Uintatherium
Uintatherium ist ein ausgestorbenes Säugetier aus der Gruppe der Dinocerata, das im Mittleren Eozän vor 48 bis 43 Millionen Jahren in Nordamerika und Ostasien lebte. Es ähnelte den heutigen Nashörnern und stellte eine der ersten Großformen von Säugetieren nach dem Aussterben der Dinosaurier dar. Charakteristisch war sein Schädel, der drei aus Knochen gebildete Hornpaare aufwies, ebenso wie der säbelartige Eckzahn des Oberkiefers. Uintatherium ernährte sich von weicher Pflanzennahrung. Einen gewissen Bekanntheitsgrad erlangte die Gattung durch ihre Entdeckungsgeschichte, die 1872 begann und in deren Verlauf sie von drei unterschiedlichen Forschern nahezu zeitgleich unter verschiedenen Namen beschrieben wurde.
| Uintatherium | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelett von Uintatherium | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Unteres?/Mittleres bis Oberes Eozän (Oberes Bridgerium bis Mittleres Uintum) | ||||||||||||
| 48,8 bis 42,9 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Uintatherium | ||||||||||||
| Leidy, 1872 | ||||||||||||
| Arten | ||||||||||||
| ||||||||||||
Merkmale
Uintatherium war neben Eobasileus einer der größten Vertreter der Uintatheriidae, ausgestorbenen frühen Huftieren. Allgemein hatte er einen massigen, eher tonnen- oder walzenförmig wirkenden Körperbau und erreichte laut einiger rekonstruierter Museumsskelette eine Kopf-Rumpf-Länge von 3,1 bis 3,2 m, wobei die Schulterhöhe rund 1,6 m maß. Das Gewicht wird auf 1500 bis 2000 kg geschätzt, wobei er damit etwas leichter als Eobasileus war. Die stämmigen, elefantenähnlichen Beine endeten in jeweils fünf Strahlen mit Hufen.[1][2]
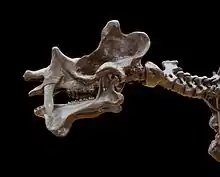
Der Schädel erreichte eine Länge von 69 bis 85 cm und war sehr flach, zusätzlich aber auch sehr breit. Von anderen großen Uintatherien unterschied er sich durch die allgemeinen Proportionen; so besaß Uintatherium ein eher kurzes Rostrum, aber einen längeren Hinterschädel.[2] Das Hinterhauptsbein war ausgezogen und verlängert, in der Ansicht von hinten besaß es eine markant rechteckige bis quadratische Form. Das Nasenbein lag deutlich oberhalb des Mittelkieferknochens und war häufig leicht nach unten gerichtet, gegenüber dem Stirnbein hatte es eine markant ausgedehnte Länge. Die Orbita erreichte recht große Ausmaße. Herausragendes Merkmal von Uintatherium waren drei Hornpaare auf dem Schädel, die durch knöcherne Auswüchse gebildet wurden. Das vorderste Hornpaar saß auf der vorderen Spitze des Nasenbeins und war sehr niedrig, es bestand nur aus zwei leichten Knochenerhebungen. Das mittlere, häufig sehr langgestreckte Paar war mit dem hinteren Ende des Oberkieferknochens verbunden und ragte über das zwischen Eckzahn und hinterer Bezahnung liegende Diastema. Das hinterste und ebenfalls recht hohe Hornpaar setzte dagegen an den Scheitelbeinen an.[3][2][1]
Der Unterkiefer war sehr robust, mit einer Länge von 49 bis 54 cm, aber im Vergleich zum Schädel relativ kurz.[4] An der Unterseite befanden sich im vorderen Bereich teils massiv ausgeprägte, nach unten ragende und konvex geformte Knochenauswüchse. Der Knochenkörper erreichte bis zu 12,5 cm Höhe, charakteristischerweise standen die Gelenkenden deutlich weit auseinander. Die Symphyse war kräftig und bis zu 15 cm lang. Das Gebiss zeichnete sich durch einige markante Reduktionen aus, so waren die oberen Schneidezähne generell nicht ausgebildet. Die Zahnformel lautete dementsprechend: . Die unteren Schneidezähne besaßen eine nur sehr kleine Form, wiesen aber zwei hintereinander liegende konische Höcker auf. Auch der untere Eckzahn war eher