das Schuppenbein vorn (anterior) wie bei den meisten anderen kleinen Varanopiden bis weit unter das seitliche (laterale) Temporalfenster reichte.[1]
Das Pflugscharbein war lang und schmal und besaß eine einzelne Reihe kleiner Zähne an seiner medialen Kante. Hierin weist Heleosaurus ebenfalls starke Ähnlichkeiten mit Mesenosaurus auf. Auch die Anordnung der beiden Knochen des Gaumenbeins, welches sich in der Nähe der internen Naris gabelt, ähnelt derjenigen von Mesenosaurus. Das Gaumenbein reichte somit zu weit nach hinten (posterior), als dass eine zweite Temporalöffnung wie bei den Diapsiden Platz gefunden hätte. Dieser Befund unterstützt die Annahme, dass Heleosaurus zu den Varanopiden gehört, also ein Synapside war.[1]
Heleosaurus besaß schlanke Rippen, die nur durch ein Gelenk mit den Rückenwirbeln verbunden waren, also nur einen Rippenkopf besaßen. Die rumpfwärts (proximal) gelegenen Enden der Rippen waren ebenfalls schmal. Die Köpfe der präsakralen Rippen waren dreieckig und verbreitert. Ähnliche Merkmale wiesen auch Mycterosaurus und Archaeovenator auf. Unterhalb der Rippen sind sehr schmale Bauchrippen (Gastralia) zu erkennen, die nur rund ein Drittel der Breite der Rippen aufweisen und circa 20 Millimeter lang waren.[4]
Ein wichtiges Merkmal von Heleosaurus waren Osteoderme (Hautknochenplatten), die rückenseits entlang der Wirbelsäule liegen. Die Knochenplatten hatten eine viereckige oder leicht gerundete Form und waren in Reihen mit bis zu fünf Plättchen angeordnet. Die Gattung Elliotsmithia wies ähnliche Osteoderme auf. Der Durchmesser der Knochenplättchen nimmt von der Mitte nach den Seiten hin ab.[4] Auch permische Diapsiden wie Youngina besaßen eine osteoderme Panzerung, was ein Grund für die ursprüngliche Einordnung von Heleosaurus als Diapside war.[1]
Die Morphologie des Femur weist starke Ähnlichkeiten mit derjenigen von Mycterosaurus and Varanops auf. Der Knochen ist schlank, länglich und besitzt eine s-förmige Kurvatur. Die Beschaffenheit des Schaftes und des Rollhügels unterscheidet sich stark von der von Youngina.[1]
Paläoökologie
Eine Aggregation von Heleosaurusfossilien weist darauf hin, dass Heleosaurus Brutpflege betrieb. So fand man in Südafrika die gut erhaltenen Überreste von fünf Individuen – einem ausgewachsenes Tier und vier Jungtieren gleicher Größe – eng beieinander. Aufgrund des guten und fast vollständigen Erhaltungszustandes sowie der engen, in eine Richtung ausgerichtete Anordnung der Fossilien schließen Botha-Brink und Modesto eine Entstehung der fossilen Aggregation durch Anschwemmung oder andere Transportprozesse nach dem Tod der Tiere aus und vermuten, dass die Individuen gemeinsam in einem Bau oder Unterschlupf starben und fossilierten. Diese These wird weiterhin durch die Tatsache gestützt, dass Heleosaurus ein sehr seltener Vertreter der mittelpermischen Fauna Südafrikas war, eine zufällige Ansammlung der Kadaver also als unwahrscheinlich gelten kann.[5]
Dies wäre der älteste Nachweis von elterlicher Fürsorge bei Pelycosauriern und Amnioten überhaupt, ca. 140 Millionen Jahre, bevor dies bei den Dinosauriergattungen Psittacosaurus und Oryctodromeus nachgewiesen werden konnte. Früheste Hinweise auf soziales Verhalten finden sich erst bei Überresten von Tropidostoma und Diictodon aus dem Oberperm. Brutpflege bedeutete einen enormen Überlebensvorteil in einer paläozoischen Umwelt, die von Therapsiden dominiert wurde. Botha-Brink und Modesto sehen in der Brutpflege von Heleosaurus Parallelen zu der australischen Gattung Egernia aus der Familie der Skinke.[5]
Einzelnachweise
- R. R. Reisz, S. P. Modesto: Heleosaurus scholtzi from the Permian of South Africa: a varanopid synapsid, not a diapsid reptile. In: Journal of Vertebrate Paleontology. 27 (3), 2007, S. 734–739.
- R. Broom: On some new fossil reptiles from the Karroo beds of Victoria West, South Africa. In: Transactions of the South African Philosophical Society. 18, 1907, S. 31–42.
- R. L. Carroll: Eosuchians and the origin of archosaurs. In: C. S. Churcher (Hrsg.): Athlon: Essays on Paleontology in Honour of Loris Shano Russell. Miscellaneous Publications of the Royal Ontario Museum, Toronto 1976, S. 58–79.
- J. Botha-Brink, S. P. Modesto: Anatomy and Relationships of the Middle Permian Varanopid Heleosaurus scholtzi Based on a Social Aggregation from the Karoo Basin of South Africa. In: Journal of Vertebrate Paleontology. 29 (2), 2009, S. 389–400.
- Jennifer Botha-Brink, Sean P. Modesto: A Mixed-Age Classed 'Pelycosaur' Aggregation from South Africa: Earliest Evidence of Parental Care in Amniotes? In: Proceedings: Biological Sciences. Vol. 274, No. 1627, 2007, S. 2829–2834.