Goldspecht
Der Goldspecht (Colaptes auratus), manchmal auch als Kupferspecht bezeichnet, ist eine Art aus der Gattung der Goldspechte (Colaptes) innerhalb der Unterfamilie der Echten Spechte. Die gut grünspechtgroße Spechtart kommt in vier deutlich differenzierten Unterartengruppen in weiten Bereichen des nördlichen Amerikas und Teilen Mittelamerikas sowie auf Kuba vor. Wie die meisten Arten dieser Gattung lebt auch der Goldspecht vor allem am Boden, wo er sich, häufig in kleineren Gruppen versammelt, von Insekten, vornehmlich Ameisen ernährt. Er ist für viele andere höhlenbewohnende Vögel, Säugetiere und Insekten ein wichtiger Höhlenlieferant. Der lange als Unterart des Goldspechts betrachtete Wüstengoldspecht wurde 1995 als eigenständige Art abgetrennt.[1] Obwohl regional der Bestand der Art abnimmt, ist der Goldspecht in weiten Teilen seines Verbreitungsgebietes eine häufige und gut bekannte Spechtart, die von der IUCN als ungefährdet eingestuft wird.
| Goldspecht | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
_B._Walker.jpg.webp)
Goldspecht (Colaptes a. auratus) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Colaptes auratus | ||||||||||||
| (Linnaeus, 1758) |
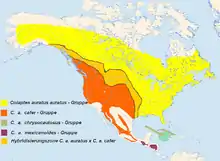
Die taxonomische Situation ist nicht restlos geklärt. Meist wurden 11 Unterarten beschrieben, die sich auf vier Unterartengruppen verteilten. 2014 wurde vom HBW eine Änderung vorgeschlagen, die die Vögel der Colaptes cafer – Gruppe, sowie Colaptes mexicanoides in Artrang stellt.
Aussehen
Mit einer Größe von bis zu 35 Zentimetern und einem maximalen Gewicht von etwa 180 Gramm ist der Goldspecht gut grünspechtgroß, aber im Durchschnitt etwas leichter als dieser. Die Spannweite beträgt bis zu 54 Zentimeter. Weibchen sind im Jahresdurchschnitt mit bis zu 20 Prozent signifikant leichter als Männchen. Die größte und schwerste Unterart ist C. a. cafer aus dem nordwestlichen Kanada, die kleinste C. a. gundlachi von Grand Cayman.[2]
Colaptes auratus auratus

Die Oberseite ist auf bräunlichem Grund unregelmäßig dunkel gebändert. Die hinteren Rumpfseiten sind ungebändert weiß, gelegentlich sind unregelmäßige schwarze Tupfer und Flecken eingestreut. Die Oberflügeldecken und die Oberschwanzdecken sind dunkel gebändert bräunlich, letztere weisen ein schwarzes Subterminalband auf. Die Schwingen sind dunkelbraun, eher undeutlich hell bräunlich gebändert, die Federschäfte sind gelb. Die Schwanzoberseite ist schwarzbräunlich, die Außenfahnen der äußeren Steuerfedern andeutungsweise weiß gefleckt. Die Zentralfedern sind gelb geschaftet, die Schwanzspitze ist undeutlich ledergelb gefärbt. Die Unterseite des Rumpfes ist hinter dem markanten halbmondförmigen schwarzen Brustabzeichen weiß, besonders an den Flanken leicht rahmfarben. Der Bauch ist unregelmäßig schwarz gefleckt, an den Flanken eher pfeilspitzenartig schwarz gezeichnet und zu den Unterschwanzdecken hin schwärzlich gebändert. Die Flügelunterseite ist gelblich, an den Spitzen und Rändern eher gelbbraun. Die Unterflügeldecken weisen schwarze Tupfer auf. Die Schwanzunterseite ist ebenfalls gelblich, die Schwanzspitze ist schwarz.

Der Kopf ist von der Stirn bis in den unteren Nackenbereich und an den Schläfenseiten bläulichgrau. Im unteren Nackenbereich befindet sich beim Männchen ein markanter, V-förmiger roter Nackenfleck. Wangen und Ohrdecken, Kehle und obere Brustseite sind zimtfarben. Der Bartstreif ist schwarz.
Der lange, spitze, abwärts gebogene Schnabel ist schieferfarben oder schwarz, die Beine und die vier zygodactyl angeordneten Zehen sind grau bis grünlichgrau. Die Iris der Augen ist satt braun.
Weibchen sind geringfügig kleiner und nicht unerheblich leichter, jedoch reichen diese Unterschiede nicht zu einer sicheren Geschlechtsbestimmung im Feld aus. Sicherster Färbungsunterschied ist das Fehlen des schwarzen Bartstreifs; diese Region ist beim Weibchen zimtfarben wie die übrige Kopfseite. Der halbmondförmige Brustlatz ist etwas weniger intensiv schwarz als beim Männchen und etwas kleiner.