klein und ähnelte den Schneidezähnen. Der obere Eckzahn dagegen erreichte riesige Ausmaße. Er war insgesamt säbelartig flach, mit einer Länge von bis zu 6,6 cm und einer Breite bis zu 2,9 cm an der Basis, und nach hinten gekrümmt. Seine Gesamtlänge erreichte 18 cm, über die äußere Krümmung gemessen sogar 30 cm, so dass er deutlich über den Unterkiefer hinausragte. Dabei dienten die unteren Knochenauswüchse am Unterkiefer zum Schutz dieses Zahns. Die hintere Bezahnung war durch ein großes, bis 11 cm messendes Diastema von der vorderen getrennt. Pro Kieferbogen waren drei Prämolaren ausgebildet, nur sehr selten kam der vorderste vor, der dann sehr klein war und direkt an den Eckzahn noch vor dem Diastema ansetzte. Prämolaren und Molaren waren vergleichsweise klein und niederkronig (brachyodont), der letzte Backenzahn wurde maximal 4,5 cm lang. Die Kauoberfläche war durch zwei querstehende Zahnschmelzfalten geprägt (bilophodont), charakteristisch war die Ausbildung einer V-förmig gestalteten Schmelzfalte auf den Oberkieferzähnen, die typisch für frühe Säugetiere ist.[3][2][1][5]

Das postcraniale Skelett ist weitgehend vollständig bekannt. Vor allem die Wirbelsäule ähnelt jener der Rüsseltiere, die Halswirbel waren aber vergleichsweise länger, das Kreuzbein bestand generell aus vier Wirbeln. Der Oberarmknochen wurde bis zu 60 cm lang, die Ulna bis zu 58 cm. Das Becken maß über die äußersten Enden bis zu 120 cm und war dadurch sehr breit und eher niedrig. Der Oberschenkelknochen erreichte 71 cm Länge und besaß charakteristischerweise keinen dritten Trochanter. Das Schienbein maß bis zu 50 cm. Alle Gliedmaßen endeten in fünf Strahlen. Dabei waren sowohl die Vorder- als auch die Hinterfüße eher breit und kurz gestaltet, auch hier ähnelte der Aufbau stark den Rüsseltieren.[3][2][1]
Fossilfunde

Der überwiegende Teil der Fossilfunde stammt aus dem US-Bundesstaat Wyoming und kam im oberen Bereich der Bridger-Formation (Abschnitte C und D) zu Tage, welche dem Mittleren Eozän angehört, allerdings sind auch Funde aus dem Beginn des Oberen Eozän bekannt.[6] Die Bridger-Formation ist weitläufig im Bridger- und Washakie-Becken im südlichen Wyoming verbreitet und wird bis zu 200 m mächtig. Die ersten Funde gehen bis in die 1870er und 1880er Jahre zurück, allein in dieser Forschungsperiode, die in die unter forschungswissenschaftlichen Gesichtspunkten wichtige Zeit der Cope-Marsh-Fehde fallen, wurden unzählige Fossilreste gefunden, darunter rund 20 Schädel oder Schädelfragmente und Hunderte Knochen des postcranialen Skelettes, unter anderem mit vollständigem Vorder- und Hinterfüßen. Von 1893 bis 1906 führte das American Museum of Natural History mehrere Expeditionen durch, bei denen unter anderem das Washakie-Becken, aber auch das Uinta-Becken im angrenzenden Bundesstaat Utah Untersuchungsschwerpunkte darstellten. Funde aus letzterer Lokalität stammen aus der etwa gleichaltrigen bis geringfügig jüngeren Green-River-Formation. Während dieser Expeditionen kamen neben rund einem Dutzend Schädel, Schädelfragmenten und Unterkiefer zahlreiche Teile des Körperskelettes zu Tage. Herausragend sind weiterhin die Funde des United States Geologicals Survey von 1940, wobei unter anderem ein fast vollständiges Skelett nahe dem Henrys Fork und Sage Creek nordwestlich von Lonetree, Wyoming entdeckt wurde, dem lediglich ein Hinterbein und ein Teil des Vorderbeins sowie die Halswirbel fehlten.