Schopfhäher
Die Schopfhäher (Cyanocitta) sind eine Gattung der Rabenvögel (Corvidae). Sie umfassen mit dem Blauhäher (Cyanocitta cristata) und dem Diademhäher (Cyanocitta stelleri) zwei kleine Vertreter dieser Familie mit blauem Gefieder, schwarz gebändertem Großgefieder und charakteristischem Federschopf. Die Verbreitung der Gattung beschränkt sich auf das gemäßigte Nordamerika, wo die Rocky Mountains die beiden Arten voneinander trennen. Schopfhäher bewohnen Laub-, Misch- und Nadelwälder und ernähren sich überwiegend von Pflanzensamen – vor allem Eicheln – Wirbellosen und kleinen Wirbeltieren. Als Allesfresser nutzen sie aber auch viele andere Nahrungsquellen bis hin zu menschlichen Lebensmitteln. Beide Arten brüten von Frühjahr bis Frühsommer. Ihre Nester bauen die Schopfhäher in Baumkronen oder Sträuchern, ihre Gelege umfassen meist drei bis sechs Eier. Als einzige amerikanische Rabenvogelgattung verwenden sie beim Nestbau Schlamm. Trotz ihrer nahen Verwandtschaft und einiger Gemeinsamkeiten zeigen sie in ihrer Lebensweise teils aber auch starke Unterschiede, etwa im Hinblick auf Zugverhalten, Sozialität oder bei der Paarung.
| Schopfhäher | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Diademhäher (Cyanocitta stelleri) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Cyanocitta | ||||||||||||
| Strickland, 1845[1] |
Die Gattung Cyanocitta wurde 1845 von Hugh Edwin Strickland erstbeschrieben. Sie gehört zu den Neuwelthähern, einer Gruppe in Amerika endemischer Rabenvogel-Gattungen mit blauem Gefieder, und stellt innerhalb dieser Entwicklungslinie eine der am stärksten abgeleiteten Gattungen dar. Am nächsten mit den Schopfhähern verwandt sind die ebenfalls nordamerikanischen Buschhäher (Aphelocoma) und der Nacktschnabelhäher (Gymnorhinus). Während man lange Zeit annahm, dass sich die Vorfahren von Diadem- und Blauhäher während der letzten Eiszeiten trennten, deuten DNA-Untersuchungen auf eine Trennung vor rund fünf Millionen Jahren hin.
Merkmale
Körperbau und Farbgebung

Beide Schopfhäherarten sind relativ zierliche Rabenvögel mit sehr ähnlichem Körperbau, aber von unterschiedlicher Größe. Der Diademhäher wird mit 30–34 cm Körperlänge beziehungsweise 100–142 g Körpergewicht größer und schwerer als der 25–34 cm große und 70–100 g schwere Blauhäher. Bei den Maßen beider Arten gibt es nur geringe Überschneidungen. Der Schnabel der Schopfhäher entspricht mit seiner kräftigen, an der Spitze gekrümmten Gestalt dem Grundbauplan der Familie. Er ist schwarz gefärbt und weist an seinem Ende einen kleinen Würgerzahn auf. Die Nasalborsten sind sehr kurz und bedecken nur knapp die Nasenlöcher des Schnabels. Der leicht gerundete Schwanz ist im Verhältnis zum Rest des Körpers mittellang bis länglich, während die Flügel relativ kurz ausfallen. Im angelegten Zustand reichen ihre Spitzen nur knapp über den Schwanzansatz hinaus. Ein diagnostisches Merkmal der Schopfhäher ist ihre Federhaube. Sie ist beim Blauhäher weniger stark ausgebildet als beim Diademhäher. Männchen werden bei beiden Arten geringfügig größer als Weibchen. [2][3]
