Burmanniaceae
Die Burmanniaceae sind eine Pflanzenfamilie aus der Ordnung der Yamswurzelartigen (Dioscoreales). Die Familie umfasst rund einhundert Arten in neun häufig monotypischen beziehungsweise sehr artenarmen Gattungen. Die stammesgeschichtlich alte Familie ist weltweit in den Tropen verbreitet, ihre Arten sind mehrheitlich vollständig ohne Blattgrün und ernähren sich stattdessen mykoheterotroph als Parasiten von Pilzen. Durch ihre Unabhängigkeit vom Licht als Energiequelle erschließen sie sich ökologische Nischen, die Pflanzen sonst weitgehend verschlossen bleiben.
| Burmanniaceae | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Gymnosiphon bekensis in Kamerun | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Burmanniaceae | ||||||||||||
| Blume |
Aufgrund der evolutionären Anpassungen an ihre Lebensweise wurde den blattgrünlosen Arten „ein bizarres Aussehen“[1] attestiert und sie wurden mit „einer künstlichen Flora aus Glas, in der die kleinen Blüten in brillanten Farben wie Kristalle auf den zarten, glasartigen Stängeln ruhen“ verglichen.[2]
Beschreibung
Erscheinungsbild
Alle Arten der Familie sind mykotrophe und – bis auf rund vierzig Arten der Gattung Burmannia – chlorophylllose, einjährige oder ausdauernde krautige Pflanzen. Sie erreichen meist Wuchshöhen von 5 bis 30 Zentimetern, einige Burmannia-Arten nur bis zu 2 Zentimetern, Höchstwerte sind 75 oder gar 100 Zentimeter (Burmannia disticha). Fast alle Arten wachsen terrestrisch, selten finden sich Epiphyten (Burmannia kalbreyeri, Burmannia longifolia).
Rhizom und Wurzeln
Das knollenähnliche, zylindrische und üblicherweise vertikal wachsende Rhizom ist verzweigt, nur wenige Zentimeter lang und dicht bedeckt mit überlappenden Schuppen und fadenförmigen, exogen gewachsenen Wurzeln, die von Mykorrhizen durchwachsen sind. Wurzelhaare fehlen, finden sich aber in der Gattung Dictyostega möglicherweise funktionell durch die Ränder der Rhizomschuppen beziehungsweise lange Härchen an deren Spitze ersetzt. Wurzelhauben fehlen den mykoheterotrophen Arten. Die Wurzeln besitzen eine ausgeprägte Endodermis mit U-förmigen Wandverdickungen und/oder Casparyschen Streifen. Der Zentralzylinder ist schwach tri- oder tetrarch ausgebildet und besitzt nur ein bis sechs Xylem-Gefäße mit ring- oder schraubenförmigen Wandverstärkungen und drei bis acht Phloemstränge. Es wurden keine Siebröhren-Plastiden nachgewiesen. Das Perizykel ist fast vollständig reduziert.
Im Zentrum der Rhizome findet sich Markgewebe sowie der – gelegentlich von einer Endodermis umgebene – Leitbündelzylinder, die Rinde ist im Vergleich zum oberirdischen Teil der Pflanze verdickt. Sowohl Markgewebe als auch Rinde dienen als Stärkespeicher.
Mittels seitlich aus dem Rhizom wachsender Sprosse beziehungsweise von Achsenknospen aus der Wurzel vermögen sich Arten der Burmanniaceae auch vegetativ zu vermehren. Von zwei Arten der Burmannia (Burmannia alba, Burmannia longifolia) ist Wurzeldimorphismus bekannt, neben einem eigentlich schwach ausgeprägten Wurzelwerk findet sich hier noch eine, gelegentlich sogar zwei sehr verdickte, knollenartige Wurzel.
Von diesem Aufbau weichen die blattgrünen Arten der Gattung Burmannia meist ab: ihnen fehlt das Rhizom und sie besitzen Wurzelhauben. Sie zählen mit bis zu 70 Zentimeter Wuchshöhe auch zu den größten Vertretern der Familie.
Sprossachsen
Oberirdische Sprossachsen werden nur in Form der Blütenstandsachsen ausgebildet, sie sind in der Regel unverzweigt und im Querschnitt zylindrisch, um ihren Ansatz findet sich eine von der Hauptwurzel ausgehende, rund 1 Millimeter hohe Scheide. Die Epidermis besitzt so gut wie keine Spaltöffnungen, wenn, dann ohne Nebenzellen (anomocytische Stomata). Nach innen folgen ein ausgeprägtes subepidermales Gewebe, eine Sklerenchymscheide, eine Endodermis ähnlich der der Wurzeln. Es gibt 3 bis 20 reduzierte kollaterale Leitbündel. Die Xylem-Gefäße besitzen ring- und spiralförmige Verdickungen.
