Ophthalmosaurus
Ophthalmosaurus ist eine Gattung der Ichthyosaurier (Ichthyopterygia), die die Fossilien mittelgroßer Thunnosaurier (Thunnosauria) versammelt. Ihre Vertreter lebten vom mittleren Jura bis zur anbrechenden Kreide (rund 166 bis 145 Millionen Jahre vor heute) am Nordrand des Tethysozeans und der Westküste Laurasias. Ophthalmosaurus war angesichts seiner Körperform und seiner großen, durch Knochenringe verstärkten Augen wohl ein schneller Jäger, der in großen Tiefen Jagd auf Fische, Ammoniten und Belemniten machte und seinen Nachwuchs lebend gebar. Die Gattung stellt einen typischen Vertreter jurassischer Ichthyosaurier dar, die sich allesamt durch einen thunfischähnlichen Bauplan auszeichneten und wohl eine breite Palette ökologischer Nischen besetzten. In ihren derzeitigen taxonomischen Grenzen ist die Gattung umstritten, die europäische Art Ophthalmosaurus icenius und der nordamerikanische Ophthalmosaurus natans bilden wohl keine monophyletische Gruppe.
| Ophthalmosaurus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
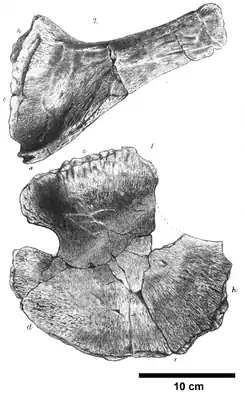
Fossiles Rabenbein (oben) und Schulterblatt (unten) aus dem Holotyp von Ophthalmosaurus icenius | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Mitteljura bis Unterkreide (Callovium bis Berriasium) | ||||||||||||
| 166,1 bis 145 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Ophthalmosaurus | ||||||||||||
| Seeley, 1874[1] | ||||||||||||
Die Erstbeschreibung von Ophthalmosaurus durch Harry Govier Seeley stammt von 1874 und basiert auf Material aus dem englischen Oxford Clay, von wo die meisten bekannten Funde stammen. Weiteres Material stammt aus Frankreich und den USA. Ophthalmosaurus gilt als häufige Art in seinen Fundschichten, wird aber gerade in Europa oft nur fragmentarisch gefunden. Während die Gattung in den vergangenen Jahrzehnten geographisch und taxonomisch weit gefasst wurde, zeichnet sich in jüngerer Zeit eine Abtrennung der Art O. natans in die Gattung Baptanodon ab, womit O. icenius als einzige Art in einer monotypischen Gattung verbleiben würde.
Merkmale

Ophthalmosaurus wurde bis zu 4 m lang, erreichte ein geschätztes Gewicht von 930 kg[2] und zeichnete sich wie alle anderen jurassischen Ichthyosaurier durch eine thunfischförmige Gestalt mit langer, schnabelähnlicher Schnauze aus: Der Rücken wölbte sich vom Kopf an relativ steil nach oben hin auf, erreichte knapp hinter den Vorderflossen seinen Scheitelpunkt und fiel dann leicht zum Schwanz hin ab. Die Bauchlinie zeigte einen ähnlichen Verlauf, während die Rücken- und die Schwanzflosse, verglichen mit anderen Gattungen der Ichthyosaurier, sehr lang, hoch und schmal waren. Während viele Ichthyosaurier eine seitlich abgeflachte Körperform besaßen, deuten Ophthalmosaurus-Fossilien auf einen runderen Körper hin. Dieser war durch seine äußere Form nicht nur äußerst stromlinienförmig, er war auch durch seinen Knochenbau stark versteift und stabilisiert: Die Rückenwirbel zeigten die für spätere Ichthyosaurier typische Eishockeypuck-Form (sehr flach und breit), saßen dicht aneinander und sorgten damit für eine sehr dicke und starre Wirbelsäule. Den Tieren waren somit keine Schlängelbewegungen – weder seitwärts noch auf und ab – mehr möglich. Umgekehrt konnten sie so aber durch den Schub der Flossen sehr hohe Geschwindigkeiten erreichen, weil der Körper kompakt und gut gegen Turbulenzen und Verformungen stabilisiert war. Seinen Antrieb gewann Ophthalmosaurus wahrscheinlich vor allem durch das seitliche Schlagen seiner mondsichelförmigen Schwanzflosse, deren unterer Teil durch die Schwanzwirbelsäule und der Rest wahrscheinlich durch Knorpelgewebe versteift war. Die ebenfalls aus Knorpel aufgebaute Rückenflosse trug wohl zur Lagestabilisierung bei.[3]

