Os quadratojugale
Das Os quadratojugale (kurz einfach Quadratojugale) ist ein paariger Schädelknochen der Osteognathostomata (Knochenfische einschließlich der Landwirbeltiere). Es handelt sich dabei um einen dermalen Deckknochen der postorbitalen, unteren Schädelseitenwand, der, sofern vorhanden, stets in enger räumlicher Beziehung mit dem Quadratum, einem Ersatzknochen des Palatoquadratums und Teil des primären Kiefergelenks, steht. Das Quadratojugale wird in zoologischen und paläontologischen Schädelzeichnungen meist mit dem Kürzel Qj oder qj gekennzeichnet.
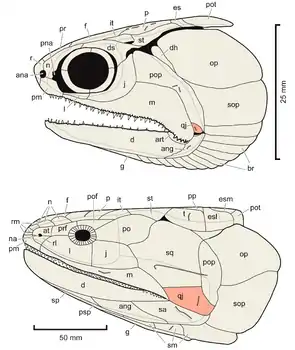
Das Vorhandensein eines Quadratojugale gilt als Merkmal im Grundbauplan der Osteognathostomata und dieser Knochen findet sich sowohl bei ursprünglichen Vertretern der Strahlenflosser (Actinopterygii) als auch bei ursprünglichen Vertretern der Fleischflosser (Sarcopterygii).[1]
Strahlenflosser
Bei vielen ursprünglichen Strahlenflossern, wie den „Palaeonisciformes“ Cheirolepis, Mimipiscis (ehem. Mimia) oder Myothomasia, ist das Quadratojugale ein kleines Knochenplättchen, das dem Quadratum seitlich (lateral) aufsitzt und zusammen mit der hinteren Partie des Maxillare und dem Präoperculum eine steife Verkleidung der Schädelseitenwand bildet.
Bei den Stören (Acipenseridae) mit ihren stark abgeleiteten und größtenteils knorpeligen Schädeln ist das Quadratojugale ein kleiner Knochen oberhalb des hinteren Endes des Maxillare. Dieser Knochen ist aber auch als Supramaxillare interpretiert worden.[1]
Bei einigen basalen Neopterygiern, wie den ausgestorbenen Macrosemiiden oder den Knochenhechten (Lepisosteidae) ist das Quadratojugale ein schmaler, manchmal teilweise mit dem Quadratum verwachsener Knochensplint, der an der Aufhängung des Kiefers (Suspensorium) beteiligt ist.[2] Bei den Teleosteern ist das Quadratojugale nicht mehr als separater Knochen vorhanden und vermutlich komplett mit dem hinteren äußeren Rand des Quadratums verschmolzen.[2] Allerdings haben ontogenetische Studien diese Aussage nicht in jedem Fall bestätigt.[1]
Generell gilt, dass bislang nicht vollständig geklärt ist, ob die bei den verschiedenen Strahlenflossertaxa als Quadratojugale identifizierten Knochen wirklich alle einander homolog sind.
Fleischflosser
Fischartige Vertreter
Das Quadratojugale liegt bei vielen frühen fischartigen Fleischflossern, ähnlich wie bei den frühen Strahlenflossern, zwischen Maxillare und Präoperculum, ist aber ein relativ großer, flächiger Knochen. Sein oberer (dorsaler) Rand steht mit dem Squamosum in Kontakt. Die Lungenfische (Dipnoi) besitzen jedoch stark modifizierte Schädel, für die aufgrund der schwierigen Homologisierbarkeit der Knochen eine gänzlich abweichende Nomenklatur in Gebrauch ist. Bei frühen Vertretern der Gruppe, wie Dipterus aus dem mittleren Devon, wird der Circumorbitalknochen Nr. 10 als dem Quadratojugale homologes Element in Betracht gezogen.[1] Die Schädelanatomie von Latimeria, dem einzigen lebenden Vertreter der Quastenflosser (Actinistia), weicht deutlich von jener seiner frühen Verwandten ab. Auch bei ihm ist kein Quadratojugale mehr vorhanden.
Landwirbeltiere

Durch die vollständige Reduktion der Opercularserie (Knochen des Kiemendeckels) und einer generellen Verkürzung des postorbitalen Schädels ist das Quadratojugale im Grundbauplan der Tetrapoden am hinteren Rand der Schädelseitenwand beteiligt und steht zudem an seinem vorderen (rostralen) Ende mit dem Jugale in Kontakt. Allerdings geht durch Beteiligung des Jugale am unteren Rand des Craniums (Schädel ohne Unterkiefer) der Kontakt zum Maxillare verloren – ist bei sehr frühen Formen wie z. B. Ichthyostega aber noch vorhanden. Die Schädel der ursprünglicheren Tetrapoden (u. a. Labyrinthodontier bzw. Stegocephalen) weisen prinzipiell alle immer ein Quadratojugale auf. Bei den modernen Amphibien (Lissamphibia) und bei den Amnioten ist es zum Teil reduziert.