Blätter
Die wechselständig angeordneten, ungestielten Blätter sind bei mykotrophen Arten zu kleinen Schuppenblättern reduziert und von blass bräunlicher, gelblicher, rötlicher Farbe, selten auch vollständig farblos. Bei blattgrünen Arten sind sie bis zu 50 Zentimeter lang, linealisch bis lanzettlich und bilden in der Regel eine Rosette am unteren Teil der Sprossachse. Nebenblätter fehlen. Die Blätter besitzen nur ein kollaterales Leitbündel, das aus wenigen Tracheiden, selten aus Gefäßen besteht. Für Burmannia fadouensis und Burmannia pingbianensis ist die Bildung von Achselbulbillen zum Zweck der vegetativen Vermehrung berichtet.[3]
Blütenstände und Blüten

In den endständigen Blütenständen sind einfache (zum Beispiel bei Apteria) oder doppelte (Gymnosiphon, Dictyostega) Wickel in denen ein bis fünfzehn gestielte Blüten über jeweils einem Tragblatt stehen. Im Falle von Doppelwickeln sind die Blüten einseitswendig ausgerichtet. Die Blütenstände sind bei Gymnosiphon und Dictyostega eher locker, bei Miersiella und einigen Burmannia kompakt aufgebaut. Die Länge der Blütenstiele ist, insbesondere innerhalb der Gattung Gymnosiphon von großer taxonomischer Bedeutung. Die in der Regel radiärsymmetrischen und röhren- bis trompetenförmigen Blüten sind dreizählig. Die Blütenfarbe ist variabel, häufig von blauer bis hellblauer Grundfarbe, daneben aber auch grünlich, gelb, weiß oder rosafarben. Oft sind verschiedene Farben kontrastreich miteinander kombiniert.
Die vollständig miteinander verwachsenen Tepalen stehen in zwei Kreisen, die äußeren drei sind deutlich größer als die inneren, nur bei Campylosiphon ist dieser Unterschied kaum ausgeprägt. Bei Marthella fehlen die Tepalen des inneren Kreises vollständig. Die Tepalen des äußeren Kreises sind dabei meist einfach, (nur bei den Gattungen Cymbocarpa und Gymnosiphon dreigelappt); die des inneren Kreises sind stark verkleinert, einfach (bei Hexapterella gelegentlich dreigelappt) und fleischig verdickt. In der Knospe umschließen die bündig aneinander stehenden äußeren Tepalen die inneren vollständig. Der Perianth ist persistent, wird also bis zur Fruchtreife nicht abgeworfen, sondern vertrocknet langsam; Ausnahmen sind Gymnosiphon, Cymbocarpa und Hexapterella gentianoides, die als circumscissil beschrieben werden, hier werden also alle oberhalb der Blütenröhre gelegenen Blütenelemente abgeworfen und hinterlassen eine „nackte Blütenröhre“.

Es ist nur ein Kreis mit drei, den inneren Tepalen gegenüber stehenden Staubblättern vorhanden, der äußere Staubblattkreis fehlt. Staubfäden fehlen mit Ausnahme der Gattungen Apteria und Marthella sowie bei Hexapterella gentianoides. Die Theken reißen quer auf und sind deutlich voneinander entfernt. Der Griffel ist ebenso lang wie die Blüte, im Querschnitt am Ansatz dreieckig bis rund, oberhalb zylindrisch, fadenförmig und an der Spitze auf Höhe der Staubbeutel in drei trichterförmige oder zweilippige Narbenäste verzweigt. Bei zweilippigen Narbenästen ist die Unterlippe mit Papillen besetzt, die als Haftpunkte für die Pollen dienen.
Die drei Fruchtblätter sind zu einem unterständigen Fruchtknoten verwachsen. Der Fruchtknoten besitzt ein oder drei Fächer. Bei drei Fächern (Burmannia) ist die Plazentation zentralwinkelständig, bei einem meist parietal. Die Samenanlagen sind anatrop, bitegmisch und tenuinucellat.
