Ganzhorns Mausmaki
Ganzhorns Mausmaki (Microcebus ganzhorni) ist eine Primatenart aus der Gattung der Mausmakis innerhalb der Teilordnung der Lemuren. Er ist im Südosten Madagaskars endemisch und wurde mit zwei weiteren Arten der Mausmakis 2016 erstbeschrieben.
| Ganzhorns Mausmaki | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Microcebus ganzhorni | ||||||||||||
| Hotaling, Foley, Lawrence, Bocanegra, Blanco, Rasoloarison, Kappeler, Barrett, Yoder, Weisrock, 2016 |
Merkmale
Da der Holotyp eine Gewebeprobe ist enthält die Erstbeschreibung keine Angaben zu den Merkmalen der Art. Der Zoo Prag bezeichnet seine zuvor als Graue Mausmakis identifizierten Tiere als Ganzhorns Mausmakis, allerdings ohne anzugeben wie die Identifizierung erfolgte. Den Angaben des Zoos zufolge haben sie ein feines gräuliches Fell, bisweilen mit einem dunklen Längsstreifen, und einen buschigen Schwanz. Ganzhorns Mausmaki gehört zu den größeren Mausmakiarten und erreicht eine Körperlänge von 12,5 bis 15 Zentimetern und eine Schwanzlänge von etwa 13,5 Zentimeter bei einem Gewicht von 39 bis 90 Gramm.[1] Für eine vergleichende Studie vermeintlicher Populationen des Grauen Mausmakis wurden zwischen 2000 und 2003 127 Exemplare von Ganzhorns Mausmaki im Mandenga Forest vermessen und einige Maße dokumentiert. Die Schwanzlänge betrug durchschnittlich 13,25 Zentimeter, die Länge der Hinterfüße 3,24 Zentimeter, Länge und Breite des Schädels 3,5 und 2,18 Zentimeter, und Länge und Breite der Ohren 2,35 und 1,76 Zentimeter. Das Gewicht der männlichen Exemplare schwankte zwischen durchschnittlich 56,6 Gramm im Juni und 76,9 Gramm im Dezember. Bei den etwas größeren weiblichen Exemplaren schwankte das Gewicht zwischen durchschnittlich 64,3 Gramm im Juni und 105,6 Gramm im November. Dabei wurde nicht zwischen trächtigen und anderen Weibchen unterschieden.[2]
Lebensweise
Ganzhorns Mausmaki ist wie alle Mausmakis nachtaktiv und hält sich meist auf den Bäumen auf. Er ist ein Allesfresser, der sich vor allem von Früchten und Insekten ernährt. Der Zoo Prag nennt eine Ernährung von Wirbellosen, Früchten, Blüten und Nektar. Eine Freilandstudie über vier Jahre erbrachte das Ergebnis, dass Ganzhorns Mausmaki sich vorwiegend von Früchten ernährt, wobei Früchte von mehr als 35 Millimeter Länge ganz gemieden und deutlich kleinere Beeren bevorzugt werden. Durch direkte Beobachtung, Samen im Kot und Fraßspuren an Früchten konnte der Verzehr der Früchte von 66 verschiedenen Pflanzenarten nachgewiesen werden. Die Samen der Früchte von 34 verschiedenen Arten passierten den Verdauungstrakt unbeschadet. Ganzhorns Mausmakis spielen in ihrem Habitat eine bedeutende Rolle bei der Verbreitung von Pflanzensamen bis zu einer Länge von 6,5 Millimeter. Während 63 Prozent aller beobachteten Nahrungsaufnahmen Früchte betrafen, wurden bei 22 Prozent Blüten, 11 Prozent Gliederfüßer und 4 Prozent Gummen gefressen. Die Untersuchung erbrachte für die beiden sympatrischen Arten der Fettschwanzmakis sehr ähnliche Ergebnisse, die Nahrungskonkurrenz wird nicht durch die Wahl unterschiedlichen Futters, sondern durch den Aufenthalt in unterschiedlichen Höhen der Baumkronen vermindert: durchschnittlich 4 Meter bei Ganzhorns Mausmaki, 5 Meter beim Westlichen Fettschwanzmaki und 7 Meter beim Braunen Fettschwanzmaki.[3]
Ganzhorns Mausmakis verbringen die Nächte alleine mit der Nahrungssuche und die Tage alleine oder in Gruppen in Schlafnestern. Dabei gibt es im Unterschied zu Lemuren wie den Fettschwanzmakis keine feste Paarbindung. Während die weiblichen Tiere auf einer Fläche von etwa einem halben Hektar verbleiben, streifen die männlichen auf bis zu drei Hektar umher.[4] Die Trächtigkeitsdauer wird vom Zoo Prag mit 54 bis 68 Tagen angegeben, ein Wurf umfasst zwei oder drei Junge.[1] Ganzhorns Mausmakis haben innerhalb eines Jahres zwei oder drei Würfe. Obgleich es zwischen Oktober und April oder Mai zu Würfen kommt, ist der November der Monat mit mehr als der Hälfte der Würfe und der Januar/Februar der Zeitraum mit mehr als einem Viertel aller Würfe. Es wird vermutet, dass es im April/Mai zu einem dritten Wurf kommt.[2]
