Doppelte Befruchtung
Eine doppelte Befruchtung findet bei der sexuellen Fortpflanzung der Bedecktsamer (Blütenpflanzen) statt. Dabei verschmelzen zwei Spermazellen mit zwei Zellen im Embryosack, woraus einerseits die Zygote und aus ihr der Embryo sowie andererseits das Endosperm hervorgeht. Dieser Vorgang ist eine Besonderheit der Blütenpflanzen, zu der es keine Parallele bei anderen Lebewesen gibt. Er wurde erstmals 1898 durch Sergei Gawrilowitsch Nawaschin beschrieben.[1]
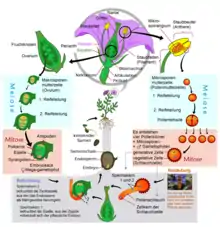
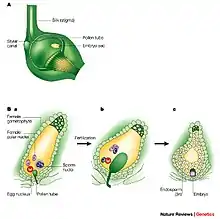
Ablauf
Im Pollenkorn werden zum Ende des Reifungsprozesses meistens zwei haploide Zellen angelegt: eine größere vegetative Zelle sowie eine kleinere generative Zelle. Die vegetative Zelle umgibt die generative Zelle.[2]
Im Rahmen der Bestäubung landet das Pollenkorn auf der Narbe eines Fruchtblatts oder des aus mehreren Fruchtblättern bestehenden Stempels in der Blüte. Das Pollenkorn keimt aus, indem die vegetative Zelle einen schlauchförmigen Auswuchs, den Pollenschlauch, hervorbringt. Dieser wächst durch den Griffel zum Fruchtknoten hinunter. Im Unterschied zu anderen Teilen der Pflanze ist das Wachstum des Pollenschlauchs nicht mit Zellteilungen verbunden, und es findet an der Spitze (apikal) statt, wie es auch für die Hyphen der Pilze charakteristisch ist. Die generative Zelle teilt sich vor oder meist kurz nach der Pollenkeimung in zwei Spermazellen, die männlichen Gameten, und diese wandern mit dem Zellkern der vegetativen Zelle den Pollenschlauch hinab.[3] Die kleinen Spermazellen bestehen nur aus einem Zellkern, der von sehr wenig Cytoplasma und einer Zellmembran umgeben ist.[4]
Die Spitze des Pollenschlauchs dringt in den Fruchtknoten ein und bis zum Eiapparat vor, der aus der Eizelle und zwei weiteren Zellen, den Synergiden, besteht.[5] Dies geschieht gewöhnlich über die Mikropyle, eine Öffnung in den Integumenten, die die Samenanlage umschließen. Der Pollenschlauch verschmilzt dann mit einer der beiden Synergiden und entleert sich in diese. Während der vegetative Pollenkern mit der betreffenden Synergide zugrunde geht, verschmilzt eine der beiden Spermazellen mit der benachbarten, ebenfalls haploiden Eizelle zur diploiden Zygote. Aus dieser entwickelt sich später der Embryo. Die zweite Spermazelle verschmilzt mit der angrenzenden diploiden oder zweikernigen Zentralzelle des Embryosacks zu der triploiden Endospermzelle. Aus dieser entwickelt sich im weiteren Verlauf das sekundäre Endosperm, ein nährstoffreiches Gewebe, welches im Normalfall den Embryo umgibt und ihn mit Nährstoffen versorgt.[6]
Die Zellverschmelzungen sind möglich, weil die Eizelle, die Synergiden und die Zentralzelle im Unterschied zu normalen Pflanzenzellen nur teilweise von dünnen Zellwänden umgeben sind.[7] Nach der Befruchtung erfolgen in der Endospermzelle schnell viele freie Kernteilungen ohne nachfolgende Zellteilungen, wodurch ein schnell wachsendes coenocytisches (vielkerniges) Endosperm entsteht. Die Entwicklung der Zygote zum Embryo beginnt erst später.[8]
Das evolutionär ursprünglichere haploide Endosperm der Nacktsamer, bei denen zumeist keine doppelte Befruchtung erfolgt, wird dagegen als primäres Endosperm bezeichnet. Einen Sonderfall innerhalb der Nacktsamer stellen die Gattungen Ephedra (Meerträubel) und Gnetum dar, bei denen aus einer doppelten Befruchtung zwei Zygoten hervorgehen, von denen sich aber zumeist nur eine zu einem Embryo entwickelt. Dagegen sind bei den Koniferen (Nadelbäume) und bei Welwitschia zwar zwei Spermazellen vorhanden, aber nur eine von ihnen befruchtet die Eizelle.[9]
Bei manchen Blütenpflanzen (Nachtkerzengewächse und Seerosenartige) enthält die Zentralzelle nur einen haploiden Kern, und das Endosperm ist daher nicht triploid, sondern diploid.[10]
Literatur
- Joachim W. Kadereit, Christian Körner, Benedikt Kost, Uwe Sonnewald: Strasburger – Lehrbuch der Pflanzenwissenschaften. 37. Auflage. Springer Spektrum, Berlin/ Heidelberg 2014, ISBN 978-3-642-54434-7, S. 169 f., 624.
Einzelnachweise
- Ilse Jahn (Hrsg.): Geschichte der Biologie. 3. Auflage, Sonderausgabe, Nikol, Hamburg 2004, ISBN 3-937872-01-9, S. 911.
- G. Czihak, H. Langer, H. Ziegler: Biologie. Springer, Berlin/ Heidelberg 1976, ISBN 3-642-96096-0, S. 224.
- B. M. Johri (Hrsg.): Embryology of Angiosperms. Springer, Berlin/ Heidelberg/ New York/ Tokyo 1984, ISBN 3-642-69302-4, S. 199 und 209.
- B. M. Johri (Hrsg.): Embryology of Angiosperms. Springer, Berlin u. a. 1984, S. 302.
- Doris Freudig: Lexikon der Biologie.: Eiapparat. Spektrum Akademischer Verlag, Heidelberg 1999, ISBN 3-8274-0326-X.
- Neil A. Campbell, Jane B. Reece: Biologie. 6. Auflage, Pearson Studium, München 2006, ISBN 3-8273-7180-5, S. 1507.
- B. M. Johri (Hrsg.): Embryology of Angiosperms. Springer, Berlin u. a. 1984, S. 174, 183, 186.
- B. M. Johri (Hrsg.): Embryology of Angiosperms. Springer, Berlin u. a. 1984, S. 306–308.
- Joachim W. Kadereit, Christian Körner, Benedikt Kost, Uwe Sonnewald: Strasburger – Lehrbuch der Pflanzenwissenschaften. 37. Auflage. Springer Spektrum, Berlin/ Heidelberg 2014, ISBN 978-3-642-54434-7, S. 169 f.
- Célia Baroux, Charles Spillane, Ueli Grossniklaus: Evolutionary origins of the endosperm in flowering plants. In: Genome Biology. Band 3, Nr. 9, 2002, Reviews 1026.1–5, PMC 139410 (freier Volltext).