Jungvögel ähneln bereits stark adulten; die Kopfseiten sind bei ihnen eher grau, der Scheitel häufig etwas schwarz gefleckt. Oft ist ein schwarzer Zügelfleck zu sehen. Die oberen Flügeldecken weisen stärkere Schwarzmarkierungen auf, die dunkle Bänderung der Oberseite ist breiter. Immature Männchen zeigen am Scheitel oft einen rötlichen Farbeinschlag, ihr Bartstreif ist noch nicht deutlich ausgebildet. Auch bei immaturen Weibchen kann ein angedeuteter Bartstreif vorhanden sein.
Der Flug des Goldspechtes ist rasch und typisch-spechtartig bogenförmig. Der Aufschwung erfolgt durch eine Folge schneller Flügelschläge, beim Abschwung sind die Flügel angelegt.
Färbung der Federschäfte
Die Färbung der Schäfte der Schwingen und der zentralen Steuerfedern, sowie von großen Teilen der Flügel- und Schwanzunterseite sind neben den unterschiedlichen Merkmalen der Kopffärbung die wesentlichsten Unterscheidungsmerkmale zwischen den verschiedenen Unterarten. Der Hauptgürtel der Rocky Mountains bildet die Trennlinie zwischen den goldgelb gefärbten Spechten der C. a. auratus – Gruppe im Osten und den Spechten mit rötlichen oder lachsfarbenen Federschäften der C. a. cafer (Syn. Colaptes cafer) – Gruppe im Westen. C. a. mexicanoides (Syn. Colaptes mexicanoides) steht C. cafer nahe und ist ähnlich gefärbt.
Die Färbungsunterschiede werden mit einer unterschiedlichen Metabolisierung von β-Carotiniden erklärt; früher angenommene unterschiedliche Nahrungsbestandteile spielen keine Rolle.[3]
Etwa ein Drittel der Individuen der Colaptes auratus – Gruppe weist einzelne rötliche oder lachsfarbene Federschäfte auf. Der Anteil der unterschiedlich gefärbten Federn kann beim gleichen Individuum von Jahr zu Jahr variieren. Rückwirkungen von zurückliegenden Hybridisierungen wurden als Erklärung für diese Erscheinung herangezogen. In einer 2017 publizierten Untersuchung weisen Jocelyn Hudon et al. nach, dass die Rhodoxanthin enthaltenden Beeren von schon im frühen 18. Jh. im östlichen Nordamerika eingeführten, ursprünglich aus Ostasien stammenden Heckenkirschen, für die abweichende Färbung einzelner Federschäfte verantwortlich sind, und Auswirkungen von Mischbruten keine Rolle spielen. Ähnliche Farbabweichungen wurden auch beim Zedernseidenschwanz und beim Baltimoretrupial beobachtet, die ebenfalls Beeren dieser Heckenkirschen konsumieren.[4]
Stimme
Der Goldspecht ist eine sehr ruffreudige Art. Der häufigste Ruf, der von der Revieretablierung bis zur Paarbildung zu hören ist, ist eine laute Serie unterschiedlich transkribierter Laute. Meist werden sie mit wick…wick…wick oder kick…kick…kick wiedergegeben. Die durchschnittliche Dauer dieses Rufes liegt bei über 5 Sekunden; er kann aber auch fast 20 Sekunden andauern. Daneben spielen die Wicka – Rufe die wesentlichste akustische Rolle. Sie sind relativ leise und werden unterschiedlich gereiht, betont und moduliert. Die Rufreihen, die vor allem bei sexuell gestimmten Interaktionen zu hören sind, dauern bis zu 4 Sekunden. Das Trommeln der Art wird mit dem Geräusch, das ein Miniatur-Luftdruckbohrer erzeugt, beschrieben. Es ist eher leise, die Einzelschläge verbleiben auf einer Tönhöhe und folgen sehr rasch aufeinander.[5]
Mauser
Wie alle Echten Spechte schlüpfen Goldspechte unbefiedert. Das erste Nestlingsgefieder beginnt ab dem sechsten Tag zu wachsen. Die Mauser dieses Gefieders beginnt mit dem Ausfliegen und ist mit dem Wechsel der zentralen Steuerfedern abgeschlossen. Vom Großgefieder werden die Armschwingen nicht oder nur zum Teil vermausert. Ins Adultgefieder mausern Goldspechte im darauf folgenden Sommer. Die Großfedern fallen absteigend, beginnend mit den proximalen Handschwingen, die Steuerfedern werden paarweise gewechselt, wobei die kürzeren äußeren zuerst ausfallen.[6]
Systematik
.jpg.webp)
_female.JPG.webp)
Der Goldspecht ist eine von 9 Arten der Gattung Colaptes innerhalb der Tribus Malarpicini, die mittelgroße bis annähernd große, ihre Nahrung meist auf dem Boden suchende Arten umfasst. Bis auf Colaptes auratus und den erst vor einigen Jahren von diesem abgetrennten Colaptes chrysoides sind alle anderen Mitglieder der Gattung in Südamerika beheimatet.[7] Colaptes ist am nächsten mit der Gattung Piculus verwandt, deren kleine bis knapp mittelgroße Arten in Mittel- und Südamerika vorkommen. Neuere phylogenetische Arbeiten zeigen, dass einige Arten der Gattung Piculus eher zur Gattung Colaptes gehören.[8]
10–11 Unterarten werden zurzeit anerkannt, die in vier Unterartengruppen eingeteilt werden können. Am weitesten verbreitet ist C. a. auratus. Diese Unterart ist oben beschrieben.