[7][3][1] Ab der zweiten Hälfte des 20. Jahrhunderts erfolgten zahlreiche Untersuchungen seitens des Field Museum of Natural History im Sweetwater County in Wyoming, wobei wenigstens elf Schädel und Schädelfragmente sowie einzelne Skelettpartien dokumentiert werden konnten. Hierbei ist auch der erste Fund eines relativ neugeborenen Tieres zu erwähnen. Zu den bedeutendsten jüngeren Funden gehört ein nahezu vollständiger, kaum zerdrückter und 75 cm langer Schädel, der in einem großen Sandsteinblock eingeschlossen war und 1984 am Salazar Butte Quadrangle im Sweetwater County in Wyoming aufgefunden wurde. Hier fanden sich später an der originalen Abbruchstelle des Blocks auch Reste des Körperskelettes, wie Unterkiefer, Wirbel, Rippen und Teile des Beckens, die vermutlich alle demselben Individuum angehören.[2]
Ursprünglich nur als nordamerikanisch verbreitete Gattung angesehen, wurden seit Beginn der 1980er Jahre häufiger Fossilien in Ostasien entdeckt. Hierzu gehört ein fast vollständiger Schädel aus der Lushi-Formation in der chinesischen Provinz Henan.[8] Ein Unterkiefer stammt aus der Nomogen-Formation im Erlian-Becken in Nei Mongol (Innere Mongolei), die bereits im Unteren Eozän entstand.[4][9] Dagegen kamen einzelne Zähne und postcraniale Skelettelemente aus der Üqbulak-Formation in Xinjiang zu Tage, welche wiederum in das Mittlere Eozän datiert.[10] Die ersten Funde von Uintatherium in Ostasien, darunter auch mehrere riesige Eckzähne, waren aber schon in den 1960er Jahren aus der Sinyu-Formation in Jiangxi publiziert worden.[11]
Paläobiologie


Uintatherium besaß einen Körperbau, der an heutige Nashörner erinnerte, jedoch weniger deutlich robust war, es besaß allerdings auch wesentlich längere Beine als diese, die sich auch in ihren Bau jenen von Rüsseltieren annäherten. Die Fortbewegung war mehr plantigrad (Sohlengänger) als bei den heutigen Elefanten, was sich aus der einfacher gebauten Vorder- und Hinterfüßen ergibt. Der langgestreckte Schädel lässt eine eher tiefe Kopfhaltung vermuten, so dass der hintere Teil des Hinterhauptsbeines eher vertikal verlief. Zu diesem Zweck setzten kräftige Nackenmuskeln am Hinterkopf an. Der typisch lophodonte Aufbau der Backenzähne gibt an, dass Uintatherium ein Pflanzenfresser war, der sich überwiegend von weicher Kost ernährte (browsing). Darauf weisen auch die typischen transversen Schleifmuster auf den Backenzähnen hin.[12] Zusätzlich besaßen die Zähne im Innern eine irreguläre Anordnung des Zahnschmelzes, um höheren Beanspruchungen zu widerstehen. Des Weiteren dienten die unteren Schneidezähne möglicherweise zum Rupfen der Nahrung, was aufgrund der häufig stark abgearbeiteten Zahnfunde vermutet wird. Das Fehlen der oberen Schneidezähne wiederum lässt eine kräftige und lange Zunge vermuten.[1][3] Das äußerst breite Becken, dass in seinen Proportionen ebenfalls an jenes der Rüsseltiere erinnert, lässt vermuten, dass bei Uintatherium ein sehr umfangreich entwickelter hinterer Darmtrakt entwickelt war. Dies führt zur Annahme, dass die Uintatherien möglicherweise ihre Nahrung im Enddarm verwerteten, ähnlich den heutigen Unpaarhufern. Über die Lebensweise ist ansonsten relativ wenig bekannt. Bemerkenswert sind die durch Pachyostose verdickten Langknochen, ob dies mit einer semi-aquatischen Lebensweise verbunden werden kann, wobei die verdickten Knochen den Auftrieb des Körpers im Wasser ausgleichen sollten, ist aber unklar.[2]