Ganzhorns Mausmakis haben zwei alternativ zu nutzende Strategien, um auf Zeiten mangelnden Nahrungsangebots und ungünstiger Umweltbedingungen mit einem reduzierten Energieverbrauch zu reagieren. Ein Winterschlaf eines Tieres dauerte 30 Tage. Dabei folgte die Körpertemperatur der Umgebungstemperatur und sank bis auf 11,5 °C ab. In unregelmäßigen Abständen wachte das Tier auf, dabei stieg die Körpertemperatur auf das normale Niveau an. Diese Wachphasen umfassten insgesamt 17,1 Prozent der Dauer des Winterschlafs. Ein zweites untersuchtes Tier fiel nur wiederholt während der Morgenstunden in Phasen des Torpors, wobei der Torpor durchschnittlich etwa eine Stunde und maximal fünf Stunden anhielt, und an 24 von 85 Untersuchungstagen auftrat. Die Körpertemperatur sank dabei nicht unter 27 °C und während der Nächte wurde normale Aktivität gezeigt.[5] Da Weibchen häufiger Winterschlaf halten kann zu Zeiten, in denen sich große Teile der Population im Winterschlaf befinden, der Anteil männlicher Tiere unter den umherstreifenden Mausmakis erhöht sein.[6] Insbesondere im Juli und August, dem Südwinter und der Zeit, in der es an Früchten und Blüten mangelt, befinden sich viele Tiere im Winterschlaf.[3]
Die Lebensdauer beträgt in der Gefangenschaft bis zu 14 Jahre.[1] In der Freiheit wird diese Lebensdauer bei weitem nicht erreicht. Im Rahmen einer Langzeitstudie wurden zwischen den Jahren 2000 und 2003 171 Exemplare gefangen. Nur 13 Prozent der Tiere, 16 Prozent der Weibchen und 9 Prozent der Männchen, wurden im folgenden Jahr erneut gefangen, und nur zwei Weibchen und ein Männchen im dritten Jahr.[2]
Parasiten
Die Parasitenfauna der frei lebenden Population von Ganzhorns Mausmaki ist eingehend untersucht worden. Im Zeitraum von März 2003 bis April 2004 wurden 101 Exemplare in Fallen gefangen und ihre Fäkalien auf Endoparasiten untersucht. Dabei wurden Spulwürmer (Prävalenz 26,7 %), Strongyliden (15,9 %), Peitschenwürmer (8,9 %), Haarwürmer (1 %), Trichostrongyliden (2 %), Pfriemenschwänze (16,8 %), nicht näher bestimmte Nematoden (59,4 %), Bandwürmer (38,7 %), Saugwürmer (2 %), Kratzwürmer (2 %) und Kokzidien (68,3 %) nachgewiesen. Den mindestens 20 nachgewiesenen Arten gastrointestinaler Parasiten bei Ganzhorns Mausmaki stehen ganze vier Arten von Nematoden gegenüber, die der französische Parasitologe Alain Chabaud mit seinen Mitarbeitern 1965 für den Grauen Mausmaki nennen konnte.[7] Die Prävalenz des Parasitenbefalls ist bei Ganzhorns Mausmaki deutlich höher als bei den ebenfalls im Mandena Forest vorkommenden Arten Brauner Fettschwanzmaki und Westlicher Fettschwanzmaki. Das wird auf die deutlich höhere Populationsdichte des Mausmakis zurückgeführt. Als Folge ihrer Promiskuität und der größeren Mobilität weisen männliche Mausmakis eine größere Diversität ihrer Parasitenfauna auf. Die Übertragung einwirtiger Parasiten wird durch das Sozialverhalten der Wirte mit engem Kontakt in Familienverbänden begünstigt, während mehrwirtige Parasiten mit Entwicklungsstadien in wirbellosen Wirten davon profitieren, dass Mausmakis auch Wirbellose verzehren. Da einige Parasiten sowohl die an den Mandena Forest gebundenen Mausmakis als auch die innerhalb und außerhalb der Fragmente des Waldes lebenden Hausratten befallen gelten die Ratten als wichtige Vektoren, die Parasiten zwischen weitgehend voneinander isolierten Populationen von Mausmakis transportieren. Da der Lebensraum von Ganzhorns Mausmaki häufig von Menschen aufgesucht wird besteht die Sorge der Übertragung von zoonotischen Parasiten auf Besucher oder menschlicher Krankheitserreger auf die Mausmakis.[4][8]
