Wonambi
Wonambi ist eine großwüchsige Schlangengattung des späten Känozoikums von Australien aus der nur fossil bekannten Familie Madtsoiidae.
| Wonambi | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktionen von Wonambi naracoortensis und Thylacoleo carnifex | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Unter-Miozän bis ? Holozän | ||||||||||||
| 23,03 bis 0,012 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Wonambi | ||||||||||||
| Smith, 1976 | ||||||||||||
Etymologie
Der Name „Wonambi“ stammt aus einer Sprache der Aborigines und bezeichnet die mythische Regenbogenschlange. Er wurde im Jahr 1976 von der australischen Paläontologin Meredith J. Smith geprägt.[1]
Merkmale
_fig_2.jpg.webp)
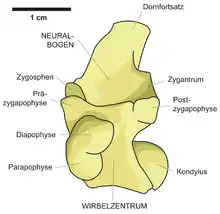
Die Gattung Wonambi bzw. ihre Typusart Wonambi naracoortensis (siehe unten) ist, wie für fossile Schlangenarten allgemein üblich, vor allem definiert über die speziellen Merkmale ihrer Rumpfwirbel. Dies sind ein hoher, nach hinten-aufwärts (posterodorsal) gerichteter Dornfortsatz mit scharfer Vorderkante („Präspinallamina“[2]) bis fast zum Rand des Zygosphens; ein relativ schmales Zygosphen mit steilen, 20 bis 30° gegen die Vertikale geneigten Gelenkflächen; große ParadiapophysenI, die seitlich bei den meisten Rumpfwirbeln über die Zygapophysen hinausragen, mit konkavem oberen (dorsalen) Rand der Diapophyse in Seitenansicht; relativ stark angestellte, 20° oder mehr gegen die Horizontale geneigte Zygapophysen sowie ein relativ schwach ausgeprägter bauchseitiger Kiel (Hämalkiel), der im hinteren Drittel des Wirbels oft charakteristisch als Zweifach- oder auch Dreifachkiel ausgebildet ist. Als weitere diagnostische Merkmale der Gattung werden genannt eine annähernd mittig auf dem Knochen verlaufende, deutlich vom inneren (medialen) Rand abgesetzte Pterygoidalzahnreihe, ein in Gaumenansicht dreieckiger äußerer (lateraler) Fortsatz des Pterygoids für die Artikulation mit dem Ectopterygoid (engl. ectopterygoid process), eine schmale, in gleichem Maße nach innen (mediad) wie nach oben (dorsad) weisende Basipterygoidgelenkgrube, ein relativ langgestrecktes und flaches Maxillare und Dentale sowie eine tiefe, anterolateral-posteromedial orientierte Hohlkehle (wahrscheinlich die Fassung für den ventralen Rand des Jugale) auf der dorsalen Seite des Maxillare.[3]
Wonambi war eine sehr großwüchsige Schlange. Berechnungen für ein in Naracoorte gefundenes, weitgehend vollständiges Skelett ergaben eine Lebendlänge von 5,4 bis 6,1 m, was bei einem maximalen Körperdurchmesser von 25 cm und einem maximalen Umfang von 80 cm ein Lebendgewicht von bis zu 250 kg ergeben hätte.[4] Auffällig ist der sehr ursprüngliche Bau des Schädels. Im Gegensatz zum Schädel der modernen Schlangen (Macrostomata) war der Oberkiefer (Prämaxillare und Maxillare) von Wonambi noch relativ fest mit dem übrigen Schädeldach verbunden. Auch besaß Wonambi noch ein Jugale („Jochbein“), ein üblicherweise am hinteren oberen Ende des Maxillare ansitzender Knochen des Schädeldachs, der bei modernen Schlangen komplett reduziert ist.[5] Des Weiteren zeigt das Flügelbein (Pterygoid) eine relativ kurze Gelenkgrube für das Basipterygoidgelenk, während moderne Schlangen langgestreckte Mulden haben, in denen das Pterygoid während des Schluckvorgangs am vorderen Hirnschädel vor- und zurückgleiten kann. All dies sind Hinweise darauf, dass der Schädel von Wonambi deutlich weniger in sich beweglich gewesen sein muss als der moderner Schlangen.[3][5]
Lebensweise
Wie alle nicht-colubroiden Schlangen besaß Wonambi keine Giftzähne. Deshalb und aufgrund ihrer Größe wird in Analogie zu den rezenten Boas und Pythons angenommen, dass Madtsoiiden und damit auch Wonambi terrestrische Würgeschlangen waren,[6] die gelegentlich auch größere Säugetiere töteten und verspeisten. Jedoch gibt es bislang keine weiteren konkreten Anhaltspunkte für die Zulässigkeit einer solchen Analogie.