Die Fossilien von Uintatherium lassen einen deutlichen Geschlechtsdimorphismus erkennen. Die Eckzähne der männlichen Tiere waren wesentlich ausgeprägter und länger als bei weiblichen Tieren. Außerdem fehlten den Weibchen die markanten unteren Knochenauswüchse am Unterkiefer. Weiterhin waren bei ihnen die knöchernen Hörner weniger deutlich ausgebildet. Die Kombination von Hörnern und ausgeprägten Eckzähnen ist bei heutigen Huftieren unbekannt, auf diesen Umstand wurde erstmals Anfang der 1940er Jahre aufmerksam gemacht. Moschustiere und Wasserrehe besitzen lange obere, als Waffen genutzte Eckzähne, aber kein Geweih. Möglicherweise dienten auch bei Uintatherium die Eckzähne als Waffen im Paarungs- oder Verteidigungskampf. Einige Schädel zeigen zu Lebzeiten abgebrochene Eckzähne, die aber Schliffspuren tragen und eine weitere aktive Nutzung bezeugen. Auch die Funktion der sechs Hörner ist nicht eindeutig, auch hier ist eine Nutzung zur Verteidigung oder der Zurschaustellung sowie der Auseinandersetzung mit Artgenossen um das Paarungsvorrecht annehmbar, ähnlich heutigen geweihtragenden Paarhufern.[3][13]
Systematik
Uintatherium ist eine Gattung aus der Familie der Uintatheriidae. Innerhalb der Familie wurde Uintatherium in einer Revision der Uintatherien von Walter Wheeler aus dem Jahr 1961 zur Unterfamilie Uintatheriinae gestellt, die sich durch Vertreter mit großem Körperbau allgemein und drei Paaren knöcherner Hornbildungen auszeichnen und der weiterhin Eobasileus angehört. Ursprünglich wurde mit Tetheopsis eine dritte Gattung beschrieben, eine Teilrevision aus dem Jahr 2002 von William D. Turnbull, ließ jedoch die Eigenständigkeit dieser offen, da zumindest eine der ihr zugewiesenen zwei Arten identisch mit Eobasileus ist.[2] Den Uintatheriinae gegenüber stehen die Gobiatheriinae, die durch die Reduktion der oberen Eckzähne und einer im vorderen Teil verknöcherten Nasenscheidewand charakterisiert sind.[14][3] Während in der Revision 2002 der Status der Gobiatheriinae als Unterfamilie bestätigt wurde, sehen einige Forscher aufgrund im Detail abweichender Zahnmorphologien diese teilweise aber auch als eigenständige Familie innerhalb der Dinocerata an.[15]
Die Uintatheriidae werden zur Ordnung der Dinocerata gestellt, innerhalb dieser ist Uintatherium einer der größten und bekanntesten Vertreter. Diese Säugetiergruppe, die aus Nordamerika und Ostasien bekannt ist, zählte zu den ersten Riesenformen, die sich nach dem Aussterben der Dinosaurier entwickelten. Gemeinsames Merkmal aller Dinocerata stellt das Fehlen des ersten Prämolaren und der oberen Schneidezähne dar (mit Ausnahme der frühesten Vertreter).[16] Die verwandtschaftlichen Verhältnisse zu anderen Säugetieren sind nicht vollständig geklärt, eine in den frühen 1980er Jahren postulierte nähere Verwandtschaft über Pseudictops mit den Anagalidae, stammesgeschichtlich urtümlichen, den heutigen Nagetieren nahestehenden kleinen Säugetieren aus Ostasien, erwies sich als falsch. Diese mögliche Verwandtschaft wurde zumeist aufgrund gemeinsamer Zahnmerkmale angenommen, vor allem aber auch die im Verhältnis zur Körpergröße extrem kleinen Backenzähne bei den Uintatherien, jedoch widerlegten skelettanatomische Untersuchungen diese Vermutung. Heute werden die Dinocerata allgemein der umfangreichen, aber heterogenen Gruppe der Huftiere zugeordnet, wo sie einen sehr ursprünglichen Zweig bilden. Dabei sind die Dinocerata möglicherweise enger verwandt mit den „Südamerikanischen Huftieren“ (Meridiungulata), etwa den Pyrotheria[17] oder auch den Xenungulata. Die nächsten heute lebenden Verwandten stellen möglicherweise die Unpaarhufer dar. Mit diesen teilen sie die Ausbildung der Hufe, die mesaxonische Fußstellung (Schwerpunkt der Achse des Fußes läuft durch den mittleren (dritten) Strahl) und das Fehlen einer tiefen Furche am Sprungbein zur Artikulation mit dem Schienbeingelenk, die beispielsweise bei den Afrotheria häufig vorkommt.[3][18]
