T-Helferzelle
T-Helferzellen (abgekürzt TH-Zellen) sind eine Gruppe der T-Lymphozyten, die eine Helferfunktion haben. Sie werden anhand der von ihnen ausgeschütteten Zytokine in zwei wichtige Untergruppen (Subpopulationen) eingeteilt, die verschiedene Funktionen haben. Eine Untergruppe (genannt Typ1-T-Helferzellen) ist an der zellulären Immunantwort beteiligt, während die andere Untergruppe (genannt Typ2-T-Helferzellen) an der humoralen Immunantwort beteiligt ist. Ein weiterer T-Helferzelltyp wurde im Jahr 2005 entdeckt. Dieser Typ bildet u. a. IL-17 und spielt daher eine Rolle bei der Regulierung von Entzündungsvorgängen.[1]
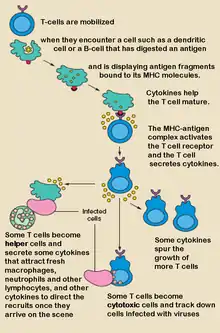
Allgemeine Charakterisierung der T-Helferzellen
Die funktionell gegensätzliche Aufteilung (Dichotomie) in Typ1- und Typ2-T-Zellen ist beispielhaft für CD4-positive Lymphozyten beschrieben worden, gilt aber auch für CD8-positive T-Zellen und für Zellen mit einem γδ-T-Zell-Antigenrezeptor. Als Typ1-T-Zellen werden CD4-positive und CD8-positive Lymphozyten bestimmt, die typischerweise Interferon-γ (IFN-γ), IL-2 und TNF-α ausschütten. Entsprechend werden CD4-positive oder CD8-positive Lymphozyten, die als typische Zelleigenschaft die Ausschüttung der Zytokine IL-4, IL-5, IL-6, IL-10 und IL-13 haben, als Typ2-T-Zellen bezeichnet.
Diese Einteilung ist eine vereinfachte Betrachtung aufgrund extremer Reaktionsmöglichkeiten der Zytokin-sezernierenden T-Zellen. Sowohl im Gewebe als auch im peripheren Blut findet man T-Zellen mit einem Zytokinmuster, das typisch sowohl für Typ1- als auch für Typ2-T-Zellen ist. Diese Zellen werden gelegentlich als Typ0-T-Zellen bezeichnet.
Die Unterschiede zwischen Typ1-T-Zellen und Typ2-T-Zellen wurden erstmals 1986 von Tim Mosmann und Robert L. Coffman beschrieben.[2]
Typ1-T-Helferzellen (TH1-Lymphozyten)
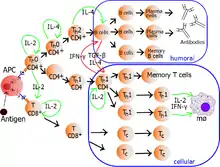
Die zellvermittelte Abwehr des erworbenen Immunsystems erfolgt überwiegend mittels aktivierter T-Lymphozyten und Makrophagen. Bestimmte Antigene bewirken die Aktivierung, beschleunigte Proliferation und Differenzierung von T-Zellen mit Spezifität für diese Antigene. Für aktivierte Typ1-T-Zellen ist die Ausschüttung von Zytokinen wie IFN-γ, IL-2 und TNF-α charakteristisch, durch die die Aktivierung und Differenzierung von Makrophagen erfolgt.
Die Makrophagen wiederum fördern durch ihre Zytokinausschüttung (IL-12) die Typ1-Polarisierung reaktiver T-Zellen. Gleichzeitig intensivieren die Makrophagen ihre Wirkung als antigenpräsentierende Zellen (APC) und verbessern ihre antimikrobielle Fähigkeit, gegen extra- und intrazelluläre Antigene vorzugehen. Es erfolgt die Hochregulation der ko-stimulierenden Oberflächenmoleküle CD40, CD80 und CD86 und der MHCII-Moleküle. In Folge nimmt die MHC-Produktion und Peptidprozessierung (also insgesamt die Peptidpräsentation) zu.
Die T-Zell-vermittelte Entzündungsreaktion steigert den Blutfluss über die Erweiterung der lokalen Gefäße (Vasodilatation). Die Hochregulation von Adhäsionsmolekülen auf Endothelzellen führt zur gesteigerten Einwanderung weiterer Zellen aus dem peripheren Blut ins Gewebe. Als lokal entzündungshemmender Faktor wird schließlich Fibrin gebildet. An der Hemmung der Ausbreitung von Krankheitsauslösern (Pathogenen) sind auch CD8-positive T-Zellen beteiligt. Über ihre Ausschüttung von IFN-γ und direkte Zytotoxizität tragen sie zum Gewebeschaden bei.
B-Lymphozyten werden auch (aber nicht so stark wie durch Typ2) angeregt, und zwar durch Klassenwechsel zur Produktion von opsonisierenden Antikörpern (vor allem IgG), das heißt solchen Antikörpern, die Objekte durch Makrophagen fressen lassen können. Gleichzeitig wird eine Typ2-Antwort durch die ausgeschütteten Zytokine gehemmt. Ausgelöst wird eine solche Antwort durch IFN-γ und IL-12, welche von Natürlichen Killerzellen (NK-Zellen) und Dendritischen Zellen gebildet werden. Dieser Weg dient insbesondere der Bekämpfung einiger intrazellulärer Bakterien sowie von Virusinfektionen. Die Produktion von IFN-γ blockiert gleichzeitig die Differenzierung zu Typ2-Zellen.