Für zahlreiche Mitglieder der Familie sind Nektarien nachgewiesen, von einfachen Septalnektarien bei Burmannia und Campylosiphon bis hin zu hochkomplexen Drüsenstrukturen bei Miersiella umbellata und Marthella trinitatis, wo sechs je paarweise miteinander angeordnete, verwachsene Drüsen oberhalb der Fruchtknoten platziert sind.[4]
Düfte werden nur schwach, vereinzelt oder kurzzeitig abgegeben, ein einheitliches Bild lässt sich nicht zeichnen. Einzelne Berichte liegen unter anderem vor für Burmannia championii (süßlich), Hexapterella gentianoides (süß), Campylosiphon purpurascens (wohlriechend), Gymnosiphon divaricatus (angenehm, primelähnlich) und eine weitere Gymnosiphon-Art (nach Kakao). Als Quelle der Düfte werden die Papillen der Perianthinnenseiten in Betracht gezogen.[5]
Pollen
Die Pollen werden als äußerst vielgestaltig beschrieben, nur sehr wenige Merkmale gelten für die gesamte Familie. Zumeist werden die Pollenkörner als Monaden, also einzeln ausgestreut, selten bilden sie Gruppen aus je vier (Tetraden) oder zwei (Dyaden) Pollenkörnern (zum Beispiel bei Apteria aphylla). Bei einigen Gymnosiphon-Arten werden große Aggregationen zu gleicher Zeit ausgestreut (Massula). Die Oberfläche der sehr dünnen Exine ist vollkommen glatt, die Form des Korns ist ellipsoid, seine Länge beträgt zwischen 17 und 54 Mikrometer. An den Aperturen dünnt die Exine aus und die Intine tritt hervor. Meist sind die Pollen porat (wobei die Poren gelegentlich exzentrisch zum Äquator des Pollenkorns liegen), die Anzahl der Poren ist sehr variabel und kann selbst innerhalb einer Art schwanken. Monoporate und diporate Pollen überwiegen, gelegentlich gibt es Arten mit drei Poren. Selten finden sich inaperturate (Gymnosiphon, Apteria aphylla, Burmannia sphagnoides), noch seltener monocolpate Pollen (Cymbocarpa refracta, Hexapterella gentianoides).

Früchte und Samen
Die Kapselfrüchte sind ein- oder dreifächrig, öffnen sich längs (mit Ausnahme mehrerer quer verlaufender Schlitze bei den Burmannia) und enthalten viele winzige Samen, die bei den blattgrünen Arten durch Wind (Boleochorie), bei den mykoheterotrophen Arten durch Wasser verbreitet werden (Hydrochorie).[6]
Die Samen sind 0,19 bis 1,52 Millimeter lang, 0,06 bis 0,55 Millimeter breit, rundlich-oval bis länglich,[7] gelb bis braun, glänzend und ungeflügelt. Der Funiculus ist relativ lang, ein Endosperm ist vorhanden, aber nur extrem schwach ausgeprägt.[6]
Genetik
Die Chromosomenzahlen reichen von 6 und 8 über 68 bis zu 87 bis 99. Das Vorhandensein von Polyploidien wird angenommen, eine genaue Chromosomengrundzahl (möglicherweise 6 oder 8) ist aber nicht bekannt.
Ökologie
Bestäubung

Bislang wurde bei den Burmanniaceae nur Selbstbestäubung beobachtet. Sie tritt bei vielen Arten regelmäßig auf und wird durch mehrere Anpassungen erleichtert: Staubbeutel und Narbe sind einander genähert, die Antheren öffnen sich in der Knospe, teilweise wachsen Pollenschläuche aus der Anthere zur Narbe. Für Apteria aphylla existiert ein Bericht über Besuche von Milben, die allerdings aufgrund ihrer eingeschränkten Mobilität die Pflanzen auch nur mit ihrem eigenen Pollen bestäuben können. Bei einigen Burmannia-Arten findet sich Kleistogamie, die Blüte bleibt also geschlossen und es kommt zur Selbstbefruchtung (Burmannia championii, Burmannia lutescens, Burmannia capitata), bei Burmannia coelestis wurde auch Apomixis beobachtet, eine vollständig ungeschlechtliche Fortpflanzungsform.