Ursprünglich umfasste Uintatherium in Nordamerika fast 30 Arten, die aber unter verschiedensten, heute synonymen Gattungsnamen geführt wurden, wie Dinoceras, Loxolophodon und Tinoceras. Nach einer Revision aus dem Jahr 1961 ist nur U. anceps gültig; diese Art erhielt bereits 1871 ihre Erstbeschreibung.[3] Aufgrund teils unterschiedlicher Merkmale im Zahnbau wird teilweise in Ostasien U. insperatus als eigenständige Gattung abgegrenzt.[4][8] Der Name Uintatherium setzt sich aus der Bezeichnung für die Uinta Mountains im Nordosten Utahs sowie Südwesten Wyomings und dem griechischen Wort θήριον (thêrion „Tier“) zusammen.[19]
Entdeckung und Benennung

Die Erforschung von Uintatherium hat eine lange und abwechslungsreiche Geschichte, in deren Verlauf zahlreiche unterschiedliche und teils unabhängig voneinander vorgenommene taxonomische Benennungen erfolgten. Die Entdeckungen begannen im Bridger- und Washakie-Becken im Süden des US-Bundesstaates Wyoming und fallen dabei in die Cope-Marsh-Fehde, welche die beiden Paläontologen Othniel Charles Marsh und Edward Drinker Cope in den 1870er und 1880er Jahren um prestigeträchtige Fossilien führten.[20] Die ersten Funde von Uintatherium wurden bereits 1870 von Marsh am Sage Creek im Bridger-Becken des südwestlichen Wyomings entdeckt, auf die Region war dieser durch Fossilfunde von Joseph Leidy aus dem Jahr 1868 aufmerksam geworden. Er verwies sie 1871 allerdings zu Titanotherium, einem Namen, der ursprünglich für die Gattung Megacerops aus der Gruppe der Brontotheriidae vergeben worden war.[21] Erst im darauf folgenden Jahr etablierte Marsh aufgrund neuer Funde die Namen Dinoceras (basierend auf einem vollständigen Schädel, Schulterblatt und Rippen) und Tinoceras (anhand eines vollständigen Schädels) für diese neue Säugetierform. Zu Tinoceras zählte er dabei auch jene ersten Funde von 1870, wobei er zwischenzeitlich auch eine Stellung dieser innerhalb der Mastodonten, urtümlichen Rüsseltieren, erwog. Seine Publikation erschien am 20. August des Jahres 1872. Nur drei Tage zuvor, am 17. August, hatte aber bereits Cope eine telegraphische Mitteilung an die Naturhistorische Akademie von Philadelphia verschickt, um gegenüber seinem Kontrahenten Marsh eine möglichst schnelle Publikation seiner Fossilien von Loxolophodon aus dem südlichen Wyoming zu erreichen – diese umfassten Reste von fünf verschiedenen Individuen, darunter auch Schädel und postcraniale Skelettteile –, so dass diese unmittelbar folgend im Palaeontological Bulletin veröffentlicht werden konnten. Jedoch verschrieb sich der Übermittler, wodurch der Name Lefalophodon bis heute überliefert ist. Zwischenzeitlich, am 20. August etablierte Cope anhand des gleichen Fundmaterials die Gattung Eobasileus, zu der ein Teil des Materials heute gestellt wird, erst am 22. August korrigierte er den Namen in seine ursprünglich vorgesehene Form Loxolophodon. Bereits am 1. August 1872 hatte allerdings Joseph Leidy Fossilien publiziert, die knapp zwei Wochen zuvor von ihm und einer Expedition am Dry Creek und Twin Buttes nahe Fort Bridger im Bridger-Becken gefunden worden waren, darunter einige Schädelknochen, Backenzähne und einen Oberarmknochen, die er der neuen Gattung Uintatherium zuwies. Einen ebenfalls entdeckten riesigen, gut 30 cm langen Eckzahn ordnete er damals Uintamastix zu, das seiner Meinung nach eher mit den Säbelzahnkatzen verwandt war.[19]