Ophthalmosaurus besaß mit 50 cm relativ lange und nach vorne hin schmal zulaufende Kiefer. Die Zähne saßen in schmalen Rillen im Ober- und Unterkiefer, waren also nicht vollständig thekodont und insgesamt nur schwach verankert. Ein diagnostisches Merkmal der Gattung ist die Form der Maxilla, die einen Teil der Nasenlöcher bildet. Die Augen waren wie bei allen Ichthyosauriern durch einen verstärkten Skleralring aus 15 Knochenplatten stabilisiert und in Relation zur Körpergröße sehr groß. Der Skleralring schützte das Auge vor hohem Wasserdruck und damit vor Verformungen, was insbesondere bei der Druckdifferenz wichtig war, die sich bei hohen Geschwindigkeiten zwischen den vorderen und hinteren Partien der Augen ergaben.[2] Die Schulterblätter waren zum Körper hin verbreitert, die Vorderflossen besaßen einen einzelnen zusätzlichen Finger und waren wie die kleineren Hinterflossen aus gerundeten Knochengliedern aufgebaut; alle drei Merkmale sind diagnostisch für die Gattung. Die Hüfte bestand aus zwei getrennten Teilen und war, wie für spätjurassische Ichthyosaurier üblich, nur noch mit den Hinterextremitäten verbunden.[4]
Ökologie
Die großen, lichtstarken Augen und der stromlinienförmige Körper von Ophthalmosaurus zeichnen das Bild eines schnellen Schwimmers, der in große Tiefen vorstieß, um dort Jagd auf seine Beute zu machen. Ophthalmosaurus war wahrscheinlich in der Lage, auch noch in einer Tiefe von über 500 m Beute aufzuspüren und zu fangen.[2] Damit spiegelt Ophthalmosaurus eine generelle Entwicklung der Ichthyosaurier im Jura wider, bei der sich thunfischartige, schnelle Formen durchsetzen konnten[5] und auf der ganzen Welt Verbreitung fanden. Diese Lebensweise bezieht sich aber vor allem auf die Typusart O. icenius. Für andere Arten, deren Zugehörigkeit zu Ophthalmosaurus vorwiegend auf der allgemeinen Körperform basiert und in der Regel umstritten ist, liegen kaum spezifische Erkenntnisse zum Lebensraum vor. In anderen Fällen deutet die Skelettstruktur im Vergleich mit O. icenius auf eine deutlich abweichende Lebensweise hin, etwa bei O. (Baptanodon) natans, der weniger schnell, dafür wendiger und eher ein Flachwasserbewohner gewesen sein dürfte.[6]
Die Nahrung von Ophthalmosaurus bestand unter anderem aus Belemniten und Ammoniten, wie sich anhand von Fanghaken und Schalen dieser Tiere in fossilen Skeletten nachweisen lässt. Hinzu dürften Fische gekommen sein. Die Beute wurde wahrscheinlich mit hoher Geschwindigkeit aus Fisch- und Kopffüßerschwärmen heraus mit den Kiefern gegriffen und anschließend geschluckt. Wie für Ichthyosaurier typisch war Ophthalmosaurus vivipar, seine Jungen wurden also lebend geboren und waren bei der Geburt bereits voll schwimmfähig.[7]
Fossilmaterial und Stratigraphie

Die ersten Ophthalmosaurus-Exemplare wurden im englischen Oxford Clay gefunden. Diese Gesteinsformation stammt aus dem mittleren beziehungsweise späten Jura und reicht vom Callovium bis ins Kimmeridgium (166,1 bis 152,1 mya). Die dort geborgenen Fossilien werden sämtlich der Art O. icenius zugeordnet, für die sich außerhalb von England auch im nordwestlichen Frankreich Nachweise finden. Insgesamt sind die westeuropäischen Funde von Ophthalmosaurus zahlreich, aber häufig nur bruchstückhaft erhalten. Die vollständigeren nordamerikanischen Fossilien, die als O. baptanodon zusammengefasst werden, stammen aus der Sundance-Formation der nordwestlichen USA, die auf das Oxfordium (rund 160 mya) datiert wird. Hinzu kommen verschiedene Funde aus der Oberjura des Wolgabeckens in Russland, deren Gattungszugehörigkeit aber umstritten ist.[8] Unklar ist auch der Status von englischem Ichthyosaurier-Material, das aus der frühkreidezeitlichen (Berriasium, 145–139,3 mya) Spilsby-Sandstone-Formation von Nettleton stammt. Es lässt sich nicht O. icenius zuordnen, gehört aber wahrscheinlich zur Gattung Ophthalmosaurus, womit diese über die Jura-Kreide-Grenze hinaus existiert hätte.[9]
Taxonomie und Systematik
| |||||||||||||||||||||||||||||||||
| Systematische Stellung von Ophthalmosaurus nach Fischer et al. (2013). Die Gattung ist im Bezug auf Acamptonectes paraphyletisch, eine entsprechende Revision steht aber bislang aus.[10] |
Das Typusexemplar der Art Ophthalmosaurus icenius und damit der Gattung Ophthalmosaurus stammt aus dem Peterborough Member des britischen Oxford Clays. Es umfasst verschiedene Teile eines disartikulierten Skelettes, darunter Stücke des Unterkiefers und des Schädels, Teile der Vorderflossen, mehrere Wirbelkörper- und Bögen sowie einige Rippen. Es wurde im 19. Jahrhundert von Charles Leeds freigelegt und präpariert. Dieses Skelett wird heute unter der Inventarnummer NHMUK R2133 im Londoner Natural History Museum verwahrt. Harry Govier Seeley beschrieb Ophthalmosaurus 1874 als zweite Ichthyosauriergattung überhaupt nach Ichthyosaurus. Der Gattungsname leitet sich vom altgriechischen „ophthalmos“ für „Auge“ (in Anlehnung an die großen Augenhöhlen der Tiere) und „sauros“ für „Echse“ her. Das Artepitheton erinnert an die Icenier, einen keltischen Volksstamm, der die Gegend um den Erstfundort bewohnte.[1]