Die Anpassungen der Blüten vieler Arten deuten jedoch auf Fremdbestäubung hin. Teilweise liegt Protandrie vor (die Staubbeutel reifen vor den Fruchtblättern, eine Selbstbestäubung wird so vermieden), die Blütenstruktur ist deutlich auf den Besuch durch Insekten angepasst, auch in Gestalt der Nektarproduktion vieler Arten. Als Bestäuber werden aufgrund der Blütenstruktur Schmetterlinge vermutet. In den Kronröhren dreier Exemplare von Gymnosiphon breviflorus, die als Untersuchungsmaterial dienten, wurde zweimal eine „Hymenoptere aus der Verwandtschaft der Erzwespen […], in der dritten Kronröhre derselben Art eine Fliegenpuppe (oder Larve) der Unterordnung Brachycera“ gefunden.[5]
Lebensweise
.jpg.webp)
Die Mehrheit aller Arten der Familie betreibt überhaupt keine Photosynthese mehr und bildet dementsprechend kein Chlorophyll. Stattdessen leben sie myko-heterotroph von arbuskulären Mykorrhizapilzen, sind also zu ihrer Ernährung vollständig von den Pilzen abhängig. Da diese Mykorrhiza-Pilze in Symbiose mit anderen Pflanzen stehen, meist Bäumen oder Sträuchern, wurde für diese Konstellation gelegentlich auch der Begriff Epiparasitismus gebraucht. Im strengen Sinne ist dieser Begriff jedoch unzutreffend, da Epiparasitismus als indirekte Miternährung am Pilzpartner ohne Schädigung des Pilzes selbst hier nicht vorliegt. Burmanniaceae sind echte Parasiten am Pilz: die in die äußeren bis zu 12 Zellschichten von Rinde und Epidermis der Wurzeln beziehungsweise Rhizome einwachsenden Hyphen des Pilzes werden nach und nach abgetötet und die Nährstoffe in Gestalt von Stärke im Gewebe eingelagert.[8] Über lange Zeiträume sind die unterirdischen Pflanzenteile (Rhizom, Wurzeln) so die einzigen aktiven Teile der Pflanzen, sie treiben erst aus, wenn sie vom Pilz hinreichende Reserven an Nährstoffen angesammelt haben.
Wenngleich allerdings der größte Teil der Arten der Burmannia blattgrün ist, so bedeutet dies doch keinesfalls, dass sie autotroph sein müssen. Auch sie parasitieren oftmals Pilze, betreiben aber parallel Photosynthese, nur wenige Arten wie Burmannia disticha sind völlig frei von Mykorrhizen. Leake konstatierte diesbezüglich: „Es ist höchstwahrscheinlich, dass viele dieser Arten mehr Kohlenstoff durch Heterotrophie als durch Photosynthese beziehen […]“ („It is most likely that many of these species obtain more carbon heterotrophically than they do from photosynthesis […]“)[9]
Die von den Pflanzen parasitierten Pilze entstammen – soweit bekannt – der Klasse Glomeromycetes, darunter neben Arten der Gattung Glomus auch einige aus der Familie der Acaulosporaceae,[10][11] sowie der Peronosporaceae.[12]
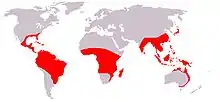
Verbreitung
Die Burmanniaceae sind weltweit in den tropischen Zonen aller Kontinente zu finden und erreichen nur selten warm gemäßigte Breiten. Besondere Verbreitungsschwerpunkte sind Südamerika und Asien (Südostasien, Malesien), ein kleinerer Schwerpunkt ist West- und Zentralafrika. Die Familie strahlt darüber hinaus nach Nordamerika, die Karibik und Ostaustralien aus. Die Verteilung zwischen Paläotropis und Neotropis ist annähernd gleich.
Die fast weltweite Verbreitung ist zusammen mit der Tatsache, dass keine Art auf mehr als einem Kontinent zu finden ist, ein deutliches Anzeichen für das Alter der Familie, das mit rund 93 Millionen Jahren für den ältesten gemeinsamen Vertreter der rezenten Arten (Crown node age) angegeben wird.[13] Bemerkenswert ist daneben besonders der hohe Endemismusgrad auf Gattungsebene: Während die 14 Arten in Afrika und 39 Arten in Australasien nur drei Gattungen entstammen (Burmannia, Gymnosiphon, Saionia), sind die 47 neuweltlichen Arten auf alle 9 Gattungen der Familie verteilt.[14]

Habitate
Die myko-heterotrophen Arten der Burmanniaceae wachsen meist auf verrottendem Holz oder Laub an sehr schattigen Standorten in luftfeuchten Primärwäldern, gelegentlich aber auch an sonnigen, offenen Standorten humusreicher Feuchtsavannen, in Bambusgebüschen, Kiefernwäldern, Mangroven oder sogar in feuchtem Sand in Höhlen.
