Metriacanthosauridae
Die Metriacanthosauridae (Syn. Sinraptoridae) sind eine Gruppe carnivorer, theropoder Dinosaurier im Rang einer Familie innerhalb der Überfamilie der Allosauroidea. Die Gruppe lässt sich vom Mitteljura bis zum Oberjura und möglicherweise noch bis weit in die Unterkreide nachweisen.
| Metriacanthosauridae | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Schädel von Sinraptor hepingensis | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Mitteljura bis Oberjura (Unterkreide?) | ||||||||||||
| 174,1 bis 145 (126,3?) Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Metriacanthosauridae | ||||||||||||
| (Paul, 1988) |
Definition
Die Metriacanthosauridae (Paul, 1988) sind definiert als die umfassendste Klade, die Metriacanthosaurus parkeri enthält aber nicht Allosaurus fragilis, Carcharodontosaurus saharicus oder Passer domesticus.[1][2]
Historisches
Die Typusgattung der Metriacanthosauridae wurde 1923 durch Friedrich von Huene zunächst als Vertreter der Gattung Megalosaurus (Megalosaurus parkeri) beschrieben und 1964 durch Alick Walker einer eigenen Gattung Metriacanthosaurus zugeordnet.[3] 1988 stellte Gregory Scott Paul die Gattung in eine eigene Unterfamilie Metriacanthosaurinae Paul, 1988.[4]
Auf Basis der Funde von Sinraptor dongi stellten Philip J. Currie und Zhao Xijin 1993 diese Gattung gemeinsam mit Yangchuanosaurus in eine neue Familie, die sie als Sinraptoridae bezeichneten.[5] 1997 definierten Kevin Padian und John R. Hutchinson die Sinraptoridae als „Sinraptor und alle Allosauroidea, die näher damit verwandt sind als mit Allosaurus“.[6][1] In den folgenden Jahren wurde die Definition schrittweise immer weiter ausgebaut und gipfelte 2005 in einem Vorschlag Paul Serenos die Sinraptoridae zu definieren als „die umfassendste Klade, die Sinraptor dongi enthält aber nicht Allosaurus fragilis, Carcharodontosaurus saharicus oder Passer domesticus“. Der Haussperling (Passer domesticus) war in die Definition als abgrenzende Art mit aufgenommen worden um sicherzustellen, dass die Gruppe stabil bleibt auch für den Fall, dass sich herausstellen sollte, dass Sinraptor doch näher mit den Coelurosauria verwandt ist als mit den übrigen Allosauroidea.[7]
In einer umfassenden phylogenetischen Analyse der Tetanurae fanden Matthew T. Carrano, Roger B. J. Benson und Scott D. Sampson 2012 Metriacanthosaurus tief in der Klade der Sinraptoridae verwurzelt. Die Auroren erweiterten die Metriacanthosaurinae Paul, 1988 in den Rang einer Familie Metriacanthosauridae (Paul, 1988) und werteten die Sinraptoridae Currie & Zhao, 1993 als deren Juniorsynonym. Gleichzeitig wurde die Unterfamilie der Metriacanthosaurinae Paul, 1988 neu definiert als „alle Metriacanthosauridae, die näher verwandt sind mit Metriacanthosaurus als mit Yangchuanosaurus“.[3][4]
Christophe Hendrickx, Scott A. Hartman und Octávio Mateus passten 2015 Serenos Definition der Sinraptoridae an die neuen Gegebenheiten an. Gleichzeitig wurde die Definition der Unterfamilie Metriacanthosaurinae Paul, 1988 umformuliert zu: „Die umfassendste Klade, die zwar Metriacanthosaurus parkeri, nicht jedoch Yangchuanosaurus shangyouensis enthält“.[1][2]
Merkmale
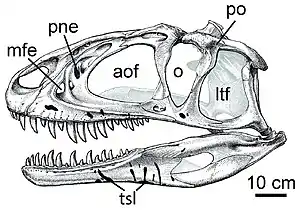
(In Klammer beigefügte Abkürzungen beziehen sich auf die Abbildung rechts)
Die Metriacanthosauridae zählen zu den ältesten Vertretern der Theropoda, die, bereits ab dem frühen Oberjura, eine Körpermasse von mehr als einer Tonne erreichen konnten.[3] Sie unterscheiden sich durch eine Reihe von Merkmalen von anderen Vertretern der Allosauroidea.