- Colaptes auratus auratus (Linnaeus, 1758) - Gruppe: Sie umfasst neben der Nominatform
- die in den nördlichen USA, Kanada und in Alaska brütende Unterart C. a. luteus Bangs, 1898. Sie ist durchschnittlich etwas größer als C. a. auratus, gleicht ihr ansonsten in den meisten Färbungsdetails. Die meisten Populationen dieser Unterart sind Mittelstrecken-, einige Langstreckenzieher.
- C. a. chrysocaulosus Gundlach, 1848: Diese Unterart kommt auf Kuba vor ist dort stellenweise häufig. Oberseite etwas heller als die Nominatform.
- C. a. gundlachi Cory, 1886: Diese kleinste Unterart ist nur auf Grand Cayman heimisch. Oberseite noch eine Spur heller, Grundfarbe der Unterseite fast rein weiß.

- Colaptes auratus cafer – Gruppe
- Sie umfasst 5 Unterarten. Gemeinsam ist ihnen das Fehlen eines roten Nackenabzeichens; die Scheitel- und Nackenregion ist bräunlich. Die Malarregion ist beim Männchen rot, bei einigen Weibchen intensiv zimtfarben. Die Federschäfte der Schwingen und der mittleren Steuerfedern sind wildlachsfarben. Wangen, Ohrdecken, Kinn und Kehle sind grau. Unterschieden werden die Unterarten
- C. c. cafer (Gmelin, 1788), die im Pazifikgürtel vom südlichsten Alaska bis ins nördlichste Kalifornien vorkommt,
- C. a. collaris Vigors, 1829, eine etwas hellere und kleinere Unterart lebt im kalifornischen Küstenstreifen südwärts bis ins nördliche Niederkalifornien. Zusätzlich besiedelt diese Unterart einige, der der kalifornischen Küste vorgelagerten Inseln, wie beispielsweise die Santa Cruz Inseln. Die nördlichsten Populationen unternehmen außerhalb der Brutzeit kleinräumige Wanderungen nach Süden.
- C. a. mexicanus Swainson, 1827: kleine und bräunliche Unterart aus Zentralmexiko.
- C. a. nanus Griscom, 1934: die kleinste Unterart der cafer – Gruppe entspricht in ihrer Größe etwa der Nominatform. Sie brütet im südöstlichen Texas und im nordöstlichen Mexiko.
- C. a. rufipileus Ridgway, 1876 †: Diese Unterart war auf der Insel Guadalupe vor der niederkalifornischen Küste heimisch.
- C. a. mexicanoides Lafresnaye, 1844: Sie kommt von Südmexiko bis Nicaragua vor. Sie Spechte sind groß, ähneln jenen der cafer-Gruppe, weisen aber eine kastanienbraune Stirn-, Scheitel und Nackenfärbung auf. Der rote Bartstreif des Männchens ist oft schwarz gerahmt. Die Federschäfte sind orange, der schwarze Brustlatz ist weniger deutlich halbmondförmig.
- C. a. chrysocaulosus: Die Vögel dieser Gruppe leben bedeutend stärker baumgebunden als die übrigen Unterarten. Sie zählen zu den kleinsten Unterarten. In der Kopf und Schaftfärbung entsprechen sie den Spechten der auratus – Gruppe. Die Oberseitenfärbung hat einen deutlichen grünlichen Anflug, die Unterseite ist sehr stark schwarz gefleckt.