Die blattgrünen Burmannia-Arten finden sich an schattigen, aber offenen, feuchten bis sumpfigen und humusarmen Standorten in Savannen, Prärien, Feuchtwiesen, Torfmoos-Mooren, an den Ufern von Teichen und Flüssen, in Kiesgruben, Zypressensümpfen oder Nadelwäldern, aber auch Reisfeldern.
Vergesellschaftet sind Burmanniaceae mit Arten der Süßgräser, Binsengewächse, Sauergrasgewächse, Xyridaceae, Eriocaulaceae, Sonnentaugewächse, Wasserschlauchgewächse, Enziangewächse, Kreuzblumengewächse und anderen, in Westafrika sind sie Charakterpflanzen der Inselberg-Vegetation. Die epiphytischen Arten leben in Wäldern auf lebenden Ästen oder den Stelzwurzeln von Palmen der Gattung Iriartea.
Die Burmannia-Arten wachsen üblicherweise einzeln und finden sich mehrheitlich in niedrigen Höhenlagen, können aber auch auf Höhenlagen von bis zu 3600 Metern vordringen.
Gefährdung und Status
Aufgrund der meist unzugänglichen Standorte, ihres disjunkten Verbreitungsgebietes, ihrer Unauffälligkeit und ihrer natürlichen Seltenheit ist keine summarische Aussage über die Entwicklung der Bestände der Familie möglich. Auf der Roten Liste der IUCN stehen drei Arten der Gattung Gymnosiphon, vier Arten der Gattung Afrothismia, vier oder fünf Arten der Gattung Burmannia und eine Art der Gattung Oxygyne.[15] In den USA ist Burmannia flava in Florida seit 1998 als „Endangered“ = „Gefährdet“ eingestuft.[16]
In besonderer Hinsicht unklar ist der Status von Marthella trinitatis, der einzigen Art ihrer Gattung; sie ist nur ein einziges Mal 1883 auf dem El Tucuche, dem zweithöchsten Berg Trinidads, gesammelt und seither nicht mehr wiedergefunden worden, einen offiziellen Status genießt sie aber nicht. Ähnliches gilt für die bisher nur dreimal gefundene Cymbocarpa saccata.
Systematik
Gattungen
Die Burmanniaceae umfassen derzeit (unter Einschluss der Thismiaceae) etwa 16 Gattungen mit rund 170 Arten, neue Arten (insbesondere der Gattung Burmannia) werden jedoch regelmäßig beschrieben. Bis auf Afrithismia mit etwa 14 Arten, Gymnosiphon mit etwa 30 Arten, Burmannia mit etwa 60 Arten und Thismia mit etwa 64 Arten haben alle Gattungen nur einen bis vier Vertreter.
- Afrothismia Schltr.: Hierher gehören etwa 14 Arten, die in Afrika vorkommen.[17] Sie werden von manchen Autoren auch in die Familie der Thismiaceae gestellt.
- Apteria Nutt.: Sie umfasst nur eine Art:
- Apteria aphylla (Nutt.) Barnhart ex Small: Sie kommt im tropischen und subtropischen Amerika vor.[17]
- Burmannia L. (Syn.: Tripterella Michx.): Hierher gehören etwa 60 Arten.[17]
- Campylosiphon Benth.: Hierher gehören zwei Arten.[17]
- Cymbocarpa Miers: Hierher gehören zwei Arten:[17]
- Dictyostega Miers: Mit nur einer Art, die sich in drei Unterarten gliedert:[17]
- Dictyostega orobanchoides (Hook.) Miers: Sie kommt im tropischen Amerika vor.[17]
- Geomitra Becc.: Sie enthält nur eine Art:
- Geomitra clavigera Becc.: Sie kommt in Thailand, Malaysia, Sumatra und Borneo vor.[17]
- Gymnosiphon Blume: Mit etwa 30 Arten in den Tropen.[17]
- Haplothismia Airy Shaw: Sie enthält nur eine Art:[17] Sie wird von manchen Autoren auch in die Familie der Thismiaceae gestellt.