Darüber hinaus sind Typ1-Zellen in der Lage, Infektionen zu bekämpfen, die durch Bakterien hervorgerufen werden, welche sich in den Vesikeln von Makrophagen vermehren. Beispiele hierfür sind der Erreger der Tuberkulose (Mycobacterium tuberculosis) und der Lepra-Erreger (Mycobacterium leprae). Die Typ1-Zellen aktivieren die befallenen Makrophagen und lösen dadurch die Fusion der befallenen Vesikel mit Lysosomen aus, wodurch die Erreger verdaut werden.
Typ2-T-Helferzellen (TH2-Lymphozyten)
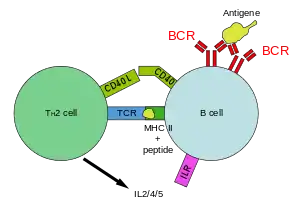
Die wichtigste Funktion der Typ2-polarisierten CD4-positiven T-Zellen ist die Interaktion mit B-Lymphozyten, die über Zytokine und zellständige Moleküle stattfindet und bei diesen zur Produktion und Ausschüttung von Antikörpern führt. Dies ist für die humorale Immunantwort wesentlich und erfolgt durch das typische Zytokinmuster von Typ2-CD4-positiven-T-Zellen (IL-4, IL-5, IL-6, IL-10, IL-13 und Lymphotoxin-α).
Die von Typ2-T-Zellen ausgeschütteten Zytokine hemmen die Mechanismen, die zu einer Typ1-Antwort führen. Durch diese Kreuzhemmung wird eine einmal eingeschlagene Richtung für die Immunantwort beibehalten. Besonders IL-10 hemmt die Makrophagenaktivierung. Die bei der Immunantwort von Typ2-T-Zellen gewährte Antigen-spezifische Hilfe führt bei B-Zellen nicht nur zu ihrer Aktivierung, Proliferation und Differenzierung, sondern stimuliert zusätzlich Mastzellen sowie basophile und eosinophile Granulozyten. Innerhalb der Reihe der Zytokine, die die Sekretion von Immunglobulinen stimulieren, hat IL-4 eine besondere Funktion. Es ist im Rahmen der Aktivierung und Differenzierung von B-Zellen vor allem für die Bildung von neutralisierenden Antikörperklassen, das heißt für einen Klassenwechsel (englisch isotype switch) zu IgA, IgG, IgE verantwortlich. Typ2-T-Zellen können B-Zellen dabei sehr stark anregen. So wird die humorale Immunantwort vor allem durch das Zytokin IL-4 ausgelöst, IL-6 verstärkt diese Tendenz. IL-4 hemmt die Aktivierung von Makrophagen. IL-10 reduziert die Produktion von IFN-γ in T-Zellen und Makrophagen.
Typ17-T-Helferzellen (TH17-Lymphozyten)
TH17-Lymphozyten scheinen eine wichtige Rolle bei der Regulierung von Entzündungsvorgängen zu spielen. Dabei ist Interleukin-6 ein notwendiges Signal, das die Umwandlung einer naiven CD4-positiven T-Zelle in eine TH17-Zelle koordiniert. Die Rolle der TH17-Lymphozyten ist noch nicht abschließend geklärt und Gegenstand aktueller Forschung.[1]
Antigenerkennung der CD4-positiven T-Zellen
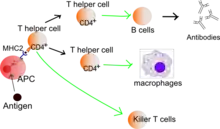
In ihrer Funktion als antigenpräsentierende Zellen können B-Zellen, dendritische Zellen oder Makrophagen Antigene anhand bestimmter Epitope erkennen und aufnehmen. Im Zellinneren wird das Antigen prozessiert und in Form von kleineren Peptiden gemeinsam mit MHC-Klasse-II Molekülen den CD4-positiven T-Zellen präsentiert.
Aktivierte CD4+-T-Helferzellen spielen eine entscheidende Rolle für die Aktivierung (Lizenzierung) reifer, antigenpräsentierender dendritischer Zellen.[3] Die Hilfe der CD4+-T-Zellen ist zwingend erforderlich, um eine langanhaltende, effektive Immunantwort zytotoxischer CD8+-T-Zellen zu erzeugen. Nur wenn die dendritische Zelle gleichzeitig mit der T-Helferzelle und der CD8+-T-Zelle interagiert, ist eine effektive, sekundäre Expansion der zytotoxischen T-Zellen möglich.[4]
Literatur
- Georg Andreas Peter Holländer: Immunologie, Grundlagen für Klinik und Praxis. 1. Auflage. Elsevier, München 2006, ISBN 3-437-21301-6.
- Michael John Owen, Jonathan R. Lamb: Immunerkennung. Thieme, Stuttgart 1991, ISBN 3-13-754101-8.
Einzelnachweise
- Harrington, Laurie E, Robin D Hatton, Paul R Mangan, Henrietta Turner, Theresa L Murphy, Kenneth M Murphy, und Casey T Weaver.: Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. In: Nature Immunology. Band 6, Nr. 11, 2005, S. 1123–1132, PMID 16200070.
- Tim Mosmann, H Cherwinski, M. W. Bond, M. A. Giedlin, R. L. Coffman: Two types of murine helper T cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins. In: The Journal of Immunology. Jg. 136, Nr. 7, 1986, S. 2348–2357 (Abstract). ISSN 0022-1767
- Immunol Res. 2009 Dec;45(2-3):209-17. doi:10.1007/s12026-009-8102-0. Epub 2009 Feb 18. Unravelling the mechanisms of help for CD8+ T cell responses. Livingstone AM., et al.
- Eur J Immunol. 2014 Dec;44(12):3543-59. doi:10.1002/eji.201444477. Epub 2014 Oct 27. Concurrent interaction of DCs with CD4(+) and CD8(+) T cells improves secondary CTL expansion: It takes three to tango. Hoyer S., et al.