Verbreitung und Lebensraum
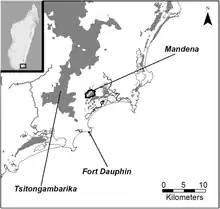
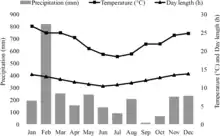
Der Typenfundort ist der Mandena Forest im Bezirk Taolanaro der Region Anosy, etwa 12 Kilometer nördlich der Stadt Tolagnaro im Südosten Madagaskars (24° 56′ 54,2″ S, 46° 59′ 42″ O). Der Mandena Forest ist ein etwa 230 Hektar großes Schutzgebiet, das eine unmittelbar benachbarte madagassische Bergbaugesellschaft der Rio Tinto Group 1999 zum Ausgleich von Eingriffen in die Natur durch den Abbau von Ilmenit einrichten musste. Über die Ausdehnung des tatsächlichen Verbreitungsgebiets und die Größe der Population ist nichts bekannt, es besteht jedoch keine Verbindung mit den Verbreitungsgebieten des Grauen Mausmakis im Westen Madagaskars.[9] Der Madena Forest liegt etwa drei Kilometer von der Küste entfernt auf 0 bis 20 Meter Höhe über dem Meeresspiegel. Er besteht aus immergrünen Bäumen von zehn bis fünfzehn Meter Höhe mit dichtem Unterholz, die in einem sandigen Substrat wurzeln. Im Vergleich zu den Regenwäldern im Osten Madagaskars hat die Küstenzone einen weniger ausgeprägten saisonalen Wechsel der Niederschlagsmengen. Die Durchschnittstemperatur liegt bei 22,5 °C und der durchschnittliche jährliche Niederschlag beträgt etwa 1600 mm, mit einer Regenzeit von November bis April. Etwa 82 Hektar des Schutzgebiets werden periodisch überflutet, die übrigen 148 Hektar bestehen aus fragmentierten Resten des Küstenwalds.[8][10]
Das bekannte Verbreitungsgebiet von Ganzhorns Mausmaki gehört zu einer für die Küstenzone im Südosten Madagaskars charakteristischen Landschaft, die bis in die 1950er Jahre durchgehend bewaldet war, und seither von starken anthropopogenen Veränderungen betroffen ist. Zur Gewinnung von Bauholz und zur Produktion von Holzkohle für die nahe gelegene Regionalhauptstadt Tolagnaro, zugunsten der menschlichen Besiedlung und für den Bergbau sind große Teile der Wälder abgeholzt worden. Zwischen 1995 und 1998 verstärkte sich das Ausmaß der Umweltzerstörung rapide und heute sind von den natürlichen Waldflächen weniger als zehn Prozent erhalten. Diese Restflächen sind stark fragmentiert und es besteht die Sorge, dass durch die Habitattrennung vereinzelte Populationen ohne genetischen Austausch entstehen. Die unmittelbare Umgebung des fragmentierten Restwaldes wird von dem Betriebsgelände der Bergbaugesellschaft, Monokulturen von Eucalyptus robusta und Zitroneneukalyptus zur Produktion von Bauholz und der Bodenerosion preisgegebenem sandigem Brachland mit Bewuchs aus Heidekräutern und Ruderalvegetation eingenommen.[10][11][6]
Jüngere populationsgenetische Untersuchungen, bei denen die variablen MHC-Komplexe im Fokus der Betrachtung standen, erbrachten die Erkenntnis, dass die Population von Ganzhorns Mausmaki im Mandena Forest vergleichsweise klein ist, diese Art aber im Vergleich zu anderen Lemuren weniger von einer Verringerung der genetischen Vielfalt betroffen ist. Bei der Untersuchung der Populationen einzelner Fragmente konnte zunächst ein Zusammenhang von Größe der Populationen und Größe und Qualität der Fragmente hergestellt werden. Größere Fragmente wiesen größere Populationen auf und starken anthropogenen Veränderungen unterworfene Fragmente hatten geringere Populationsdichten. Die Fragmentierung der Habitate hatte den Austausch zwischen den Populationen nicht völlig unterbunden, die eingerichteten Korridore haben den genetischen Austausch allerdings deutlich gesteigert.[10][11][12]