Die Gefiederfärbung der Schopfhäher entspricht mit ihren Blau-, Schwarz- und Weißtönen dem typischen Muster der Neuwelthäher. Die beiden Arten zeigen sehr unterschiedliche Zeichnungen: Während der Diademhäher eine variable Färbung mit schwarzem Schwanz, blauem Rumpfgefieder und schwarzem bis blauem Kopf aufweist, zeigt der Blauhäher ein recht einheitliches Muster mit blauer Körperoberseite, weißer Unterseite und schwarzer Gesichtsmaske. Allerdings besitzen beide Arten schwarz gebänderte Schwanzfedern, Armschwingen, Schirmfedern und Flügeldecken; ein Merkmal das allen anderen Neuwelthähern fehlt und unter allen amerikanischen Rabenvogelarten diagnostisch ist. Schnabel und Beine sind bei beiden Schopfhähern schwarzgrau. Zwischen Männchen und Weibchen gibt es in der Färbung keinen Unterschied, auch Jungtiere unterscheiden sich nur geringfügig von adulten. [4]
Flugbild und Fortbewegung
Während der Diademhäher mit kräftigen, bedächtigen Flügelschlägen fliegt, ist der Streckenflug des Blauhähers von flachen Flügelschlägen geprägt. Kürzere Distanzen überwindet der letztere mit einem stufenweise abfallenden Flug, bei dem sich Phasen schneller Flügelschläge mit Gleitphasen abwechseln. Auf dem Boden tendieren beide Arten aufgrund ihrer geringen Größe und ihrer recht kurzen Beine eher zum Hüpfen als zum Schreiten. Im Geäst bewegen sich Schopfhäher sehr agil von Ast zu Ast. [2][3]
Lautäußerungen
Schopfhäher besitzen ein sehr breites Repertoire an Rufen. Darunter fallen sowohl krächzende, energische Lautäußerungen als auch gutturale oder melodiöse Rufe wie Subsongs. Das Vokabular beider Arten ist sehr unterschiedlich. Sie zeigen nur punktuelle Gemeinsamkeiten, etwa im Geschäcker weiblicher Vögel. Blauhäher wie Diademhäher gelten als gute Imitatoren und geben häufig Rufe von sich, die denen anderer Vögel und von Säugetieren stark ähneln. Beide Arten sind bekannt dafür, die Rufe von Bussarden (Buteo spp.) und anderen Greifvögeln täuschend echt nachzuahmen. Auch die Laute von Säugetieren oder mechanische Umgebungsgeräusche werden von Blau- und Diademhähern imitiert. Wie stark dieses Verhalten in der Natur aber auf bewusster, aktiver Nachahmung basiert und welchem Zweck es dient, ist umstritten. [2][3]
Verbreitung und Wanderungen
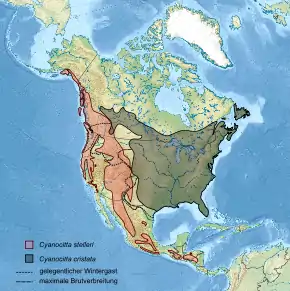
Die Schopfhäher sind im gesamten gemäßigten Nordamerika verbreitet und werden durch die nordamerikanischen Kordilleren voneinander getrennt. Das heutige Verbreitungsmuster ist wahrscheinlich ein Ergebnis der Vergletscherung großer Teile Nordamerikas während der letzten Kaltzeiten. Während der Blauhäher im eher flachen Ostteil des Kontinents bis zu den Rocky Mountains vorkommt, lebt der Diademhäher parapatrisch zu ihm in den westlichen Gebirgen und den angrenzenden gemäßigten und bergigeren Regionen. Lediglich im südwestlichen Kanada kommen Blau- und Diademhäher regional sympatrisch vor. Beide Arten stoßen bis an die Nordgrenze der warmgemäßigten Zone in Kanada und Alaska vor. Sie fehlen jeweils in den ariden Wüsten, Tälern und Küstenregionen des westlichen Kontinents. Nach Süden hin wird das Verbreitungsgebiet fragmentierter. Nur der Diademhäher stößt in die Tropen vor, bewohnt dort aber auch nur die kühleren Gebirgsregionen. Seine südliche Verbreitungsgrenze stellt das Tiefland von Nicaragua dar. [2][3] Im Pleistozän wurde er wahrscheinlich in verschiedenen, eisfreien Wärmeinseln im Süden und Westen isoliert und drang nach dem Abschmelzen der Gletscher bis in die Rocky Mountains vor.[5]