Ein anteriorer Ast der Maxilla ist nur schwach oder gar nicht ausgebildet. Das Laterosphenoid steht nicht nur mit dem Postorbitale („po“), sondern auch mit dem Frontale in Kontakt. Das Schuppenbein bewirkt keine Einschnürung im Bereich des Temporalfensters („ltf“), überdeckt mit einem flanschartigen Fortsatz jedoch den Kopf des Quadratbeines.[3]
Die laterale Seite des zahntragenden Unterkieferknochens (Dentale) zeigt eine in Längsrichtung verlaufende, deutlich ausgeprägte Rille mit einer Reihe von Foramina für Nerven und/oder Blutgefäße.[3]
Die ventrale Fläche des vorderen Wirbelkörpers (Interzentrum) des Axis-Wirbels ist anterodorsal geneigt und die spinopostzygapophyseale Lamina, eine Knochenlamelle zwischen Dornfortsatz und den Postzygapophysen, des Axis-Wirbels ist breit und deutlich ausgebildet. Die Dornfortsätze der mittleren Schwanzwirbel sind plattenförmig und im Umriss abgerundet rechteckig.[3]
Die Manus ist kürzer als Ober- und Unterarm zusammengenommen. Ein Os metacarpale quartum (MC IV) ist vorhanden, die entsprechenden Fingerknochen fehlen jedoch ebenso wie der ganze fünfte Finger.[3]
Die distalen Enden der beiden Sitzbeine sind bei adulten Individuen miteinander verschmolzen. Der Querschnitt der Sitzbeinschäfte ist mittig eher herzförmig als oval.[3]
Bezahnung
Alle Vertreter der Metriacanthosauridae weisen, soweit bekannt, zwei charakteristische Merkmale in Bezug auf die Bezahnung auf. Der Zahnschmelz zeigt eine unregelmäßige Oberflächentextur ohne Vorzugsorientierung und die Kronen der lateralen (weiter hinten im Kiefer stehenden[8]) Zähne zeigen auf der labialen Seite eine annähernd ebene, zentral gelegene Fläche.[9]
Metriacanthosaurinae
Die Unterfamilie der Metriacanthosaurinae zeichnet sich durch eine Reihe gemeinsamer Merkmale aus. Die Antorbitalgrube (Fossa antorbitalis), jene Schädelgrube vor den Augenhöhlen, in der sich auch das Antorbitalfenster (Fenestra antorbitalis, „aof“) befindet, wird anteroventral von einem erhöhten Knochenwall begrenzt. Die vorderen Rückenwirbel weisen ventral einen deutlichen Kiel auf. Der posteriore Rand des postacetabulären Fortsatzes des Darmbeins ist gerade und die „T-förmige“ Verlängerung („pubic boot“) des Schambeinschaftes schließt mit dessen Längsachse einen Winkel von <60° ein. Die Crista fibularis, ein Knochenkamm am Schienbein, ist bauchig vorgewölbt.[3]
Systematik
| Äußere Systematik | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||
| Systematische Stellung der Metriacanthosauridae innerhalb der Allosauroidea, vereinfacht nach Hendrickx et al., 2015.[1] |
Die Metriacanthosauridae sind neben den Allosauridae, den Neovenatoridae und den Carcharodontosauridae die vierte und gleichzeitig basalste Gruppe innerhalb der Allosauroidea.[1]
Intern lassen sich die Metriacanthosauridae zwei Hauptlinien zuordnen. Der Unterfamilie der Metriacanthosaurinae mit Metriacanthosaurus, Sinraptor, Siamotyrannus und möglicherweise auch Shidaisaurus steht die Gattung Yangchuanosaurus mit mindestens zwei Arten als Schwestertaxon gegenüber. Die Gattung Xuanhanosaurus zählt möglicherweise ebenfalls zu den Metriacanthosauridae, lässt sich innerhalb der Gruppe jedoch nicht genauer einordnen (Metriacanthosauridae incertae sedis).[3] Andere Autoren sehen in dieser Gattung jedoch eher einen basalen Vertreter der Allosauroidea abseits der Metriacanthosauridae[10] oder gar einen Vertreter der Megalosauroidea.[11]
Auch die systematische Stellung von Siamotyrannus ist keineswegs zweifelsfrei geklärt und einige Autoren halten es für möglich, dass es sich bei dieser Gattung um einen Vertreter der basalen Coelurosauria handelt und nicht um einen Vertreter der Metriacanthosauridae.[12] Die ebenfalls nur sehr bruchstückhaft erhaltene Gattung Erectopus kommt als weiterer möglicher Vertreter der Metriacanthosauridae in Frage.[3][9]
Zeitliche und geographische Verbreitung
Die ältesten Vertreter der Metriacanthosauridae (Shidaisaurus, Yangchuanosaurus zigongensis) lassen sich in Sedimenten des Mitteljuras von China nachweisen.[1][3] Die Typusgattung Metriacanthosaurus stammt aus dem frühen Oberjura (Oxfordium) von Dorset in England. Der gleichen Serie lassen sich auch Sinraptor und Yangchuanosaurus shangyouensis aus China zuordnen.[3]
Aus der Phu-Kradung-Formation Thailands, die sich in den obersten Jura bis in die unterste Kreide einstufen lässt, wurden zwei noch namenlose Vertreter der Metriacanthosauridae beschrieben und für denn Fall, dass sich die Zuordnung von Siamotyrannus zu den Metriacanthosauridae bestätigen sollte, würde sich die zeitliche Verbreitung der Familie bis in das Barremium der Unterkreide ausdehnen.[12] Die systematische Zuordnung der Gattung Erectopus aus Frankreich ist ebenfalls unsicher, könnte die zeitliche Verbreitung der Metriacanthosauridae aber sogar noch weiter bis in das Albium ausdehnen.[3][9]
Fossilfunde von Vertretern der Metriacanthosauridae beschränken sich dementsprechend auf Asien und Europa und die Familie war vermutlich in diesem Teil Laurasias endemisch.[12]
Weblinks
Einzelnachweise
- Ch. Hendrickx, S. A. Hartman & O. Mateus: An overview of non-avian theropod discoveries and classification. In: PalArch's Journal of Vertebrate Palaeontology, Band 12, Nummer 1, 2015, S. 1–73, (Digitalisat).