Über die Jahrzehnte wurden Ophthalmosaurus neben der Typusart O. icenius eine Reihe weiterer Fossilien beziehungsweise fossil überlieferter Arten zugerechnet, deren Fundorte sich über Nordamerika und Eurasien verteilen: Die 1880 von Othniel Marsh für Fossilien aus dem Oxfordium Wyomings aufgestellte Art Baptanodon („Sauranodon“) natans wurde 1910 von Charles William Andrews zu Ophthalmosaurus gestellt, ebenso wie alle anderen in dieser Gattung beschriebenen nordamerikanischen Fossilien. Auch die argentinischen Ichthyosaurier des Bajociums (Ophthalmosaurus periallus) sowie die von Maxim Arkhangelsky und Vladimir Efimov aufgestellten Arten aus dem Jura Russlands (O. yasikovi, O. gorodischensis, O. saveljeviensis) wurden in den Ichthyopterygia-Monografien von Michael Maisch und Andreas Matzke[4] beziehungsweise von Christopher McGowan und Ryosuke Motani[11] Ophthalmosaurus zugeschlagen. Die Grundlage hierfür bildeten zunächst vor allem Kopf- und Körperformen von Fossilien und Skelettrekonstruktionen. Dreidimensionale Schädelrekonstruktionen von Robert Appleby, die dieser als ein Argument zur Trennung von Baptanodon und Ophthalmosaurus anführte, wurden von späteren Autoren aufgrund methodischer Zweifel zurückgewiesen. Die Unterschiede zwischen beiden Formen galten als zu klein, um die Gattungen zu trennen.[12] Phylogenetische Analysen auf Basis von osteologischen Merkmalen gruppierten die unter Ophthalmosaurus zusammengefassten Formen bis Anfang des 21. Jahrhunderts stets zusammen, was gegen ein Splitten der Gattung sprach. Ophthalmosaurus war entsprechend eine kosmopolitische Gattung, für deren Paläoökologie O. icenius als beispielhaft galt.[13]
Erst mit einer Reihe ophthalmosaurider Fossilfunde aus verschiedenen jurassischen Fossillagerstätten Europas und vergleichenden Analysen der Skelettmorphologie ergab sich eine Tendenz zur Aufspaltung von Ophthalmosaurus in verschiedene Gattungen: Mittlerweile werden die argentinischen Funde der Gattung Mollesaurus zugeschlagen, während eine Reihe von Studien konsistente Unterschiede zwischen O.-icenius-Material und nordamerikanischen Fossilien (Baptanodon) herausarbeiten konnten.[14] Mit dem Fund von Fossilien in Großbritannien und Deutschland, die deutlich von Ophthalmosaurus abwichen, in phylogenetischen Analysen aber als Schwestertaxon von O. natans aufschienen, ergaben sich 2012 endgültig Zweifel an der Zusammenfassung in einer gemeinsamen Gattung. Während die Neufunde als Acamptonectes eine eigene Gattung erhielten, wurde O. natans zunächst in der nun paraphyletischen Gattung Ophthalmosaurus belassen. Der taxonomische Status der russischen und amerikanischen Fossilien ist seitdem ungeklärt, was vor allem auf das Fehlen detaillierter osteologischer Vergleiche zurückzuführen ist.[15]
Entwicklungsgeschichtlich ist Ophthalmosaurus ein typischer Vertreter einer mitteljurassischen Radiation der Thunnosaurier (Thunnosauria). Diese Gruppe erschien am Beginn des Juras und zeichnete sich vor allem durch ihre thunfischartige Körperform aus. Im Mittleren Jura entstanden mehrere Formen, die diesen Bauplan weiter optimierten und als Ophthalmosauridae zusammengefasst werden. Innerhalb der Ophthalmosauridae gehört Ophthalmosaurus gemeinsam mit Mollesaurus und Acamptonectes (und wohl auch Baptanodon) zur Untergruppe der Ophthalmosaurinae, die sich gegenüber ihrer Schwestergruppe Platypterygiinae durch einen im Schnitt größeren Körper und weniger spezialisierte Kiefer- und Schädelformen auszeichnete. Während man aufgrund des Fossilberichts lange davon ausging, dass die Ophthalmosaurinae zum Ende des Juras ausstarben, dokumentiert die 2012 beschriebene Gattung Acamptonectes, die derzeit noch innerhalb von Ophthalmosaurus steht, den Fortbestand ophthalmosauriner Ichthyosaurier bis in die Unterkreide.[16]
Quellen
Literatur
- Richard Ellis: Sea Dragons. Predators of the Prehistoric Oceans. University Press of Kansas, Lawrence, Kansas 2003, ISBN 0-7006-1269-6.