Erst nachdem die Funde veröffentlicht waren, erkannte zuerst Leidy 1873, dass sie jeweils die gleiche Säugetierform oder nahe verwandte Gattungen beschrieben hatten, auch musste er die Zugehörigkeit des riesigen Eckzahns, den er zuerst einer Raubkatze zuschrieb, als Teil dieses bizarren Säugetiers zugestehen. Leidys Entdeckung von Uintatherium ignorierten Marsh und Cope anfangs weitgehend, beide Forscher sahen dabei ihre jeweils vergebenen Bezeichnungen als die in ihren Augen gültige für diese neue Tierart und jene des Konkurrenten als Synonyme an. Allerdings erwies sich Leidys Beschreibung aufgrund der Regeln der zoologischen Nomenklatur als die wissenschaftlich gültige, da sie zuerst erfolgt war.[21] Cope stellte seine Entdeckungen in seiner heute als Cope-Bibel bezeichneten und mehr als 1000 Seiten umfassenden Publikation The Vertebrata of the Tertiary formations of the West im Jahr 1885 vor. Hier sah er die Funde als frühe Vertreter der Rüsseltiere an, was sich auch in einigen bekannten Lebendrekonstruktionen niederschlug, in denen die Tiere mit Rüssel dargestellt wurden. Marsh selbst veröffentlichte im gleichen Jahr seine Monographie Dinocerata, a monograph of an extinct Order of gigantic mammals über seine Funde,[13] in der er diese zur Gruppe der Dinocerata verwies, eine heute noch gültige Bezeichnung, auch wenn der eigentliche Gattungsname Dinoceras nur ein Synonym für Uintatherium ist; den Begriff Dinocerata kreierte er aber bereits 1873. Erst nach dem Ende der Cope-Marsh-Fehde und dem Tod der beiden Forscher Ende des 19. Jahrhunderts war es einer neuen Forschergeneration vorbehalten, die zahlreichen Fossilien genau zu klassifizieren. Die nachfolgende Tabelle stellt alle zwischen 1870 und 1873 in den ersten Publikationen beschriebenen Uintatherium-Formen vor, etwa ein weiteres Dutzend neu beschriebener Arten folgten bis 1885, die hauptsächlich von Marsh etabliert wurden.[3][22]
| Taxon | Autor | Jahr der Beschreibung | Typusexemplar | Funde | Fundort |
|---|---|---|---|---|---|
| Titanotherium? anceps | Marsh | 1871 | YPM 11030 | Schädel, Wirbel, Langknochen | Sage Creek, Bridger-Becken (Wyoming) |
| Uintatherium robustum | Leidy | 1872 | ANSP 12607, 12609–126013, 12619, 12622 | Schädel- und Unterkieferfragmente | Twin Buttes, Bridger-Becken (Wyoming) |
| Uintatherium (Uintamastix) atrox | Leidy | 1872 | ANSP 12606, 12608 | Oberkiefereckzahn, Prämolaren | Dry Creek, Bridger-Becken (Wyoming) |
| Loxolophodon furcatus ("Lefalophodon bifurcatus") | Cope | 1872 | AMNH 5045 | Schädel | Haystack Mountain, (Wyoming) |
| Loxolophodon pressicornis | Cope | 1872 | AMNH 5042 (Nr. 3), 5043 | Schädel, Langknochen | Haystack Mountain, (Wyoming) |
| Tinoceras grande | Marsh | 1872 | YPM 11040 | Schädel, Hals- und Brustwirbel | Barrel Springs, Washakie-Becken (Wyoming) |
| Dinoceras mirabile | Marsh | 1872 | YPM 11036 | Schädel, Hals-, Brust- und Lendenwirbel | Big Bone Buttes, Bridger-Becken (Wyoming) |
| Dinoceras (Tinoceras) lacustre | Marsh | 1872 | YPM 11037 | Schädel | Bitter Creek, Washakie-Becken (Wyoming) |
| Dinoceras lucare | Marsh | 1873 | YPM 11038 | Schädel | Big Bone Buttes, Bridger-Becken (Wyoming) |
| Dinoceras laticeps | Marsh | 1873 | YPM 11039 | Schädel | Spanish John’s Meadow, Bridger-Becken (Wyoming) |
Literatur
- T. S. Kemp: The Origin & Evolution of Mammals. Oxford University Press, Oxford 2005. ISBN 0198507615
- William D. Turnbull: The Mammalian Faunas of the Washakie Formation, Eocene Age, of Southern Wyoming. Part IV. The Uintatheres. Fieldiana 47, 2002, S. 1–189
- Walter H. Wheeler: Revision of the Uintatheres. Bulletin of the Peabody Museum of Natural History Yale University 14, 1961, S. 1–93
Einzelnachweise
- Henry Fairfield Osborn: A memoir upon Loxolophodon and Uintatherium, two genera of the sub-order Dinocerata. Contributions from the E. M. Museum of Geology and Archeology of the College of new Jersey 1 (1), Princeton, 1881