[7] Alternativ wird für Wonambi eine Lebensweise vorgeschlagen, die ähnlich ist wie die der rezenten australischen Arafura-Warzenschlange (Acrochordus arafurae). Diese tötet ihre Beute zwar durch Würgen, jedoch handelt es sich um eine aquatische Schlange, die sich fast ausschließlich von relativ kleinen Fischen ernährt. Für diese Hypothese spräche, dass Wonambi im Verhältnis zur Körpergröße einen relativ kleinen Schädel mit schwach gebauten Kiefern besaß, die mit großen Beutetieren nicht gut zurechtgekommen wären. Auch die weniger stark ausgeprägte Schädelkinetik spricht gegen größere Beutetiere, da die Kiefer wesentlich weniger weit hätten geöffnet werden können als bei modernen Schlangen. Wonambi hat möglicherweise in lagunenartigen Küstengewässern Fische gejagt. In den Höhlen, in denen die Überreste von W. naracoortensis gefunden wurden, könnte seinerzeit permanent Wasser gestanden haben, womit diese einen attraktiven Unterschlupf geboten hätten. Die zum Ende des Pleistozäns zunehmende Trockenheit im Süden Australiens könnte schließlich zum Aussterben der großen Schlangen geführt haben.[4]
Systematik und Fossilbericht
Äußere Systematik
Wonambi wird innerhalb der Schlangen zumeist eine sehr basale Stellung zugewiesen.[5][6][8] Sie ist wahrscheinlich mit einer Reihe weiterer „primitiver“ und ausgestorbener Schlangengattungen aus Südamerika, Afrika und Indien verwandt und wird mit ihnen zusammen in die Familie Madtsoiidae eingeordnet. Das bedeutet, trotz ihrer äußerlichen Ähnlichkeit mit heutigen Pythons und Boas ist sie mit diesen nicht näher verwandt (siehe → Konvergenz). Der Ursprung der Madtsoiiden liegt auf dem alten großen Südkontinent Gondwana. Die meisten Funde stammen aus der oberen Oberkreide und ab der Wende vom Eozän zum Oligozän ist die Gruppe außerhalb Australiens nicht mehr nachgewiesen.[9][10] Wonambi war demnach ein lebendes Fossil, ein Reliktvorkommen der Madtsoiiden, das sich im zunehmend geographisch isolierten Australien bis ins jüngste Känozoikum halten konnte. Eine alternative Hypothese sieht Wonambi jedoch als basale moderne Schlange innerhalb der Macrostomata.[11]
Arten
Es sind zwei Arten bekannt. Wonambi naracoortensis Smith 1976[1] ist die Typusart der Gattung und sowohl die geologisch jüngere als auch die häufigere. Sie ist aus mehreren Lokalitäten in South Australia und New South Wales bekannt, bei denen es sich zumeist um Karsthöhlen mit pleistozänen Ablagerungen („Höhlenerde“) handelt, so auch in der Typlokalität, den Höhlen von Naracoorte, die namensgebend für die Art sind. Das älteste Vorkommen von Wonambi naracoortensis datiert auf spätes Miozän oder frühes Pliozän, das jüngste auf spätes Pleistozän oder sogar frühes Holozän.[12] Damit dürfte Wonambi noch Zeitgenosse der ersten Menschen in Australien gewesen sein.[5]
Die zweite Art, Wonambi barriei Scanlon in Scanlon & Lee 2000[5], ist bislang nur aus dem Unter-Miozän von Riversleigh im Bundesstaat Queensland bekannt.[13] Sie ist nach dem australischen Privatsammler und Autodidakten John Barrie benannt, der wichtige Fundstücke der Gattung ausgrub, präparierte und teilweise auch wissenschaftlich bearbeitete.[5]
Wonambi naracoortensis unterscheidet sich von ihrer Schwesterart W. barriei vor allem durch ihre Größe, mit Rumpfwirbeln von oft mehr als 3 Zentimetern Breite und einer geschätzten Gesamtkörperlänge von mehr als 5 Metern. Die Rumpfwirbel ausgewachsener Exemplare von W. barriei sind hingegen weniger als 1,5 Zentimeter breit und ihre Gesamtkörperlänge betrug weniger als 3 Meter. Bei W. naracoortensis ist das Wirbelloch (die Durchtrittsstelle des Rückenmarks) im Verhältnis zum Wirbel klein, bei W. barriei ist es groß. Des Weiteren besitzt W. barriei ein relativ großes, auffälliges Foramen in der caudalen Oberfläche des Neuralbogens lateral des Zygantrums (engl.: parazygantral foramen). Bei W. naracoortensis treten oft mehrere, jedoch relativ unscheinbare und von den grübchenartigen Vertiefungen in besagter Region des Neuralbogens kaum zu unterscheidende Foramina auf. Die Zygapophysen sind bei W. naracoortensis mit ca. 25° steiler gegen die Horizontale aufgerichtet als bei W. barriei, bei der die Neigung nur 20° beträgt, und während das Palatinum (ein Gaumenknochen) bei W. naracoortensis relativ breit ist, ist es bei W. barriei eher schmal.[5]
Anmerkungen
- I Die Paradiapophyse ist ein seitlicher Fortsatz des Squamatenwirbels, gebildet aus Diapophyse (dorsal) und Parapophyse (ventral), der mit dem proximalen Ende der Rippe artikuliert.