In einer Revision der Taxonomie schlägt das HBW eine Abtrennung der Colaptes cafer – Gruppe als eigenständige Art mit 5 rezenten Unterarten, von denen eine, der Guadalupe-Goldspecht (C. cafer rufipileus) kurz nach 1906 ausgestorben ist.[9] Ebenso wird Colaptes mexicanoides in Artrang gestellt.[10] Nach dieser Einschätzung würden neben der Nominatform noch C. a. luteus, C. a. chrysocaulosus und C. a. gundlachi als Unterarten des Goldspechtes zu betrachten sein.[11]
Hybridisierungen
Alle Unterarten hybridisieren in ihren Kontaktzonen. Die Nachkommen zeigen Merkmale beider Elternteile in stufenloser Variation. Die längste Hybridisierungszone, die in ihrer Länge und Breite in den letzten Jahrzehnten im Wesentlichen stabil blieb, erstreckt sich von Südalaska bis Texas. Ob Partner aus der eigenen Unterartengruppe bevorzugt werden, ist unklar. Nicht bestätigt wurden bisher auch Mischbruten mit dem Wüstengoldspecht. Eine sehr umfangreiche Untersuchung von Colaptes auratus x Colaptes cafer ssp. Mischbruten konnte keinerlei genetische Handicaps der Nachkommen feststellen.[12]
Verbreitung und Lebensraum
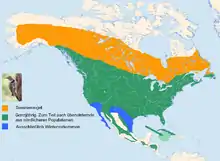
Der Goldspecht ist vom nördlichen Übergang der Baumtundra in die Taiga fast flächendeckend über ganz Nordamerika südwärts bis Nordmexiko verbreitet. Weiter nach Süden hin in Zentral- und Südmexiko beziehungsweise in Mittelamerika wird das Verbreitungsgebiet fragmentierter und zerfällt in einzelne Verbreitungsinseln. Zusätzlich bewohnt die Art Kuba und Grand Cayman. Nicht besiedelt sind die Wüstengebiete (Mojave und Sonora) im Südwesten der USA und Nordwesten Mexikos und die Halbinsel Niederkalifornien. Diese Gebiete sind der Verbreitungsraum des Wüstengoldspechts. Auch völlig baumlose Gebiete wie etwa die Great Plains kann die Art nur entlang baumbestandener Flüsse besiedeln.
Die Art ist einige Male in Europa, vor allem in Irland und in Schottland, aufgetreten. Wahrscheinlich überquerten die Vögel den Atlantik zumindest zum Teil auf Schiffen. Ein Nachweis stammt aus Dänemark.[13]
Entsprechend seinem sehr großen Verbreitungsgebiet bewohnt der Goldspecht eine Vielzahl unterschiedlicher Landschaftsformen. Wesentlich ist neben dem ausreichenden Nahrungsangebot auf offenen, nur mit niedriger Vegetation bewachsenen Flächen ein ausreichendes Angebot geeigneter Höhlenbäume. Solche Voraussetzungen findet die Art in lichten Wäldern, insbesondere am Rande großflächiger Kahlschläge, in jungen Aufforstungen oder in von Brandereignissen, Windwurf oder Schädlingsbefall heimgesuchten Waldgebieten. Er besiedelt baumbestandenes Farmland, vor allem Weideflächen, und dringt in dicht besiedelten Regionen in größere Gärten, Parks und in ähnliche Grüninseln vor. Was die Baumzusammensetzung seines Lebensraumes anbelangt, ist die Art flexibel. Im Norden spielt die Amerikanische Zitterpappel als Brutbaum eine herausragende Rolle. In Mexiko bewohnt die Art lichte Eichen–Kiefern Mischgehölze, in Mittelamerika vor allem Sekundärwälder im Regenwaldgürtel.
Soweit die Tiere Zugvögel sind, entsprechen die Winterhabitate weitgehend jenen der Brutsaison. Gelegentlich, vor allem auf Grand Cayman und auf Kuba, wird beobachtet, dass die Art winters dichter bewachsene Gebiete aufsucht.
Die vertikale Verbreitung der Art reicht vom Meeresniveau bis in Höhen von an die 4000 Meter in Mexiko.
Da die Art nur geringe interspezifische Aggression zeigt und auch in Gruppen nach Nahrung sucht, wird außer der Brut-, eventuell auch einiger Schlafhöhlen kein Territorium gegen Artgenossen verteidigt. Der Brutplatz selbst wird jedoch energisch sowohl gegen Artgenossen als auch gegen artfremde Höhlenkonkurrenten behauptet. Die Abstände aktiver Nisthöhlen können im Extremfall unter 10 Meter betragen, liegen aber meistens zwischen 100 und 200 Metern.[14]
Wanderungen
Je nach Brutgebiet sind Goldspechte Fernzieher, Kurz- oder Mittelstreckenzieher oder Standvögel. Die Zugdistanzen nehmen von Norden nach Süden hin ab. Vor allem die Mehrheit der nördlichen Populationen der auratus−Gruppe verlassen ab Mitte September ihre Brutgebiete und ziehen meist in südöstliche Richtungen. Die Rückkehr in die Brutgebiete erfolgt ab Mitte März. Als Grenze zwischen vornehmlich Zugvögel und vornehmlich Standvögel wird in etwa der 38. nördliche Breitengrad angenommen. In letzter Zeit wird ein scheinbar paradoxer Nordzug beobachtet, der Goldspechte in Gebiete führt, in denen sie dauerhaften Zugang zu Futterplätzen haben. So wurden Goldspechte seit 1984 regelmäßig bei der weihnachtlichen Vogelzählung in Saskatoon (52°N!) bei Außentemperaturen von bis zu minus 40° beobachtet. Die zurückgelegten Distanzen können beträchtlich sein: Ein Jungvogel, der im Juni im zentralen British Columbia beringt wurde, wurde Ende September auf Neufundland wiedergefunden, nach einer zurückgelegten Strecke von fast 5000 Kilometern.[15]
Schlechter untersucht sind die Zugtraditionen der Spechte der cafer-Gruppe. Die Rocky Mountains scheinen jedoch eine Zugscheide zu bilden, sodass diese Vögel in südliche oder südwestliche Richtungen abziehen. Außer diesen horizontalen Wanderungen werden vertikale Wanderbewegungen montaner Populationen beobachtet. Dies gilt auch für die Goldspechte Mexikos und Mittelamerikas.