- William D. Turnbull: The Mammalian Faunas of the Washakie Formation, Eocene Age, of Southern Wyoming. Part IV. The Uintatheres. Fieldiana 47, 2002, S. 1–189
- Walter H. Wheeler: Revision of the Uintatheres. Bulletin of the Peabody Museum of Natural History Yale University 14, 1961, S. 1–93
- Bai Bin: New Materials of Eocene Dinocerata (Mammalia) from the Erlian basin, Nei Mongol (Inner Mongolia). Vertebrata Palasiatica 44 (3), 2006, S. 250–261
- Horace Elmer Wood: The problem of the Uintatherium molars- Bulletin American Museum of Natural History 48, 1923, S. 599–604
- Gregg F. Gunnell, Paul C. Murphey, Richard K. Stucky, K. E. Beth Townsend, Peter Robinson, John-Paul Zonneveld und William S. Bartels: Biostratigraphy and biochronology of the Latest Wasatchian, Bridgerian, and Uintan North American mammal "ages". In: L. B. Albright III (Hrsg.): Papers on Geology, Vertebrate Paleontology, and Biostratigraphy in Honor of Michael O. Woodburne. Museum of Northern Arizona Bulletin 65, 2009, S. 279–330
- Nature: Fossil skeleton of Uintatherium. Nature 147, 1941, S. 114
- Tong Yongsheng und Wang Jinweng: A skull of Uintatherium from Henan. Vertebrata Palasiatica 19 (3), 1981, S. 208–213
- Jin Meng, Yuanqing Wang, K. Christopher Beard, Chengkai Sun, Qian Li, Xun Jin und Bin Bai: New Stratigraphic Data from the Erlian Basin: Implications for the Division, Correlation, and Definition of Paleogene Lithological Units in Nei Mongol (Inner Mongolia). American Museum Novitates 3570, 2007, S. 1–31
- Tong Yongsheng: Some Eocene Mammals from the Üqbulak area of the Junggar basin, Xinjiang. Vertebrata Palasiatica 27 (3), 1989, S. 182–196
- Chow Minchen und Tong Yongsheng: Notes on some new Uintathere materials of China. Vertebrata Palasiatica 6 (4), 1962, S. 368–374
- Wighart V. Koenigswald und Kenneth D. Rose: The Enamel Microstructure of the Early Eocene Pantodont Coryphodon and the Nature of the Zigzag Enamel. Journal of Mammalian Evolution 12 (3/4), 2005, S. S. 419–432
- L. P.: Professor Marsh's Monograph of Dinocerata. American Journal of Sciences 3, 24, 1885, S. 173–204
- Spencer George Lucas: Gobiatherium (Mammalia: Dinocerata) from the Middle Eocene of Asia: Taxonomy and biochronological Significance. Paläontologische Zeitschrift 74 (4), 2001, S. 591–600
- J. G. M. Thewissen und P. D. Gingerich: Systematics and evolution of Probathyopsis (Mammalia, Dinocerata) from the Late Paleocene and Early Eocene of Western North America. Contributions from the Museum of Paleontology The University of Michigan 8, 1987, S. 195–219
- A. H. Garrod: On the Order Dinocerata. Journal of Anatomy and Physiology 7 (2), 1873, S. 267–270
- Donald R. Prothero, Earl M. Manning und Martin Fischer: The phylogeny of the ungulates. In: M. J. Benton (Hrsg.): The phylogeny and classification of Tetrapods, Volume 2: Mammals. Systematics Association, Special Volume 35B, Oxford, 1988, S. 201–234
- Benjamin J. Burger: The systematic position of the sabre-toothed and horned giants of the Eocene: The uintatheres (Order Dinocerata). Journal of Vertebrate Paleontology 35 (suppl.), 2015, S. 99
- Joseh Leidy: On some new species of fossil mammalia from Wyoming. Proceedings of the Academy of Natural Sciences Philadelphia 24, 1872, S. 167–169 ()
- Walter H. Wheeler: The Uintatheres and the Cope-Marsh War. Sciences 131, 1960, S. 1171–1176
- W. H. Flower: The Uintatherium. Nature 13, 1876, S. 404–405
- Donald R. Prothero und Robert M. Schoch: Horns, tusks, and flippers. The evolution of hoofed mammals. Johns Hopkins University Press, Baltimore, 2003, ISBN 0-8018-7135-2 (S. 9–13)
Weblinks
- The Paleobiology Database Uintatherium