Ossifikation
Ossifikation (von lateinisch Os „Knochen“ und facere „herstellen“) ist die Bildung von Knochengewebe im Wachstum, nach Brüchen oder bei pathologischer (krankhafter) Verknöcherung (heterotope Ossifikation). Osteogenese bezeichnet die Bildung eines individuellen Knochens. Während der Entwicklung können Knochen auf unterschiedlichen Wegen entstehen:
- aus Bindegewebe: desmale Osteogenese (Ossifikation),
- aus Knorpelgewebe: chondrale Osteogenese (Ossifikation),
- durch Anlagerung von Knochengewebe an bestehendes: appositionelle Ossifikation.
Desmale Ossifikation
Bei der desmalen Ossifikation entsteht das Knochengewebe direkt aus dem embryonalen Bindegewebe (Mesenchym), weshalb man auch von direkter Ossifikation spricht. So gebildete Knochen nennt man Bindegewebs-, Geflecht-, Deck- oder Belegknochen. Auf diese Art und Weise entstehen die Knochen des Schädeldachs und des Gesichtsschädels sowie das Schlüsselbein.
Mesenchymzellen verdichten sich inselartig zu Vorläuferzellen. Diese differenzieren sich zu Osteoblasten und bauen die Knochengrundsubstanz (Osteoid) auf, die anschließend mineralisiert. Dabei mauern sie sich durch die Anlagerung weiterer Osteoidschichten allmählich selbst ein und werden zu Osteozyten (ruhenden Zellen). Es entstehen so einzelne Verknöcherungspunkte, die sich zu Knochenspangen vereinigen und so den fertigen Knochen bilden.
Kommt es zu einem Knochenbruch, so wird immer zuerst einmal neues Knochengewebe gebildet, welches später zu Lamellenknochen umgewandelt wird (→ Knochenheilung).
Chondrale Ossifikation
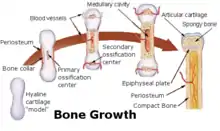
Bei dieser Form entstehen aus dem Mesenchym zunächst durch die Differenzierung von Mesenchymzellen zu Chondroblasten knorpelige Skelettelemente, das hyaline Primordialskelett, weshalb sie auch als indirekte Ossifikation bezeichnet wird. Man nennt diese Knochen auch Ersatzknochen.
Es gibt die Verknöcherung von innen her (enchondrale Ossifikation), dabei wachsen in das Knorpelgewebe Blutgefäße ein, in deren Begleitung sich Mesenchymzellen befinden. Diese differenzieren sich zu Chondroklasten (Knorpelabbau) und Osteoblasten (für den Knochenaufbau). Im Bereich der Epiphysenfugen kommt es durch den ständigen Auf- und Abbau zu einem Längenwachstum (interstitielles Wachstum).
Bei der Verknöcherung von außen (perichondrale Ossifikation) sondern sich von der Knorpelhaut (Perichondrium) Osteoblasten ab. Diese lagern sich ringförmig um das Knorpelmodell und es entsteht so eine Knochenmanschette. Die perichondrale Ossifikation findet am Mittelschaft (Diaphyse) der langen Röhrenknochen statt. Die perichondrale Ossifikation dient somit dem Dickenwachstum (appositionelles Wachstum). Auch entsteht im Inneren des Knochens ein Innenraum (primäres Mark), welches durch pluripotente Mesenchymzellen ersetzt wird und somit das eigentliche Knochenmark darstellt.
Bei beiden Formen der chondralen Ossifikation sondern die Osteoblasten eine Grundsubstanz, das Osteoid, ab. Durch den Einfluss der Osteoblastenfermente werden Kalksalze abgelagert. Die Osteoblasten differenzieren sich daraufhin zu Osteozyten. Die Punkte, von denen die Verknöcherung ausgeht, nennt man Knochenkerne oder Ossifikationszentren. Die Ossifikationszentren entstehen bei Nestflüchtern vor, bei den Nesthockern zumeist erst nach der Geburt.
Appositionelle Ossifikation
Bei der appositionellen Ossifikation wird Knochengewebe an bestehendes angelagert. Auf diese Weise erfolgt das Dickenwachstum von Knochen.
Knochenumbildung
Sowohl durch desmale als auch durch chondrale Ossifikation und auch bei der Heilung von Knochenbrüchen entsteht zunächst Geflecht- oder Faserknochen. Hier sind die Kollagenfibrillen der Knochengrundsubstanz noch ungeordnet. Die Faserknochen haben eine große Wachstumspotenz, jedoch eine geringe mechanische Festigkeit. Durch mechanische Beanspruchung (formative Reize) kommt es in den ersten Lebensjahren zum Umbau in den stabileren und strenger organisierten Lamellenknochen.