Goldspechte ziehen in zuweilen sehr großen Gruppen; sie fliegen vor allem nachts.
Nahrung und Nahrungserwerb

Goldspechte ernähren sich selbst und ihren Nachwuchs hauptsächlich von Insekten. In bedeutend geringerem Maße werden verschiedene Samen, Früchte und Beeren aufgenommen. In der Insektennahrung spielen Ameisen und deren Entwicklungsstadien die größte Rolle. Goldspechte wählen die Ameisenart, die im Verbreitungsgebiet am häufigsten vorkommt und am energiesparendsten zu erreichen ist. Die Ameisennahrung kann bis zu 80 Prozent der Gesamtnahrungsmenge ausmachen. Daneben werden andere Insekten erbeutet, insbesondere Laufkäfer, Schnabelkerfe, Zweiflügler, Schmetterlinge, Springschrecken und Grillen. Selten werden Blattläuse, Würmer oder Schnecken aufgenommen. Regelmäßig fressen Goldspechte die Samen verschiedener Pflanzen wie die des Eichenblättrigen Giftsumach und anderer Sumachgewächse, Sonnenblumen- und Distelsamen, sowie verschiedene Beeren. Auch Eicheln gehören ins Nahrungsspektrum. Häufig besuchen Goldspechte Futterstellen. Goldspechte sind oft an mit Diatraea grandiosella befallenen Maispflanzen zu sehen, wo sie die Raupen dieser Mottenart erbeuten.[16]
Die kontinentalen Unterarten des Goldspechtes erbeuten den Großteil ihrer Nahrung auf dem Boden, während die beiden Inselrassen stärker baumgebunden sind. Auf dem Boden hüpfen die Spechte von Nahrungsquelle zu Nahrungsquelle, stochern und bohren im Boden, gelegentlich hämmern sie den Boden auf, um an Ameisennester zu gelangen. Sie wenden Blätter, Steine und Zweige, um an darunter lebende Insekten zu gelangen und stochern im Dung von Weidetieren. Baumbewohnende Insekten werden vor allem an absterbenden oder bereits toten Bäumen gesucht. Auch hier sind Stochern und Ablesen die Hauptstrategien, eher selten werden tiefer liegende Bohrgänge durch Hämmern freigelegt. Früchte und Beeren pflücken Goldspechte von den Zweigen oder picken heruntergefallene vom Boden auf. Gelegentlich jagen sie Insekten im Fluge.
Verhalten
Aktivität
Wie alle Echten Spechte ist der Goldspecht tagaktiv. Er verlässt seinen Schlafplatz kurz vor Sonnenaufgang und sucht ihn kurz nach Sonnenuntergang wieder auf. Als Schlafplatz dient eine Vielfalt an geschützten Stellen, wie Baumhöhlen, alte eigene oder fremde Spechthöhlen, Dachgiebel in Holzbauten, gelegentlich auch nur ein Stammabschnitt unter einem mächtigen Hauptast. Seine Tagesaktivität verbringt er außerhalb der Brutzeit zu etwa zwei Drittel auf Bäumen und nutzt diese Zeit nur zu einem Bruchteil zur Nahrungssuche. Den Rest der Zeit döst er, beobachtet die Umgebung oder putzt sein Gefieder. Das übrige Drittel verbringt er auf dem Boden, hier fast ausschließlich mit der Nahrungssuche beschäftigt.[17] Während der Zugzeit verbringen die ziehenden Populationen die Nächte fliegend, die Tage etwa zu gleichen Teilen mit Dösen, Ruhen, Gefiederpflege und Futtersuche. In den Wintermonaten wird verhältnismäßig mehr Zeit der Nahrungssuche gewidmet.