- Haplothismia exannulata Airy Shaw: Sie kommt im südlichen Indien vor.[17]
- Hexapterella Urb.: Hierher gehören zwei Arten, die in der Neotropis vorkommen.[17]
- Marthella Urb.: Mit nur einer Art:
- Marthella trinitatis (Johow) Urb.: Sie kommt im nördlichen Trinidad vor.[17]
- Miersiella Urb.: Mit nur einer Art:
- Miersiella umbellata (Miers) Urb.: Sie kommt im tropischen Südamerika vor.[17]
- Oxygyne Schltr.: Sie enthält vier Arten, die von Kamerun bis zur Zentralafrikanischen Republik vorkommen.[17] Sie werden von manchen Autoren auch in die Familie der Thismiaceae gestellt.
- Saionia Hatus.: Sie wurde 2015 aufgestellt. Sie werden von manchen Autoren auch in die Familie der Thismiaceae gestellt. Ihre drei Arten kommen nur im südlichen Japan bis zu den Nansei-Inseln vor.[17][18]
- Thismia Griff.: Hierher gehören etwa 64 Arten.[17] Sie werden von manchen Autoren auch in die Familie der Thismiaceae gestellt.
- Tiputinia P.E.Berry & C.L.Woodw.: Sie umfasst nur eine Art. Sie wurde 2007 erstbeschrieben.[17] Sie wird von manchen Autoren auch in die Familie der Thismiaceae gestellt.
- Tiputinia foetida P.E.Berry & C.L.Woodw.: Sie kommt im östlichen Ecuador vor.[17]
Geschichte der Systematik

Die Geschichte der Systematik der Burmanniaceae ist in vieler Hinsicht geprägt von Unsicherheiten aufgrund der teils extrem reduzierten Morphologie ihrer mykoheterotrophen Vertreter.
Die Familie der Burmanniaceae wurden 1830 von Blume mit den drei Gattungen Burmannia, Gymnosiphon und Gonyanthes (letztere wurde später als Synonym zu den Burmannia gestellt) aufgestellt, nachdem Linné die beiden von ihm bekannten Burmannia-Arten Burmannia disticha und Burmannia biflora bei ihrer Erstveröffentlichung 1753 der Ordnung Monogynia (Klasse Hexandria) zurechnete (Familien als Taxa wurden in Linnés System ursprünglich nicht verwendet).[19] Der Gattungsname Burmannia ehrt den niederländischen Botaniker und Arzt Johannes Burman, einen Freund Linnés. Holotyp ist ein Exemplar von Burmannia disticha aus Ceylon, heute verwahrt im Schwedischen Museum für Naturgeschichte.[20]
Einer ersten Revision wurde die Familie 1847 bis 1851 durch Miers unterzogen. Er führte zugleich eine weitere Untergliederung der Familie in zwei Unterfamilien ein, die Thismiae und die Burmanniae, die mit teils variierenden Rangstufen noch bis zur Jahrtausendwende vertreten wurde (siehe zum Beispiel Maas-van de Kamer 1998).
Hooker und Bentham stuften die Familie 1883 anhand morphologischer Merkmale wie Mykotrophie, unterständigem Fruchtknoten, sehr kleinen Samen und einem – wie bei den Orchideen – als fehlend angenommenen Endosperm gemeinsam mit den Orchideen in der Ordnung Microspermae ein und gliederten sie in drei Tribus, da sie auch noch die Corsiaceae in die Familie eingliederten. Diese Zusammenfassung war bis zum Erscheinen von Jonkers Monografie 1938 maßgeblich, auch Engler übernahm sie, zählte aber darüber hinaus auch noch die monotypische Gattung Geosiris zur Familie.
Die scheinbare Verwandtschaft mit den Orchideen findet sich noch bei Cronquist wieder, der die Burmanniaceae in die Ordnung Orchidales eingliederte. Armen Tachtadschjan hingegen platzierte die Burmanniaceae, ebenso wie Robert Folger Thorne, mit den Corsiaceae in einer eigenen Ordnung Burmanniales.[21]
Bedeutende Beiträge zur Kenntnisse der Familie nach Jonker leisteten insbesondere das Ehepaar Hiltje Maas-van de Kamer und Paul J. M. Maas von der Universität Utrecht, Traudel Rübsamen und in neuester Zeit Dianxiang Zhang, Vincent Merckx und Peter Schols.
