Dinophysis acuta
Dinophysis acuta ist eine Art (Spezies) von Dinoflagellaten, die zur Gattung Dinophysis gehört. Dieser Einzeller ist einer der wenigen ungewöhnlichen photosynthetischen Protisten, die durch Endosymbiose Plastiden von Algen erwerben. Durch die Bildung massiver Blüten, insbesondere im Spätsommer und Frühjahr, verursacht er rote Tiden (englisch red tides). D. acuta produziert giftige Substanzen (Toxine), und die roten Tiden verursachen weit verbreitete Infektionen von Meeresfrüchten, insbesondere von Krabben und Muscheln. Wenn infizierte Tiere dann verzehrt werden, kommt es zu schweren Durchfällen (Diarrhoe); dieses klinische Symptom wird als diarrhöische Schalentiervergiftung bezeichnet (en. diarrhetic shellfish poisoning, DSP).[1]
| Dinophysis acuta | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Dinophysis acuta | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Dinophysis acuta | ||||||||||||
| Ehrenberg, 1839 |
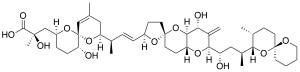
Die wichtigsten chemischen Toxine wurden 2006 als Okadasäure und Pectenotoxine[2][3][4][5] identifiziert.[6][7][8] Sie können in ihren Fressfeinden nicht tödliche oder auch tödliche Mengen an Toxinen produzieren, die auch beim Menschen Vergiftungen hervorrufen können.
Dinophysis acuta ist Einzeller des Jahres 2020.[9]
Beschreibung
Morphologie
Dinophysis acuta ist ein mariner einzelliger Protist und der größte in der Gattung Dinophysis. Es handelt sich um eine gepanzerte Art mit einer ausgeprägten Körperhülle, die Theca genannt wird. Der Körper ist seitlich zusammengedrückt und die Theca besteht aus einer kleinen, kappenartigen Epitheca und einer viel größeren Hypotheca. Sie hat einen doppelten Kragen (das so genannte Cingulum) um den oberen Teil der Zelle und einen weiteren Flügel (den so genannten Sulcus), der vertikal in der Zelle verläuft. Sie hat eine längliche Form mit einem fast vollständig abgerundeten hinteren Ende, das jedoch an der Spitze leicht spitz ist. Die Größe reicht von 54 bis 94 µm in der Länge und 43 bis 60 µm in der dorso-ventralen Breite, wobei der breiteste Bereich unterhalb der Mitte liegt. Die kleine Epitheca besteht aus vier Platten. Sie ist niedrig, flach oder schwach konvex und in der Seitenansicht nicht sichtbar, was ein gutes Erkennungsmerkmal ist. Der Sulcus besteht aus mehreren unregelmäßig geformten Platten und enthält die Geißelpore. Die Hypotheca besteht aus vier großen Platten, die den größten Teil der Zelle ausmachen. Die vorderen zwei Drittel der Hypotheca haben konvexe Ränder, während das hintere Drittel ein breites asymmetrisches Dreieck mit einem geraden dorsalen Rand und gelegentlich einem leicht konkaven ventralen Rand bildet.[10][11][12]
Fortpflanzung
Die Fortpflanzung erfolgt gewöhnlich durch einfache binäre Spaltung. Lange Zeit glaubte man, dass die Dinophysis-Arten keinen Sexualzyklus (mit geschlechtlicher Fortpflanzung) hat. Inzwischen ist jedoch klar, dass sich bei D. acuminata und D. acuta Gametenzellen bilden können. Dies wurde festgestellt, als sich kleine, kugelförmige Zellen innerhalb größerer Zellen zu bilden schienen.[13]
Kleptoplastidie
Das Ungewöhnlichste an der Zellstruktur von D. acuta wie auch von D. acuminata sind zahlreiche rötlich-gelbe (phycobilinhaltige) Chloroplasten, die von ihrer Beute, Wimpertierchen der Gattung Mesodinium (M. rubrum, synonym Myrionecta rubra) stammen, die sie ihrerseits von Cryptophyceen erworben hat (Kleptoplastidie).[14][15]
Photosynthetische Arten der Gattung Dinophysis sind obligate Mixotrophe, die zum Überleben und Wachsen Licht, Nährstoffe und lebende Beute benötigen.[14]
Der bereits früher entdeckte Fressmechanismus von Phalacroma rotundatum und Dinophysis hastata, die sich von dem Wimpertierchen Tiarina fusus ernähren, ist eine Art Phagozytose (Myzozytose), bei der der Inhalt der Beute durch einen Pedunkel (englisch feeding peduncle) in die Zelle des Räuberzelle befördert wird. Eine ähnliche Struktur wird von D. acuminata und D. acuta benutzt, um sich von M. rubrum zu ernähren.[14]