Agonistisches Verhalten
Die innerartliche Aggression ist beim Goldspecht stark reduziert. Er verteidigt nur den Brutplatz und den Partner, Nahrungsrivalität scheint kaum zu bestehen. Die Auseinandersetzungen sind stark ritualisiert und führen sehr selten zu Berührungskämpfen. Wichtigstes Element ist der Flicker-Tanz. Dabei sitzen sich die Konkurrenten auf einem Ast gegenüber, Kopf und Schnabel sind leicht erhoben. Rhythmisch wird der Kopf gesenkt und gewendet, sodass die Schnabelspitze eine 8 beschreibt. Dabei werden Flügel und Schwanz gespreizt, um die Federsignale maximal zur Geltung zu bringen, und bei äußerster Erregung hüpft und schaukelt der Vogel auf der Stelle. Diese Duelle, die bis zu einer halben Stunde dauern können, werden immer von gleichgeschlechtlichen Rivalen ausgetragen; sie sind begleitet von lauten Wicka – Rufen. Wenn ein Rivale aufgibt, wird er vom anderen unter lautem Rufen verfolgt.

Einer Reihe von Höhlenkonkurrenten begegnet der Goldspecht in der Vorbrutzeit und der eigentlichen Brutzeit äußerst aggressiv, greift sie direkt an und versucht sie vom Nistbaum zu vertreiben. Wichtigster unter ihnen ist der eingeführte und sich sehr rasch ausbreitende Europäische Star. Die Verteidigungsintensität hängt von früher gemachten Erfahrungen mit dieser Art ab, sodass vor allem Erstbrüter häufig unterliegen. Höhlenkonkurrenten, denen der Goldspecht aggressiv begegnet sind weiters der Buntfalke und die Büffelkopfente. Eine ganze Reihe von Höhlenkonkurrenten sind gleichzeitig Prädatoren.
Brutbiologie
Balz und Paarbildung
Die Paarbildung beginnt im Februar in den südlichsten Brutgebieten und endet Mitte Mai in den nördlichsten. Insgesamt liegen zu ihrer Phänologie jedoch sehr wenige Daten vor. Hauptelement der Balz ist ein modifizierter, langsamer Flicker-Tanz, bei dem beschwichtigende Elemente die aggressiven überwiegen. Nähert sich ein Artgenosse, beginnt unmittelbar die aggressive, gleichgeschlechtliche Auseinandersetzung.[18] Weitere Balzelemente sind Höhlenzeigen, ritualisiertes Klopfen an geeigneten Nistbäumen und lange Rufreihen. Die Kopulationen geschehen auf einem Ast in der Nähe der Nisthöhle.
Goldspechte werden als Jährlinge geschlechtsreif. Sie führen meist eine monogame saisonale Partnerschaft. Familiengruppen können noch gemeinsam den Zug antreten oder im Winterquartier im losen Kontakt bleiben. Wiederverpaarungen letztjähriger Partner dürften nicht selten sein. Polyandrie kommt bei einem kleinen Prozentsatz der Weibchen vor. Diese legen in leicht gestaffelten Abständen Gelege in die Bruthöhlen zweier Männchen. Während sich das Weibchen beim ersten Gelege noch stärker beteiligt, obliegen Brut und Jungenaufzucht beim zweiten fast ausschließlich dem Männchen.[19]
Nisthöhle

Der Höhlenbau beginnt etwa Mitte Mai in den nördlicheren und entsprechend früher in den südlicheren Brutgebieten. Auf Cuba und Grand Caymann schon gegen Ende März. Die Wahl des Höhlenbaums obliegt dem Weibchen, die Hauptarbeit bei der Errichtung der Höhle leistet das Männchen. Die Art des Höhlenbaumes ist sehr variabel, in nördlichen und nordöstlichen Gebieten wird die Amerikanische Zitterpappel überproportional häufig ausgewählt. Die Mehrzahl der Nistbäume ist vorgeschädigt oder bereits tot, in lebenden Bäumen werden oft Astausbrüche oder andere Schädigungen ausgenutzt. Die Höhe ist äußerst variabel. Auf Nantucket, wo die dort vorherrschenden niederwüchsigen Bäume nur in tiefen Stammregionen ausreichende Durchmesser aufweisen, liegt die Durchschnittshöhe bei einem Meter, in alten Douglasienbeständen des westlichen Nordamerikas in über 11 Meter.[20] Bruthöhlen werden auch in unterschiedliche Holzkonstruktionen wie Telegraphenmasten oder Holzbuhnen an der Küste geschlagen. Auch Nistkästen werden angenommen beziehungsweise adaptiert.
Da der Nisthöhlenbau Bestandteil des Paarbildungsprozesses ist, wird jedes Jahr mit der Errichtung einer neuen Bruthöhle begonnen, die aber später nicht unbedingt als tatsächliche Bruthöhle dienen muss. Oft werden alte Nistplätze wiederverwendet, oder letztjährig unvollendet gebliebene fertiggestellt. Die Dauer für die Errichtung einer neuen Brutstätte hängt von vielen Faktoren ab, unter günstigen Bedingungen liegt sie etwas unter zwei Wochen.[21] Auch die Arbeitsverteilung ist recht variabel, immer leistet das Männchen die Hauptarbeit.