Phylogenetik
Anhand molekulargenetischer Untersuchungen wurden all die zuvor geschilderten Ansätze zunehmend in Frage gestellt. Weder Thismiae noch Corsiae konnten als Teil der Burmanniaceae bestätigt werden, zu den Corsia als Teil der Liliales besteht wohl nicht einmal mittelbar eine Verwandtschaft.[22][23][24] Seit Anfang des 21. Jahrhunderts werden alle Tribus daher als separate, einander auch nicht unmittelbar nahestehende Familien begriffen. Die Angiosperm Phylogeny Website führt die Burmanniaceae als Teil der Dioscoreales, mit den Dioscoreaceae als Schwestergruppe.[25]
Auch die innere Systematik der bisherigen Burmanniae, die als einziges Mitglied der Familie verbleiben, ist den Untersuchungen zufolge revisionsbedürftig. So sind die Burmannia paraphyletisch.[22] Auch gilt der Status der Gattung Cymbocarpa als zweifelhaft, sie ist eventuell in die Gattung Gymnosiphon einzugliedern.
Die derzeit umfangreichste und neueste Untersuchung der Familie von Merckx[24] gibt die Verhältnisse in der Familie wie folgt wieder (stark vereinfacht, fettgedruckte Namen repräsentieren mehrere Taxa):
| |||||||||||||||||||||||||||||||||||||||||||
Im Rahmen der Untersuchungen zeigte sich auch, dass der Verlust des Chlorophylls sich mehrfach unabhängig voneinander eingestellt hat. Dies wird interpretiert als Hinweis darauf, dass generell eine grundsätzliche genetische Disposition innerhalb der Familie vorhanden ist, die eine Umstellung auf eine mykotrophe Lebensweise erlaubt.
Verwendung
Für den Menschen sind die Arten der Burmanniaceae weitgehend bedeutungslos. Die nach vereinzelten Proben als adstringierend und bitter schmeckend geschilderten[26] Arten Apteria aphylla und Burmannia biflora wurden als Teepflanzen verwandt, für Burmannia coelestis wurde von den Santal in Westbengalen ein Gebrauch als Heilkraut berichtet.[27]
Literatur
Die Informationen dieses Artikels entstammen zum größten Teil den folgenden Quellen, für weitere Angaben siehe unter Einzelnachweise:
- Hiltje Maas-van de Kamer: Burmanniaceae. In: Klaus Kubitzki (Hrsg.): The Families and Genera of Vascular Plants. Volume 3, Berlin 1998, ISBN 3-540-64060-6.
- Traudel Rübsamen: Morphologische, embryologische und systematische Untersuchungen an Burmanniaceae und Corsiaceae (Mit Ausblick auf die Orchidaceae-Apostasioideae). 1986, ISBN 3-443-64004-4.
- Dianxiang Zhang: Systematics of Burmannia L. (Burmanniaceae) in the Old World. In: Hong Kong University Theses Online, Thesis (Ph. D.). University of Hong Kong, 1999.
- Fredrik Pieter Jonker: A monograph of the Burmanniaceae. In: Meded. Bot. Mus. Herb. Rijks Univ. Utrecht, Band 51, 1938, S. 1–279.
- P. J. M. Maas, H. Maas-van de Kamer, J. van Bentham, H. C. M. Snelders, T. Rübsamen: Burmanniaceae. In: Flora Neotropica, Monogr. 42, 1986, S. 1–189.
Einzelnachweise
- Traudel Rübsamen: Morphologische, embryologische und systematische Untersuchungen an Burmanniaceae und Corsiaceae (Mit Ausblick auf die Orchidaceae-Apostasioideae), 1986, S. 9.
- R. Ciferri: L’habitat e la micorrizia di alcune Burmanniacee della Repubblica Dominicana. In: Atti Ist. Bot. „Giovanni Brosi“, Ser. V, 7, S. 25–34, Übersetzung zitiert nach: Traudel Rübsamen: Morphologische, embryologische und systematische Untersuchungen an Burmanniaceae und Corsiaceae (Mit Ausblick auf die Orchidaceae-Apostasioideae), 1986, S. 9.
- Dianxiang Zhang: Systematics of Burmannia L. (Burmanniaceae) in the Old World. In: Hong Kong University Theses Online, Thesis (Ph.D.), University of Hong Kong, 1999, S. 16–17.
- Traudel Rübsamen: Nectaries of the Burmanniaceae (Burmannieae). In: Acta Bot. Neerl., Volume 32, Issue 4, 1983, S. 351.
- Traudel Rübsamen: Morphologische, embryologische und systematische Untersuchungen an Burmanniaceae und Corsiaceae (Mit Ausblick auf die Orchidaceae-Apostasioideae), 1986, S. 90–91, Tafel 75.