Bei den Froschlurchen (Anura) steht das Quadratojugale aufgrund der Reduktion des Jugale wieder mit dem Maxillare in Kontakt. Bei den meisten Schwanzlurchen (Urodela) ist es im Zuge der Auflösung der Wangenregion nicht mehr vorhanden. Der sehr kompakte Schädel der Blindwühlen (Gymnophiona) weist kein separates Quadratojugale auf.
Bei den Synapsiden, in deren Entwicklungsgeschichte generell ein Trend zur Vereinfachung und Kompaktion des Schädels besteht, wird das Quadratojugale bereits früh komplett reduziert. Nur bei einigen „Pelycosaurier“-Gruppen ist es vorhanden und an der Umgrenzung des Temporalfensters beteiligt. Alle Therapsiden – und mit ihnen die Säugetiere – besitzen kein Quadratojugale mehr.
Bei den Pareiasauriern und einigen Procolophonoiden, permischen bzw. permo-triassischen Parareptilien, bildet das Quadratojugale einen charakteristischen, seitlich nach unten weisenden, manchmal bestachelten Auswuchs.
Im Grundbauplan der Diapsiden ist es an der Umgrenzung des unteren Temporalfensters beteiligt. Hierbei besteht allerdings ein Unterschied zwischen frühen und moderneren Vertretern mit „echtem“ diapsiden Schädel, d. h. mit geschlossenem unteren Temporalbogen: Während bei frühen Diapsiden das Quadratojugale in etwa zur Hälfte am unteren Temporalbogen beteiligt ist, wird der untere Temporalbogen z. B. bei den Archosauriern zu einem großen Teil vom Jugale gebildet. Dies ist ein Grund für die Annahme, dass der untere Temporalbogen u. a. der Archosaurier eine sekundäre Neubildung ist.[3]
Bei den Schuppenkriechtieren (Squamata) ist, vermutlich im Zuge der Ausbildung einer speziellen Form der Schädelkinetik (mesokinetischer Schädel in Verbindung mit Streptostylie), der untere Temporalbogen komplett reduziert und kein separates Quadratojugale mehr vorhanden.
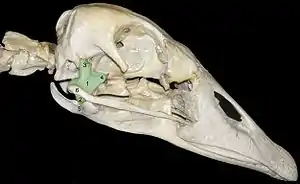
Die Vögel (Aves) haben den ursprünglich „triapsiden“ Archosaurierschädel stark modifiziert und ebenfalls eine spezielle Form der Schädelkinetik entwickelt (prokinetischer Schädel). Im Gegensatz zu den Squamaten ist hierbei allerdings das Quadratojugale mit involviert. Es ist mit dem Jugale zu einem stabförmigen Knochen verschmolzen, der zusammen mit einem nach hinten gerichteten Fortsatz des Maxillare (Processus jugalis) einen Knochensteg bildet, der als Arcus jugalis (auch Arcus zygomaticus) bezeichnet wird. Dieser ist Teil des „Schubstangensystems“ des Vogelschädels: Er ist an seinem hinteren Ende, d. h. über das Quadratojugale, gelenkig mit dem pendelartig beweglichen Quadratum verbunden. Dies bewirkt beim Öffnen des Schnabels eine aktive Aufwärtsbewegung des Oberschnabels, indem die Bewegung des beim Absenken des Unterschnabels nach vorn schwingenden Quadratums pleuelartig auf den hinteren unteren Rand des Oberschnabels weitergeleitet wird.
Literatur
- Robert L. Carroll: Vertebrate Paleontology and Evolution. W. H. Freeman and Co., New York 1988.
- Milton Hildebrand, George E. Goslow: Vergleichende und funktionelle Anatomie der Wirbeltiere. Springer, 2004, ISBN 3-540-00757-1.
- Ulrich Lehmann: Paläontologisches Wörterbuch. 4. Auflage. Ferdinand Enke Verlag, Stuttgart 1996, ISBN 3-432-83574-4.
- Gerhard Mickoleit: Phylogenetische Systematik der Wirbeltiere. Verlag Dr. Friedrich Pfeil, München 2004, ISBN 3-89937-044-9.
- Franz-Viktor Salomon, Maria-Elisabeth Krautwald-Junghanns: Anatomie der Vögel. In: Salomon u. a. (Hrsg.): Anatomie für die Tiermedizin. 2. erw. Auflage. Enke Verlag, Stuttgart 2008, ISBN 978-3-8304-1075-1, S. 754–814.
Einzelnachweise
- Gloria Arratia, Hans-Peter Schultze: Palatoquadrate and its Ossifications: Development and Homology within Osteichthyans. In: Journal of Morphology. Bd. 208, Nr. 1, 1991, S. 1–81, doi:10.1002/jmor.1052080102 (siehe speziell S. 2 und S. 64 ff.)
- Adriana López-Arbarello: Phylogenetic Interrelationships of Ginglymodian Fishes (Actinopterygii: Neopterygii). In: PLoS ONE. Band 7, Nr. 7, 2012, S. e39370. doi:10.1371/journal.pone.0039370
- Johannes Müller: Early loss and multiple return of the lower temporal arcade in diapsid reptiles. In: Naturwissenschaften. Band 90, Nr. 10, 2003, S. 473–476. doi:10.1007/s00114-003-0461-0