Anders als früher gedacht, besitzen die photosynthetischen Arten der Gattung Dinophysis aber keine eigenen Chloroplasten. Stattdessen behalten sie die Kleptoplastiden ihrer Wimpertierchens-Beute M. rubrum vorübergehend zurück. Diese Wimpertierchen ernähren sich von Cryptophyceen (insbesondere aus dem Teleaulax/Plagioselmis/Geminigera-Komplex[16]), wobei sie deren Chloroplasten als Kleptoplastiden (und auch Mitochondrien) in sich aufnehmen. Man geht davon aus, dass unterschiedliche Plastiden bei Dinophysis letztlich von unterschiedlichen Cryptophyceen-Quellen stammen. Feldstudien haben bestätigt, dass Teleaulax-ähnliche Arten wie T. amphioxeia die häufigste Quelle von Plastiden in Dinophysis sind. Es kann aber die Möglichkeit nicht ausgeschlossen werden, dass auch andere Kleptoplastid-tragende Wimpertierchen wie Cyrtostrombidium, Laboea, Strombidium und/oder Tontonia neben Mesodinium als Überträger von Teleaulax-ähnlichen Plastiden auf Dinophysis in Frage kommen. Dafür spricht die Entdeckung von Plastiden aus mehreren Algenquellen in Dinophysis-Arten aus koreanischen Gewässern.[14][17][15][18]
Toxizität
Die ersten Fälle von diarrhöischen Schalentiervergiftungen (en. diarrhetic shellfish poisoning, DSP) durch D. acuta wurden 1972 in Peru festgestellt, der wissenschaftlichen Gemeinschaft jedoch erst 1991 gemeldet.[19] Es handelt sich um eine sehr milde Form von Vergiftung durch Meeresfrüchte, die sich durch schweren Durchfall bemerkbar macht.[1] Die ersten Toxine, die aus dieser Art isoliert wurden, waren im Jahr 2003 die Pectenotoxine PTX-2[4] und PTX-11[5] aus Exemplaren, die an der Westküste der Südinsel Neuseelands gesammelt wurden,[20] und PTX-12[2] unabhängig davon in Skjer im Sognefjord (auf dem Südarm Nærøyfjord) in Norwegen.[21] Im Jahr 2004 wurde das Vorhandensein von Okadasäureestern gemeldet.[22] 2006 wurden weitere dieser Verbindungen identifiziert und ihre Bedeutung als ursächliche Faktoren von DSP entdeckt.[6][7][8]
Einzelnachweise
- David Arieti, Jakob Nieva, Randolf Swiller: Prognosis Disaster: The Environment, Climate Change, Human Influences, Vectors, Disease and the Possible End of Humanity?. Author House, 2011, ISBN 9781456731496, S. 404.
- Zhaoxin Li, Guo Mengmeng, Yang Shouguo, Wang Qingyin, Tan Zhijun: Investigation of Pectenotoxin Profiles in the Yellow Sea (China) Using a Passive Sampling Technique, in: MDPI Marine Drugs, Band 8, Nr. 4, S. 1263–1272, Special Issue Algal Toxins, 15. April 2010, doi:10.3390/md8041263
- Pectenotoxin-1, auf: Pubchem
- pectenotoxin 2, auf Chemical Book
- PTX-11 -Lösung, auf: Merck Sigma-Aldrich
- Christopher Owen Miles, Alistair L. Wilkins, Allan D. Hawkes, Dwayne J. Jensen, Janine M. Cooney, Kristofer Larsen, Dirk Petersen, Frode Rise, Veronica Beuzenberg, A. Lincoln MacKenzie: Isolation and identification of a cis-C8-diol-ester of okadaic acid from Dinophysis acuta in New Zealand. In: Toxicon. 48, Nr. 2, August 2006, S. 195–203. doi:10.1016/j.toxicon.2006.04.018. PMID 16784765.
- Christopher Owen Miles, Alistair L. Wilkins, Allan D. Hawkes, Dwayne J. Jensen, Andrew I. Selwood, Veronica Beuzenberg, A. Lincoln MacKenzie, Janine M. Cooney, Patrick T. Holland: Isolation and identification of pectenotoxins-13 and -14 from Dinophysis acuta in New Zealand. In: Toxicon. 48, Nr. 2, August 2006, S. 152–159. doi:10.1016/j.toxicon.2006.04.005. PMID 16828828.
- Toshiyuki Suzuki, John A. Walter, Patricia LeBlanc, Shawna MacKinnon, Christopher O. Miles, Alistair L. Wilkins, Rex Munday, Veronica Beuzenberg, A. Lincoln MacKenzie, Dwayne J. Jensen, Janine M. Cooney, Michael A. Quilliam: Identification of pectenotoxin-11 as 34S-hydroxypectenotoxin-2, a new pectenotoxin analogue in the toxic dinoflagellate Dinophysis acuta from New Zealand. In: Chemical Research in Toxicology. 19, Nr. 2, 2006, S. 310–318. doi:10.1021/tx050240y. PMID 16485908.