- J. H. Kirkbride Jr., C. R. Gunn, M. J. Dallwitz: Burmanniaceae In: Family Guide for Fruits and Seeds, Version 1.0, 2006, Zugriff: 12. August 2007, (beim weblinkarchiv. (Memento vom 19. Januar 2008 im Internet Archive))
- Traudel Rübsamen: Morphologische, embryologische und systematische Untersuchungen an Burmanniaceae und Corsiaceae (Mit Ausblick auf die Orchidaceae-Apostasioideae), 1986, Tabelle XVII, S. 141.
- Stephan Imhof: Subterranean structures and mycotrophy of the achlorophyllous Dictyostega orobanchoides (Burmanniaceae). In: Revista de Biología Tropical, Volume 49, No. 1, 2001, Online, Zugriff am 3. September 2007
- Jonathan R. Leake: The Biology of Myco-Heterotrophic ('Saprophytic') Plants. In: New Phytologist, Volume 127, Issue 2, 1994, S. 179.
- Thassilo Franke, Ludwig Beenken, Matthias Döring, Alexander Kocyan, Reinhard Agerer: Arbuscular mycorrhizal fungi of the Glomus-group A lineage (Glomerales; Glomeromycota) detected in myco-heterotrophic plants from tropical Africa. In: Mycological Progress, 2006, Vol. 5, p. 28
- Jonathan R. Leake: Plants parasitic on fungi: unearthing the fungi in myco-heterotrophs and debunking the ‚saprophytic‘ plant myth. In: Mycologist Volume 19, 2005, S. 113–122.
- Dianxiang Zhang: Systematics of Burmannia L. (Burmanniaceae) in the Old World, in: Hong Kong University Theses Online, Thesis (Ph.D.), University of Hong Kong, 1999, S. 13.
- T. Janssen, K. Bremer: The age of major monocot groups inferred from 800+ rbcL sequences. In: Bot. J. Linnean Soc., Band 146, 2004, S. 385–398.
- Dianxiang Zhang: Systematics of Burmannia L. (Burmanniaceae) in the Old World. In: Hong Kong University Theses Online, Thesis (Ph.D.), University of Hong Kong, 1999, Tabelle 1–1, S. 30.
- Suche nach „Burmanniaceae“ in der Roten Liste gefährdeter Arten der IUCN.
- Siehe Eintrag im Atlas of Florida Vascular Plants, Online, Zugriff am 30. September 2007
- Rafaël Govaerts (Hrsg.): Burmanniaceae. In: World Checklist of Selected Plant Families (WCSP) – The Board of Trustees of the Royal Botanic Gardens, Kew, abgerufen am 24. Juni 2018.
- Hiroyoshi Ohashi: Resurrection of Saionia (Thismiaceae). In: Journal of Japanese Botany. Volume 90, Issue 2, 2015, S. 116.
- Carolus Linnaeus: Species Plantarum, Band 1, S. 287, 1753 (botanicus.org)
- Eine Abbildung des Holotyps findet sich auf der Website des Schwedischen Museums für Naturgeschichte
- Übersicht über die verschiedenen Platzierungen nach Flowering Plant Gateway: Vascular Plant Family Finder, Online, Zugriff am 25. September 2007.
- Dianxiang Zhang: Phylogenetic reconstruction of Burmannia L. (Burmanniaceae): a preliminary study. In: Acta Phytotaxonomica Sinica, 2001, Volume 39, 3, S. 203–223.
- Ray Neyland: A phylogeny inferred from large-subunit (26 S) ribosomal DNA sequences suggests that Burmanniales are polyphyletic. In: Australian Systematic Botany, Volume 15, 2002, S. 19–28
- V. Merckx, P. Schols, H. Maas-van de Kamer, P. Maas, S. Huysmans, E. Smets: Phylogeny and evolution of Burmanniaceae (Dioscoreales) based on nuclear and mitochondrial data. In: Am. J. Bot., Volume 93, 2006, S. 1684–1698.
- Siehe dazu das Kladogramm zu den Dioscoreales auf der Angiosperm Phylogeny Website.
- Traudel Rübsamen: Morphologische, embryologische und systematische Untersuchungen an Burmanniaceae und Corsiaceae (Mit Ausblick auf die Orchidaceae-Apostasioideae), 1986, S. 56.
- Fredrik Pieter Jonker: A monograph of the Burmanniaceae, In: Meded. Bot. Mus. Herb. Rijks Univ. Utrecht, Band 51, 1938, S. 14.
{kind=link}