Der Durchmesser des häufig leicht hochovalen Einfluglochs liegt im Durchschnitt bei 7–8,5 Zentimeter. Der Brutraum befindet sich durchschnittlich 40 Zentimeter unter dem Eingang; die Ausmaße der leicht konkaven Bodenfläche betragen etwa 165 Quadratzentimeter.[21]
Gelege und Brut
Goldspechte brüten einmal im Jahr. Nur bei frühem Gelegeverlust kommt es zu einer Zweitbrut. Früheste Gelege wurden Mitte April festgestellt, späteste, wahrscheinlich Zweitgelege, Anfang Juli. In Guatemala beginnt die Eiablage Anfang März. Für die beiden Inselrassen liegen keine Angaben vor. Die Gelege bestehen im Durchschnitt aus über 6 (4–9)Eiern, die im Tagesabstand gelegt werden. Gelege mit bis zu 12 Eiern wurden beobachtet, bei noch größeren dürfte Brutparasitismus vorliegen, der bei dieser Art generell relativ häufig aufzutreten scheint.[22] Die Eier sind weiß, leicht glänzend und messen im Durchschnitt 2,79 × 2,16 Zentimeter. Wie bei anderen Vögeln auch sind frühere Gelege größer als spätere. Die Gelege von Hybridpaaren sind meist kleiner.[23] Goldspechte beginnen vor dem vorletzten oder letzten Ei fest zu brüten, sodass das älteste Küken etwa zehn Tage nach Ablage des letzten Eis schlüpft, das jüngste maximal zwei Tage später. Beide Eltern brüten, das Männchen häufiger und immer während der Nacht. Die Nestlinge werden von beiden Eltern mit hervorgewürgter Insektennahrung versorgt. Sie nehmen in den ersten 10 Tagen sehr schnell an Gewicht zu, danach verlangsamen sich Wachstum und Gewichtszunahme. Nach 24–27 Tagen verlassen sie die Bruthöhle und oft gemeinsam mit ihren Eltern das unmittelbare Brutgebiet. Umfassende Daten zur Führungszeit liegen nicht vor, sie dürfte jedoch, wie Einzelbeobachtungen zeigen, kaum länger als drei Wochen dauern.[24] Auch die Jugenddispersion ist noch unzureichend erforscht. Die vorliegenden Untersuchungen weisen auf weiträumige Zerstreuungswanderungen hin.[25]
Zum Bruterfolg liegen nur wenige Daten vor. Eine größere Untersuchung von über 1000 Bruten in British Columbia lieferte folgende Ergebnisse: Aus 71 % der begonnenen Bruten flog zumindest ein Nestling aus. 16 % wurden durch verschiedene Nesträuber beendet, fast 6 % wegen andauernder Höhlenkonkurrenz vor allem durch Stare aufgegeben. Einige Bruten gingen durch Windbruch des Höhlenbaums verloren. Aus welchen Gründen die restlichen etwa 6 % scheiterten, blieb unklar.[26]
Lebenserwartung und Gefährdung
Wie bei den meisten Tierarten ist die Sterberate auch beim Goldspecht im ersten Jahr, insbesondere in den ersten Monaten nach dem Ausfliegen am größten. Etwa 43 Prozent der Goldspechte werden älter als ein Jahr. Danach verflacht die Mortalitätskurve etwas. Als Höchstalter wurden 9 Jahre und zwei Monate bei einem Specht aus der auratus-Gruppe festgestellt.[27]
Jungspechte verunglücken relativ häufig, indem sie gegen Hindernisse fliegen, sie werden häufiger Opfer von Prädatoren als ältere und überstehen Nahrungsengpässe oder anhaltende Schlechtwetterlagen schlechter als erfahrene, ältere Artgenossen. Unter den Prädatoren werden einige wie Eichhörnchen, Gleithörnchen, Waschbären und baumkletternde Schlangen nur den Eiern und Nestlingen gefährlich, andere, wie verschiedene Greifvögel, insbesondere der Habicht erbeuten adulte Goldspechte, oder überraschen, wie Marderarten, ruhende Spechte.
Bestandssituation und Bestandsentwicklung
Von der IUCN wird der Bestand der Art als ungefährdet betrachtet. Bei einem Verbreitungsgebiet von 15 Millionen Quadratkilometern wird der Gesamtbestand auf 16 Millionen Individuen geschätzt.[28]
Untersuchungen des BBS (Breeding Bird Survey) ergaben jedoch einen nicht unerheblichen Bestandsrückgang in weiten Teilen des Verbreitungsgebietes. Als Gründe dafür werden Habitatverlust verursacht insbesondere durch modernes Forstmanagement, die zunehmende Höhlenkonkurrenz mit dem in seinem Bestand rasch zunehmenden Europäischen Star und Pestizidkontamination in intensiv landwirtschaftlich genutzten Gebieten genannt. Über die Bestandssituation der beiden Inselrassen und der Unterart C. a. mexicanoides liegen keine Daten vor.