Der Aufbau des Lamellenknochens ist in der Substantia compacta am deutlichsten ausgeprägt. Er entsteht zunächst durch Osteoklasten, die größere, gefäßführende Kanäle in der Längsrichtung des (Geflecht-)Knochens freilegen, indem sie sich mit der Sekretion starker proteolytischer Enzyme einen Weg durch das Gewebe bahnen. Anschließend lagern sich Osteoblasten an die Wände dieses Kanals an, der als Havers-Kanal bezeichnet wird, und produzieren neue Grundsubstanz, bis sie sich „eingemauert“ haben (von da an bezeichnet man sie als Osteozyten). Durch die Anlagerung weiterer Schichten von Osteoblasten wird der Durchmesser des Kanales allmählich kleiner. Auf diese Weise entsteht ein sogenanntes Osteon – ein System aus etwa 5–20 mineralisierten Knochenlamellen, die konzentrisch um einen Havers-Kanal mit kleineren („Haverschen“) Blutgefäßen angeordnet sind. Zwischen den längs ausgerichteten Haverschen Gefäßen untereinander und den Gefäßen des Periosts gibt es Querverbindungen, die Volkmann-Kanäle.
Die Kollagenfaserbündel innerhalb der Lamellen verlaufen schraubenförmig um den Kanal, wobei sich der Drehsinn mit jeder angrenzenden Lamelle ändert. Sie sind hier durch Vorspannung gestreckt und nicht – wie sonst im Bindegewebe – gewellt. Dieser Aufbau aus gegenläufigen, vernetzten Spiralen wandelt Druck- und Zugbelastungen in Flächenpressungen um und verleiht dem Knochen seine besondere Stabilität.
Durch den ständigen Umbau, der im fertigen Knochen stattfindet, kommt es immer wieder zur Bildung von neuen Osteonen (siehe auch Knochengeweberemodellierung). Reste älterer Osteone, die man zwischen den jüngeren intakten Osteonen findet, bezeichnet man als Schaltlamellen.
Knochenwachstum
Kurze und platte Knochen wachsen durch äußere Anlagerung von Knochensubstanz. Auf diese Weise erfolgt auch das Dickenwachstum der langen Knochen.
Das Längenwachstum der langen Knochen erfolgt im Bereich einer Wachstumsplatte zwischen der endochondral ossifizierten Diaphyse (Mittelstück), um deren Rand eine perichondral entstandene Knochenmanschette liegt, und der enchondral entstandenen Epiphyse (Endstück). Man nennt diese Wachstumsplatte später, wenn sich die Diaphyse und die Epiphyse angenähert haben, auch Epiphysenfuge. Das Wachstum beruht darauf, dass sich die Chondrozyten mit gleicher Geschwindigkeit Richtung Epiphyse vermehren, wie sie von der Ossifikation aus der Diaphysenmitte „gejagt“ werden. Die Wachstumsplatte kann in vier Zonen unterteilt werden (aufgezählt entgegen der Wachstumsrichtung, also von Epiphyse zu Diaphyse):
- Reservezone: Hier liegt noch ein Vorrat von undifferenzierten Chondrozyten, aus dem Nachschub für die Proliferationszone bezogen werden kann.
- Proliferationszone: Hier liegen aktive Chondrozyten, die sich schnell mitotisch vermehren. Sie vermehren und wachsen dabei jedoch so, dass longitudinale Säulen gebildet werden. Diese Säulen sind durch die Longitudinalsepten getrennt, während die Chondrozyten in einer Säule durch die Transversalsepten getrennt werden.
- Hypertrophe Zone: Die in Säulen geordneten Chondrozyten wachsen hypertroph und mineralisieren dann die Longitundinalsepten, nicht aber die Transversalsepten.
- Öffnungszone: Die Chondrozyten sondern nun Enzyme ab, durch die die Transversalsepten abgebaut werden. Dann sterben sie ab (Apoptose). Dadurch entstehen Löcher, in die Makrophagen eindringen, um die Reste abzutragen. Diese Höhlen zwischen den Longitudinalsepten werden nun von Osteoblasten besetzt und ossifiziert.
Frakturen oder mechanische Traumata in diesem Bereich können dazu führen, dass die Fuge das Wachstum einstellt und der Knochen nicht mehr wächst. Mit dem Wachstumabschluss verschwindet diese Knorpelplatte vollständig und Dia- und Epiphysen wachsen knöchern zusammen. Die Grenze ist aber häufig noch als Fugenlinie (Linea physealis) zu erkennen.
Siehe auch
- Fibrodysplasia ossificans progressiva
- Osteogenesis imperfecta
- Achondroplasie
- Multiple epiphysäre Dysplasie
- Kraniosynostose (Verknöcherung der Schädelnähte)
- Syndesmophyt
- Osteophyt
Literatur
- Werner Linß, Jochen Fanghänel: Histologie: Zytologie, allgemeine Histologie, mikroskopische Anatomie. Walter de Gruyter, 1998, ISBN 3-11-014032-2, S. 65–68.
- Renate Lüllmann-Rauch: Taschenlehrbuch Histologie. Verlag Thieme, 2009, ISBN 978-3-13-129243-8, S. 162–165.