Deutliche Unterschiede zwischen den beiden Arten der Gattung zeigen sich im Zugverhalten. Während der an kältere Höhenlagen angepasste Diademhäher als Standvogel gilt, und im Winter nur die höchsten Lagen verlässt,[3] ist der Blauhäher die einzige Art unter den Neuwelthähern, die über das Jahr hinweg auch Breitenwanderungen unternimmt. Zwar verbleiben stets einige Blauhäher im Winter in den sommerlichen Brutgebieten, aber ein großer Teil der Population zieht zumindest in den nördlicheren Teilen des Verbreitungsgebiets nach Süden. In südlicheren Breiten – etwa in Florida – besteht die gesamte Brutpopulation aus Standvögeln. [2] Während der Vogelzug beim Blauhäher zeitlich und geographisch weitgehend konstant ist, kann es beim Diademhäher nach Ende der Brutsaison zu irruptiven Wanderungsbewegungen kommen, wenn aufgrund des Populationsanstiegs Nahrungsknappheit herrscht. Viele einjährige Vögel verlassen dann die Brutgebiete und wandern auch in außerhalb gelegene Regionen ab. [3]
Lebensraum
Schopfhäher sind Waldbewohner, meiden aber wie die meisten Rabenvögel dichte und geschlossene Wälder. Stattdessen sind sie vor allem in lückigen Vegetationsformen, entlang von Waldrändern, in Parkland oder urbanen Grünanlagen anzutreffen. Beide Arten zeigen eine große Affinität zu Eichen (Quercus spp.), wo vorhanden,[6] sind aber nicht zwingend auf sie angewiesen. Typische Habitate der Gattung sind Nadel- und Mischwälder, der Blauhäher bewohnt auch reine Laubwälder. Beide Arten sind auch in anthropogenen Habitaten anzutreffen, nutzen sie aber unterschiedlich. Während der Blauhäher häufig in Städten anzutreffen ist und dort höhere Bestandsdichten als in Wäldern aufweist,[2] verhält es sich beim Diademhäher umgekehrt. Seine Siedlungsdichte fällt in auch nur geringfügig mit Dörfern oder Städten durchsetzten Landstrichen deutlich gegenüber etwa Nationalparks ab.[7] Während sich die Vorkommen des Diademhähers auf Höhenlagen von 1000 bis 3500 m konzentrieren,[3] zeigt der Blauhäher keine derartige Bindung und kommt kontinuierlich von Meeresstränden bis in die Appalachen vor.[8]
Lebensweise
Ernährung
Schopfhäher sind Allesfresser, ernähren sich aber vorwiegend von pflanzlicher Nahrung. Für beide Arten ergaben Studien eine Verteilung von 75–80 % Pflanzenmaterial und 25–20 % tierischem Material in der Nahrung. Eicheln sind für die Schopfhäher in weiten Teilen ihres Verbreitungsgebiets eine wichtige Nahrung und machen, wo vorhanden, oft den Großteil der pflanzlichen Nahrung aus. Sie können aber auch durch Haselnüsse, Kiefernsamen, Bucheckern oder vergleichbare Nussfrüchte ersetzt werden. Der Blauhäher tendiert dabei eher zu Samen von Laubbäumen, während der Diademhäher aufgrund seines montanen Habitats eher Koniferensamen nutzt. Wo sich andere Nahrungsquellen bieten, werden diese gerne ausgenutzt. Gliederfüßer, kleine Singvögel und Säugetiere sowie Eier werden nicht verschmäht. [2][3]