Die Lemurenfauna Madagaskars wird seit Jahrzehnten intensiv erforscht. Auch die Mausmakis des Mandena Forest waren Objekte von Untersuchungen, einschließlich mehrerer parasitologischer und populationsbiologischer Langzeitstudien.[13] In den auf diesen Studien basierenden Veröffentlichungen werden Ganzhorns Mausmakis durchweg als Graue Mausmakis bezeichnet. Da der Graue Mausmaki im Mandena Forest nicht vorkommt können diese früheren Veröffentlichungen sicher Ganzhorns Mausmaki zugeordnet werden.[14] Neben Ganzhorns Mausmaki lebt im Madena Forest eine Reihe weiterer Lemuren: die kathemeralen Arten Halsbandmaki und Südlicher Bambuslemur sowie die ebenfalls nachtaktiven Arten Brauner Fettschwanzmaki, Westlicher Fettschwanzmaki und Südlicher Wollmaki.[10] Bei den beiden Fettschwanzmakis ist unklar ob sie den Sumpfbereich des Mandena Forest besiedeln, alle anderen Arten sind auch dort vertreten.[15]
Systematik
Ganzhorns Mausmaki ist eine Art der Gattung Mausmakis in der Familie Katzenmakis. Bis zum späten 20. Jahrhundert waren nur wenige Mausmakis beschrieben worden. Seither wurden zahlreiche Arten abgetrennt und teilweise nur nach Fotos oder DNA-Proben beschrieben. Ganzhorns Mausmaki wurde im Jahr 2016 zusammen mit Microcebus boraha und Microcebus manitatra beschrieben. Damit enthält die Gattung Microcebus 24 Arten.[9]
Erstbeschreibung
Bereits 2009 stellte eine Forschergruppe um David W. Weisrock von der Duke University im Rahmen einer umfassenden Studie zur Abgrenzung der Arten madagassischer Mausmakis fest, dass die Mausmakis des Mandena Forest eine Klade bilden. Ein Artstatus wurde ihnen jedoch noch nicht zugestanden.[16] Erst 2016 erfolgte die Erstbeschreibung von Ganzhorns Mausmaki und zwei weiteren bereits 2009 identifizierten Kladen durch ein Team um den US-amerikanischen Zoologen Scott Hotaling, dem auch Weisrock und die meisten Autoren von 2009 angehörten.[9]
Der Holotyp ist eine Gewebeprobe, die am 8. August 1998 durch Jörg Ganzhorn im Mandena Forest vom Ohr eines ausgewachsenen männlichen Tieres entnommen wurde, das zunächst als Grauer Mausmaki galt. Die Probe wurde unter der Nummer DLC #100 in die Sammlung des Duke Lemur Center der Duke University aufgenommen. Die Erstbeschreibung enthält keine Maße oder sonstige Angaben zum Phänotyp. Durch eine Analyse der nukleären und mitochondrialen DNA konnte bestätigt werden, dass die Mausmakis des Mandena Forest eine Klade bilden. Mit Hilfe statistischer Analysen wurde die Klade sicher als Art identifiziert.[9] Die hier vorgenommene Festlegung einer DNA-Probe als Holotyp – an Stelle eines konservierten Exemplars – ist ungewöhnlich, wurde aber in den vergangenen Jahren bei neu beschriebenen Primaten wiederholt praktiziert, wenn es sich bei den Typexemplaren um Zootiere handelte oder der Artenschutz ihre Freilassung erforderte. Die Internationalen Regeln für die Zoologische Nomenklatur erlauben ein solches Vorgehen, das allerdings in der Vergangenheit umstritten war.[17]
Der Artname ganzhorni ehrt den deutschen Biologen und Hochschullehrer Jörg Ganzhorn vom Zoologischen Museum der Universität Hamburg, der seit mehr als 30 Jahren zur Ökologie und Biodiversität Madagaskars forscht und die der Erstbeschreibung zugrunde liegende Gewebeprobe genommen hat. Die Benennung erfolgte in Würdigung seiner Beiträge zur Kenntnis der Mausmakis und anderer Lemuren und seiner Bemühungen um ihren Schutz.[9][18]
Seit der Erstbeschreibung von Ganzhorns Mausmaki meldeten der Zoo Pilsen und der Zoo Prag, diese Art zu halten. Zwei Paare in Pilsen waren zuvor als Graue Mausmakis identifiziert worden.[19][1]
Gefährdung und Schutz
Für Ganzhorns Mausmaki ist noch keine Gefährdungsanalyse durch die IUCN vorgenommen worden. Alle Katzenmakis unterliegen als Arten des Anhang I den Handelsbeschränkungen des Washingtoner Artenschutzübereinkommens.[20] Da die Habitatzerstörung die größte Bedrohung für die Lemuren Madagaskars darstellt ist die Habitaterhaltung die wichtigste Schutzmaßnahme.