- Ch. Hendrickx & M. T. Carrano: Erratum on „An overview of non-avian theropod discoveries and classification“. In: PalArch's Journal of Vertebrate Palaeontology, Band 13, Nummer 2, 2016, S. 1–7, (Digitalisat).
- M. T. Carrano, R. B. J. Benson & S. D. Sampson: The phylogeny of Tetanurae (Dinosauria: Theropoda). In: Journal of Systematic Palaeontology, Band 10, Nummer 2, 2012, doi:10.1080/14772019.2011.630927, S. 211–300, (Digitalisat).
- M. T. Carrano, R. B. J. Benson & S. D. Sampson: Corrigendum: The phylogeny of Tetanurae (Dinosauria: Theropoda). In: Journal of Systematic Palaeontology, Band 10, Nummer 3, 2012, S. 599, (Digitalisat).
- P. J. Currie & X.-J. Zhao: A new carnosaur (Dinosauria, Theropoda) from the Jurassic of Xinjiang, People’s Republic of China. In: Canadian Journal of Earth Sciences, Band 30, Nummer 10, 1993, S. 2037–2081, (Abstract)
- K. Padian & J. R. Hutchinson: Allosauroidea. In: P. J. Currie & K. Padian (Hrsg.): Encyclopedia of Dinosaurs, Elsevier, 1997, ISBN 978-0-08-049474-6, S. 6f, (Leseprobe).
- P. C. Sereno: Sinraptoridae. In: TaxonSearch: Stem Archosauria 1.0. Paul Sereno, 7. November 2005, abgerufen am 15. Mai 2021.
- Ch. Hendrickx, O. Mateus & R. Araújo: A proposed terminology of theropod teeth (Dinosauria, Saurischia). In: Journal of Vertebrate Paleontology, Band 35, Nummer 5, 2015, Artikel e982797, doi:10.1080/02724634.2015.982797, S. 1–18, (Digitalisat)..
- Ch. Hendrickx, O. Mateus, R. Araújo & J. Choiniere: The distribution of dental features in non-avian theropod dinosaurs: Taxonomic potential, degree of homoplasy, and major evolutionary trends. In: Palaeontologia Electronica, 2019, Artikel 22.3.74, doi:10.26879/820, S. 1–110.
- O. W. M. Rauhut & D. Pol: Probable basal allosauroid from the early Middle Jurassic Cañadón Asfalto Formation of Argentina highlights phylogenetic uncertainty in tetanuran theropod dinosaurs. In: Scientific Reports, Band 9, 2019, Artikel 18826, doi:10.1038/s41598-019-53672-7.
- H. Dai, R. Benson, X. Hu, Q. Ma, C. Tan, N. Li, M. Xiao, H. Hu, Y. Zhou, Z. Wei, F.Zhang, S. Jiang, D. Li, G. Peng, Y. Yu & X. Xu: A new possible megalosauroid theropod from the Middle Jurassic Xintiangou Formation of Chongqing, People’s Republic of China and its implication for early tetanuran evolution. In: Scientific Reports, Band 10, 2020, Artikel 139, doi:10.1038/s41598-019-56959-x.
- A. Samathi, P. Chanthasit & P. M. Sander: A review of theropod dinosaurs from the Late Jurassic to mid-Cretaceous of Southeast Asia. In: Annales de Paléontologie, Band 105, Nummer 3, 2019, doi:10.1016/j.annpal.2019.03.003, S. 201–215, pdf.