Wie alle Neuwelthäher besitzen die Schopfhäher ein modifiziertes Kiefergelenk. Sie gehören zu den Gattungen, bei denen dieses Merkmal besonders stark ausgeprägt ist. Es erlaubt ihnen, Nahrungsstücke mit geöffnetem Schnabel aufzuhämmern und deren Schale dabei gleich mit zu entfernen. [9] Laborstudien mit Blauhähern deuten darauf hin, dass Schopfhäher bei der Nahrungssuche mit Suchbildern arbeiten, bei denen bestimmte Strukturen, die auf Nahrungsobjekte hindeuten, deutlicher hervorstechen.[2] Der für Corviden typische Verstecktrieb ist bei den Schopfhähern stark ausgeprägt. Überschüssige Nahrung wird in Spalten und Löchern von Bäumen platziert[3] oder zunächst mit dem Schnabel in den Boden gehämmert und das Loch anschließend bedeckt. [10]
Sozial- und Territorialverhalten
Das Territorialitätsverhalten von Schopfhähern ist im Familienvergleich schwach bis moderat ausgeprägt. Der Diademhäher verteidigt einen kleinen Bereich rund um das Nest, der Blauhäher nur den Nistplatz selbst. Die Dominanz am eigenen Nistplatz wird vor allem durch Imponierhaltungen und -figuren zur Schau gestellt. Bei beiden Arten ist das Brutpaar die übliche soziale Einheit, in der sich Vögel bewegen. Sowohl Blau- als auch Diademhäher können sich in größeren Schwärmen zusammenfinden. Während bei Diademhäherschwärme ein gewisser Zusammenhalt zu erkennen ist, formen sich größere Blauhähergruppen eher spontan und zufällig und besitzen keine Eigendynamik. Die Individualdistanz zwischen einigen Vögeln beträgt in der Regel wenige Meter, kann zwischen Individuen mit niedrigem Status (etwa diesjährige Vögel) oder Brutpartnern auch auf wenige Zentimeter reduziert werden. [2][3]
Fortpflanzung und Brut

Wie bei Rabenvögeln üblich bilden auch Schopfhäher meist lebenslange, monogame Brutpaare. Trennungen kommen nur in seltenen Fällen vor. Das Balzverhalten ist bei beiden Arten verschieden; während beim Schopfhäher meist das Männchen die Paarung einleitet, geschieht das beim Blauhäher durch das Weibchen. Schopfhäher bauen ihr Nest in Baumkronen oder dem Geäst von Sträuchern. Die Höhe des Nestes variiert und kann 1–30 m betragen. Der Diademhäher baut meist in 3–5 m Höhe, beim Blauhäher zeigt sich keine klare Präferenz. Beide Geschlechter beteiligen sich am Nestbau, die eigentliche Konstruktion wird aber vorrangig vom Weibchen übernommen. Das Nest besteht außenseitig aus einer Schale aus dicken, grob geschichteten Zweigen, die mit weicheren Materialien – etwa Blätter, Haare, Federn oder auch Papier – verwoben werden. Innenseitig tragen Schopfhäher Schlamm auf, der dann mit weichen, feinen Wurzeln ausgekleidet wird. Sie sind die einzige Gattung der Neuwelthäher, die Schlamm beim Nestbau verwendet. [2][3]
Die Eier sind auf hellem Grund dunkel gesprenkelt und werden wahrscheinlich im Abstand von je einem Tag gelegt. Bis zum Schlüpfen der Jungen vergehen rund zweieinhalb Wochen, bis zum Flüggewerden der Jungen vergeht etwa die gleiche Zeit. Das Weibchen bebrütet die Eier allein und wird während dieser Zeit vom Männchen gefüttert. Auch nach dem Ausflug aus dem Nest werden die Jungen noch von den Eltern gefüttert. [2][3]
Systematik
Taxonomie und Forschungsgeschichte