- Einzeller des Jahres 2020: Dinophysis acuta, Deutsche Gesellschaft für Protozoologie
- Identifying harmful marine dinoflagellates. Department of Botany, Smithsonian Institution. Abgerufen am 25. August 2014.
- P. Lassus, M. Seguel, P. Truquet: Morphological study of atypical Dinophysis acuta Ehrenberg from Chilean coastal waters by a digital pattern-recognition system. In: Botanica Marina. 41, Nr. 1–6, 1998, S. 435–441. doi:10.1515/botm.1998.41.1-6.435.
- D. A. Burns, J. S. Mitchell: Dinoflagellates of the genus Dinophysis Ehrenberg from New Zealand coastal waters. In: New Zealand Journal of Marine and Freshwater Research. 16, Nr. 3–4, 1982, S. 289–298. doi:10.1080/00288330.1982.9515972.
- Beatriz Reguera, Lourdes Velo-Suárez, Robin Raine, Myung Gil Park: Harmful Dinophysis species: A review. Harmful Algae, in: Harmful Algae, Band 14, Februar 2012, S. 87–106, doi:10.1016/j.hal.2011.10.016
- María García-Portela, Beatriz Reguera, Manoella Sibat, Andreas Altenburger, Francisco Rodríguez, Philipp Hess: Metabolomic Profiles of Dinophysis acuminata and Dinophysis acuta Using Non-Targeted High-Resolution Mass Spectrometry: Effect of Nutritional Status and Prey, in: MDPI Marine Drugs, Band 16, Nr. 15, 143, 26. April 2018, doi:10.3390/md16050143, PMID 29701702, PMC 5982093 (freier Volltext).
- A. Mitra: Meeresbiologie – Das Beste aus zwei Welten., Spektrum der Wissenschaft, 20. März 2019, S. 54–57.
- Cryptomonads. In: Tree of Life Project. 2. April 2010. Abgerufen am 7. Dezember 2021.
- S. Minnhagen, W. F. Carvalho, P. S. Salomon, S. Janson: Chloroplast DNA content in Dinophysis (Dinophyceae) from different cell cycle stages is consistent with kleptoplasty. In: Environ. Microbiol. Band 10, Nr. 9, September 2008, S. 2411–2417, doi:10.1111/j.1462-2920.2008.01666.x, PMID 18518896.
- G. Nishitani, S. Nagai, K. Baba, S. Kiyokawa, Y. Kosaka, K. Miyamura, T. Nishikawa, K. Sakurada, A. Shinada, T. Kamiyama: High-level congruence of Myrionecta rubra prey and Dinophysis species plastid identities as revealed by genetic analyses of isolates from Japanese coastal waters. In: Applied and Environmental Microbiology. Band 76, Nr. 9, 2010, S. 2791–2798, doi:10.1128/AEM.02566-09, PMID 20305031, PMC 2863437 (freier Volltext).
- G. Lembeye, T. Yasumoto, J. Zhao, R. Fernandez; Theodore J. Smayda, Yuzuru Shimizu (Hrsg.): DSP outbreak in Chilean Fiords. In: Toxic phytoplankton blooms in the sea. Elsevier Science Publishers, Amsterdam 1993, ISBN 9780444897190, S. 525–529.
- Toshiyuki Suzuki, Veronica Beuzenberg, A. Lincoln Mackenzie, Michael A. Quilliam: Liquid chromatography-mass spectrometry of spiroketal stereoisomers of pectenotoxins and the analysis of novel pectenotoxin isomers in the toxic dinoflagellate Dinophysis acuta from New Zealand. In: Journal of Chromatography A. 992, Nr. 1–2, 2003, S. 141–50. doi:10.1016/s0021-9673(03)00324-8. PMID 12735470.
- Christopher O. Miles, Alistair L. Wilkins, Ingunn A. Samdal, Morten Sandvik, Dirk Petersen, Michael A. Quilliam, Lars J. Naustvoll, Thomas Rundberget, Trine Torgersen, Peter Hovgaard, Dwayne J. Jensen, Janine M. Cooney: A novel pectenotoxin, PTX-12, in Dinophysis spp. and shellfish from Norway. In: Chemical Research in Toxicology. 17, Nr. 11, 2003, S. 1423–1433. doi:10.1021/tx049870a. PMID 15540940.
- Toshiyuki Suzuki, Veronica Beuzenberg, A. Lincoln Mackenzie, Michael A. Quilliam: Discovery of okadaic acid esters in the toxic dinoflagellate Dinophysis acuta from New Zealand using liquid chromatography/tandem mass spectrometry. In: Rapid Communications in Mass Spectrometry. 18, Nr. 10, 2004, S. 1131–1138. bibcode:2004RCMS...18.1131S. doi:10.1002/rcm.1455. PMID 15150838.