Trivia
Der Goldspecht ist der Staatsvogel von Alabama, das nach ihm auch Yellowhammer State genannt wird.
Literatur
- Karen L. Wiebe, William S. Moore: Northern Flicker (Colaptes auratus). In: A. Poole (Ed.): The Birds of North America Online. Cornell Lab of Ornithology, Ithaca 2008.
- Hans Winkler, David A. Christie und David Nurney: Woodpeckers. A Guide to the Woodpeckers, Piculets and Wrynecks of the World. Pica Press, Robertsbridge 1995, ISBN 0-395-72043-5, S. 146–147, 319–321.
Weblinks
- Colaptes auratus in der Roten Liste gefährdeter Arten der IUCN 2008. Eingestellt von: BirdLife International, 2008. Abgerufen am 2. Januar 2009.
- Videos, Fotos und Tonaufnahmen zu Colaptes auratus in der Internet Bird Collection
- Federn des Ostgoldspechts
Einzelnachweise
- Karen L. Wiebe, William S. Moore (2008) Introduction
- Karen L. Wiebe, William S. Moore (2008) Systematics; Measurements
- Jocelyn Hudon, Karen L. Wiebe, Elena Pini und Riccardo Stradi: Plumage pigment differences underlying the yellow-red differentiation in the Northern Flicker (Colaptes auratus). In: Comparative Biochemistry and Physiology, Part B 183 (2015) 1–10.
- Jocelyn Hudon, Robert J. Driver, Nathan H. Rice, Trevor L. Lloyd-Evans, Julie A. Craves und Daniel P. Shustack: Diet explains red flight feathers in Yellow-shafted Flickers in eastern North America. In: The Auk, Volume 134, 2017, SS 22–33; doi:10.1642/AUK-16-63.1
- Karen L. Wiebe, William S. Moore (2008) Sounds
- Karen L. Wiebe, William S. Moore (2008) Appearance
- Winkler et al. (1995) S. 307–326
- Brett W. Benz, Mark B. Robbins, A. Townsend Peterson: Evolutionary history of woodpeckers and allies (Aves: Picidae): Placing key taxa on the phylogenetic tree. In: Molecular Phylogenetics and Evolution 40 (2006): S. 389–399; S. 395 ff.
- Juian P. Hume und Richard Walters: Extinct Birds. Poyser-London 2012. S. 224. ISBN 978-1-4081-5725-1
- del Hoyo, J., Collar, N. & Christie, D.A. (2016). Red-shafted Flicker (Colaptes cafer). In: del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A. & de Juana, E. (eds.). Handbook of the Birds of the World Alive. Lynx Edicions, Barcelona. (abgerufen auf http://www.hbw.com/node/467472 on 13 October 2016).
- Winkler, H., Christie, D.A. & Kirwan, G.M. (2016). Yellow-shafted Flicker (Colaptes auratus). In: del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A. & de Juana, E. (eds.). Handbook of the Birds of the World Alive. Lynx Edicions, Barcelona. (abgerufen auf http://www.hbw.com/node/56268 on 13 October 2016).
- D. T. Tyler Flockhart und Karen L. Wiebe: Absence of Reproductive Consequences of Hybridization in the Northern Flicker (Colaptes auratus) Hybrid Zone In: The Auk April 2009 : Vol. 126, Issue 2 (Apr 2009), SS. 351–358.
- Karen L. Wiebe, William S. Moore (2008) Distribution
- Karen L. Wiebe, William S. Moore (2008) Behavior
- Karen L. Wiebe, William S. Moore (2008) Migration
- Karen L. Wiebe, William S. Moore (2008) Food Habits
- Karen L. Wiebe, William S. Moore (2008) Behavior/Daily Time Budget
- Karen L. Wiebe, William S. Moore (2008) Courtship and Agonistic Displays
- Karen L. Wiebe, William S. Moore (2008) Mating System And Sex Ratio
- Karen L. Wiebe, William S. Moore (2008) Breeding
- Karen L. Wiebe, William S. Moore (2008) Breeding~Nest
- Karen L. Wiebe, William S. Moore (2008) Eggs~Clutch Size
- Winkler et al. (1995) S. 321
- Karen L. Wiebe, William S. Moore (2008) Demography and Population
- Karen L. Wiebe, William S. Moore (2008) Range
- Karen L. Wiebe, William S. Moore (2008) Annual And Lifetime Reproductive Success
- Karen L. Wiebe, William S. Moore (2008) Life Span And Survivorship
- Datenblatt BirdLife international (PDF) engl.