Die Gattung der Schopfhäher wurde 1845 von Hugh Edwin Strickland erstbeschrieben. Er trennte sie von den Blauraben (Cyanocorax) ab, um ihrer unterschiedlichen Morphologie und Lebensweise Rechnung zu tragen. Die Schopfhäher, so Strickland, unterschieden sich im gleichen Maße von den Blauraben wie die Gattung Garrulus von den Raben und Krähen (Corvus). Als Typart wählte er den Blauhäher (Cyanocitta cristata). Der Gattungsname Cyanocitta setzt sich aus den griechischen Begriffen κύανος kyanos für blau und κίττα kitta, einem Namen für die Elster ab. Strickland stellte auch jene Arten in die Gattung, die heute den Buschhähern (Aphelocoma) zugerechnet werden. [11] Zeitweise wurden neben den Buschhähern auch die Dohlenhäher (Cyanolyca) und die „Cissilopha“-Blauraben als Schopfhäher betrachtet,[12] spätestens mit John William Hardys Revision von 1969 umfasste die Gattung aber nur noch den Blau- und den Diademhäher.[13] Der Blauhäher war den europäischen Naturforschern bereits im 16. Jahrhundert bekannt und wurde 1758 von Carl von Linné erstbeschrieben.[14] Der Diademhäher wurde 1741 während der Zweiten Kamtschatkaexpedition von Georg Wilhelm Steller entdeckt und 1789 von Johann Friedrich Gmelin beschrieben.[15]
Äußere Systematik
| |||||||||||||||||||||||||||
| Innere Systematik der rezenten Neuwelthäher nach Bonaccorso & Peterson 2007. Die nächstverwandte Gattung der Schopfhäher sind die nordamerikanischen Buschhäher. |
Die Beziehungen der Schopfhäher zu anderen Neuwelthähergattungen galten lange als unklar. Hardy vermochte anhand von Ökologie und Morphologie keine nähere Verwandtschaft zu einer der anderen Gruppen zu erkennen,[16] Derek Goodwin vermutete 1986 eine nähere Verwandtschaft zu den Buschhähern.[17] Tatsächlich ordneten Analysen verschiedener Gensequenzen durch Elisa Bonaccorso und A. Townsend Peterson die Busch- und die Schopfhäher 2007 als Schwestertaxa ein. Gemeinsam stehen sie dem Nacktschnabelhäher (Gymnorhinus cyanocephalus) als Schwesterklade gegenüber. Der Bootstrap-Support für diese Anordnung ist zwar gering; auch eine andere Anordnung der drei Gattungen untereinander wäre auf Basis der analysierten Sequenzen denkbar. Busch- und Schopfhäher zeigen aber im Unterschied zu allen anderen Neuwelthähern einen seitlichen Randknochen am Skleralring, was für eine enge Verwandtschaft spricht. Die von Nacktschnabel-, Busch- und Schopfhähern gebildete Klade entwickelte sich vermutlich gegen das späte Miozän oder frühe Pliozän[3] im südlichen Nordamerika und besiedelte in der Folgezeit den Norden des Kontinents. [18] Trotz der relativ großen Distanz zu anderen Gattungen hybridisiert etwa der Blauhäher mit einigen Buschhäher- und Blaurabenarten.[19]
Innere Systematik
Weil Diadem- und Blauhäher jeweils den Westen beziehungsweise den Osten Nordamerikas bewohnen, wurde lange Zeit vermutet, dass die Kaltzeiten des Pleistozäns und die Vergletscherung der Rocky Mountains das ausschlaggebende Ereignis für die Artbildung waren. Da beide Arten miteinander hybridisieren, galt eine späte Trennung vor etwa 100.000 Jahren als wahrscheinlich. Relativ große Unterschiede in den Cytochrom-b-Sequenzen von 10,7–24,8[20] % deuten aber auf eine frühere Trennung hin. Im Schema der Molekularen Uhr müssten sich die Vorfahren der beiden Arten damit vor rund 5 Millionen Jahren, also zu Beginn des Pliozäns getrennt haben. [3]
Quellen
Literatur
- Brian S. Arbogast, Joseph B. Slowinski: Pleistocene Speciation and the Mitochondrial DNA Clock. In: Science 282, 1998. S. 1995a.