Einzelnachweise
- Meredith J. Smith: Small fossil vertebrates from Victoria Cave, Naracoorte, South Australia. IV. Reptiles. Transactions of the Royal Society of South Australia. Bd. 100, Nr. 1, 1976, S. 39–51 (BHL).
- Emanuel Tschopp: Nomenclature of Vertebral Laminae in Lizards, with Comments on Ontogenetic and Serial Variation in Lacertini (Squamata, Lacertidae). In: PLoS ONE. Bd. 11, Nr. 2, 2016, e79420, doi:10.1371/journal.pone.0149445
- John D. Scanlon: Cranial morphology of the Plio–Pleistocene giant madtsoiid snake Wonambi naracoortensis. Acta Palaeontologica Polonica. Bd. 50, Nr. 1, 2005, S. 139–180 (online).
- D. John Barrie: Skull elements and additional remains of the Pleistocene boid snake Wonambi naracoortensis. Memoirs of the Queensland Museum. Bd. 28, Nr. 1, 1990, S. 139–151 (BHL).
- John D. Scanlon, Michael S. Y. Lee: The Pleistocene serpent Wonambi and the early evolution of snakes. Nature. Bd. 403, 2000, S. 416–420, doi:10.1038/35000188 (alternativer Volltextzugriff: ResearchGate).
- John D. Scanlon: Skull of the large non-macrostomatan snake Yurlunggur from the Australian Oligo-Miocene. Nature. Bd. 439, 2006, S. 840–842, doi:10.1038/nature04137 (alternativer Volltextzugriff: ResearchGate).
- Menna Jones, Chris R. Dickman, Michael Archer: Predators with Pouches: The Biology of Carnivorous Marsupials. CSIRO Publishing, Collingwood 2003, ISBN 0-643-06634-9, S. 119
- Allison Y. Hsiang, Daniel J. Field, Timothy H. Webster, Adam D.B. Behlke, Matthew B. Davis, Rachel A. Racicot, Jacques A. Gauthier: The origin of snakes: Revealing the ecology, behavior, and evolutionary history of early snakes using genomics, phenomics, and the fossil record. BMC Evolutionary Biology. Bd. 15, 2015, Art.-Nr. 87, doi:10.1186/s12862-015-0358-5.
- John D. Scanlon: The Basicranial Morphology of Madtsoiid Snakes (Squamata, Ophidia) and the Earliest Alethinophidia (Serpentes). Journal of Vertebrate Paleontology. Bd. 23, Nr. 4, 2003, S. 971–976, doi:10.1671/24 (alternativ: JSTOR 4524401).
- vgl. auch Madtsoiidae in der Paleobiology Database, abgerufen am 19. April 2016
- Olivier Rieppel, Arnold G. Kluge, Hussam Zaher: Testing the Phylogenetic Relationships of the Pleistocene Snake Wonambi naracoortensis Smith. Journal of Vertebrate Paleontology. Bd. 22, Nr. 4, 2002, S. 812–829, doi:10.1671/0272-4634(2002)022[0812:TTPROT]2.0.CO;2 (alternativ: JSTOR 4524280).
- Wonambi naracoortensis in der Paleobiology Database, abgerufen am 19. April 2016
- Wonambi barriei in der Paleobiology Database, abgerufen am 19. April 2016