- Valentin Fischer, Michael W. Maisch, Darren Naish, Ralf Kosma, Jeff Liston, Ulrich Joger, Fritz J. Krüger, Judith Pardo Pérez, Jessica Tainsh, Robert M. Appleby: New Ophthalmosaurid Ichthyosaurs from the European Lower Cretaceous Demonstrate Extensive Ichthyosaur Survival across the Jurassic–Cretaceous Boundary. In: PLoS ONE. Band 7, Nr. 1, 2012, S. e29234, doi:10.1371/journal.pone.0029234.
- Valentin Fischer, Robert M. Appleby, Darren Naish, Jeff Liston, James B. Riding, Stephen Brindley, Pascal Godefroit: A basal thunnosaurian from Iraq reveals disparate phylogenetic origins for Cretaceous ichthyosaurs. In: Biology Letters. Band 9, Nr. 4, 2013, S. 1–6, doi:10.1098/rsbl.2013.0021.
- Valentin Fischer, Maxim S. Arkhangelsky, Gleb N. Uspensky, Ilya M. Stenshin, Pascal Godefroit: A new Lower Cretaceous ichthyosaur from Russia reveals skull shape conservatism within Ophthalmosaurinae. In: Geological Magazine. 2013, S. 1–11, doi:10.1017/S0016756812000994.
- Judy A. Massare, Emily A. Buchholtz, Jennifer M. Kenney, Anne-Marie Chomat: Vertebral morphology of Ophthalmosaurus natans (Reptilia: Ichthyosauria) from the Jurassic Sundance Formation of Wyoming. In: Paludicola. Band 5 (4), 2006, S. 242–254.
- Christopher McGowan, Ryosuke Motani: Ichthyopterygia. In: Handbook of Paleoherpetology, Part 8. Friedrich Pfeil, München 2003, ISBN 3-89937-007-4.
- Michael Maisch, Andreas Matzke: The Ichthyosauria. In: Stuttgarter Beiträge zur Naturkunde. Serie B (Geologie und Paläontologie). Band 298, 2000, S. 1–155.
- Ryosuke Motani, Bruce M. Rothschild, William Wahl, Jr: Large eyeballs in diving ichthyosaurs. In: Nature. Band 402, 1999, S. 747.
- Harry Govier Seeley: On the Pectoral Arch and Fore Limb of Ophthalmosaurus, a New Ichthyosaurian Genus from the Oxford Clay. In: Quarterly Journal of the Geological Society. Band 30, Nr. 1–4, 1874, S. 696–707, doi:10.1144/GSL.JGS.1874.030.01-04.64.
- Motani R.: Scaling effects in caudal fin propulsion and the speed of ichthyosaurs. In: Nature. Band 415, Nr. 6869, 17. Januar 2002, S. 309–312, PMID 11797005.
Weblinks
Einzelnachweise
- Seeley 1874, S. 699.
- Motani et al. 1999, S. 747.
- McGowan & Motani 2003, S. 47.
- Maisch & Matzke 2000, S. 78.
- Motani et al. 2002, S. 309.
- Massare et al. 2006, S. 251.
- Ellis 2003, S. 92–99.
- McGowan & Motani 2003, S. 114–115.
- Fischer et al. 2012, S. 17.
- Fischer et al. 2013a, S. 4.
- McGowan & Motani 2003, S. 110.
- Massare et al. 2006, S. 242.
- McGowan & Motani 2003, S. 110–113.
- Massare et al. 2006, S. 245–247.
- Fischer et al. 2012, S. 19–20.
- Fischer et al. 2013b, S. 10.