- Elisa Bonaccorso, Andrew Townsend Peterson: A Multilocus Phylogeny of New World Jay Genera. In: Molecular Phylogenetics and Evolution 42, 2007. doi:10.1016/j.ympev.2006.06.025, S. 467–476.
- Theresa M. Burg, Anthony J. Gaston, Kevin Winker, Vicky L. Friesen: Rapid Divergence and Postglacial Colonization in Western North American Steller’s Jays (Cyanocitta stelleri). In: Molecular Ecology 14, 2005. doi:10.1111/j.1365-294x.2005.02710.x, S. 3745–3755.
- Johann Friedrich Gmelin: Caroli a Linné systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis … Editio decima tertia, aucta, reformata. Regnum Animale. Georg Emanuel Beer, Leipzig 1788. doi:10.5962/bhl.title.545
- Derek Goodwin: Crows of the World. 2. Auflage. The British Museum (Natural History), London 1986. ISBN 0565009796.
- John William Hardy: A Taxonomic Revision of the New World Jays. In: The Condor 71, 1969. S. 360–375. (Volltext; PDF; 1,5 MB)
- Joseph del Hoyo, Andrew Elliot, David Christie (Hrsg.): Handbook of the Birds of the World. Volume 14: Bush-shrikes To Old World Sparrows. Lynx Edicions, Barcelona 2009. ISBN 9788496553507.
- Carl von Linné: Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Lars Salvi, Stockholm 1758. (Volltext)
- Steve Madge, Hilary Burn: Crows & Jays. Princeton University Press, Princeton 1994, ISBN 0-691-08883-7.
- Hugh Edwin Strickland: On Cyanocitta, a Proposed New Genus of Garrulinae, and on C. superciliosa, a New Species of Blue Jay, hitherho Confounded with C. ultramarina, Bonap.. In: The Annals and Magazine of Natural History, Including Zoology, Botany and Geology 15 (98), 1845. S. 260–261. (Volltext)
- Richard L. Zusi: A Feeding Adaption of the Jaw Articulation in the New World Jays (Corvidae). In: The Auk 104, 1987. S. 665–680.
Weblinks
- Erick Greene, William Davison, Vincent R. Muehter: Steller’s Jay (Cyanocitta stelleri). In: A. Poole: The Birds of North America Online (343), Cornell Lab of Ornithology, Ithaca 1998. doi:10.2173/bna.343.
- Keith A. Tarvin, Glen E. Woolfenden: Blue Jay (Cyanocitta cristata). In: A. Poole: The Birds of North America Online (469), Cornell Lab of Ornithology, Ithaca 1999. doi:10.2173/bna.469.
Einzelnachweise
- Strickland 1845, S. 261.
- Tarvin & Woolfenden 1999. Abgerufen am 3. April 1999.
- Greene et al. 1998. Abgerufen am 3. April 2012.
- Madge & Burn 1994, S. 67–70.
- Burg et al. 2005, S. 3752.
- Goodwin 1986, S. 227.
- del Hoyo 2009, S. 569.
- Goodwin 1986, S. 228.
- Zusi 1987, S. 669–671.
- Goodwin 1986, S. 227–233.
- Strickland 1845, S. 260–261.
- Amadon 1944, S. 6.
- Hardy 1969, S. 370–371.
- von Linné 1758, S. 106.
- Gmelin 1788, S. 370.
- Hardy 1969, S. 370.
- Goodwin 1986, S. 222.
- Bonaccorso & Peterson 2007, S. 472–474.
- del Hoyo 2009, S. 568.
- Arbogast & Slowinski 1998, S. 1995a.
Fußnoten direkt hinter einer Aussage belegen diese einzelne Aussage, Fußnoten direkt hinter einem Satzzeichen den gesamten vorangehenden Satz. Fußnoten hinter einer Leerstelle beziehen sich auf den kompletten vorangegangenen Text.