Das Schutzgebiet Mandena Forest dient dem Schutz aller dort lebenden Lemuren. Zu den Umweltschutz-Auflagen, denen die in unmittelbarer Nähe tätige Bergbaugesellschaft unterworfen wird, gehört die Einrichtung von Korridoren durch die Anpflanzung von Bäumen, mit denen die Fragmente des Reliktwaldes vernetzt werden. Diese Korridore bestehen zu 20 Prozent aus autochthonen Arten und zu 80 Prozent aus Exoten. Sie werden von Ganzhorns Mausmakis innerhalb von fünf Jahren nach der Pflanzung angenommen, sofern sie mit geeigneten Bäumen bepflanzt sind. Mischungen einheimischer Bäume mit dem australischen Eucalyptus robusta sowie Korridore mit der ebenfalls exotischen Acacia mangium werden gut angenommen, solche mit der in Madagaskar invasiven Art Melaleuca quinquenervia hingegen nicht. Korridore mit dichtem Unterholz werden bevorzugt genutzt.[10][11][21]
Literatur
- Scott Hotaling, Mary E. Foley, Nicolette M. Lawrence, Jose Bocanegra, Marina B. Blanco, Rodin Rasoloarison, Peter M. Kappeler, Meredith A. Barrett, Anne D. Yoder, David W. Weisrock. Species discovery and validation in a cryptic radiation of endangered primates: coalescent-based species delimitation in Madagascar's mouse lemurs. In: Molecular Ecology 2016, Band 25, S. 2029–2045, doi:10.1111/mec.13604 (Erstbeschreibung);
- Robert D. Martin: A preliminary field-study of the lesser mouse lemur (Microcebus murinus J. F. Miller 1777). In: Zeitschrift für Tierpsychologie 1972, Beiheft 9, S. 43–89 (nicht eingesehen);
- Robert D. Martin: A review of the behaviour or the lesser mouse lemur (Microcebus murinus, J. F. Miller 1777). In: Richard P. Michael und John Hurrell Crook (Hrsg.): Comparative Ecology and Behaviour of Primates. Academic Press, London 1973, ISBN 0-12-493450-1, S. 1–68 (nicht eingesehen).
Weblinks
- ScienceDaily (15. April 2016): Three new primate species discovered in Madagascar
- Ganzhorn’s mouse-lemur (Microcebus ganzhorni) at Zoo Plzen, Video eines als Ganzhorns Mausmaki identifizierten Mausmakis im Zoo Pilsen auf der Website von Joel Sartore
- Ganzhorn’s mouse-lemur (Microcebus ganzhorni) at Zoo Plzen, weiteres Video Joel Sartores auf Vimeo
Einzelnachweise
- Maki ganzhurnov (Microcebus ganzhorni), Website des Zoo Prag, abgerufen am 25. März 2019.
- Petra Lahann, Jutta Schmid und Jörg U. Ganzhorn: Geographic Variation in Populations of Microcebus murinus in Madagascar: Resource Seasonality or Bergmann’s Rule? In: International Journal of Primatology 2006, Band 27, Nr. 4, doi:10.1007/s10764-006-9055-y.
- Petra Lahann: Feeding ecology and seed dispersal of sympatric cheirogaleid lemurs (Microcebus murinus, Cheirogaleus medius, Cheirogaleus major) in the littoral rainforest of south-east Madagascar. In: Journal of Zoology 2007, Band 271, S. 88–98, doi:10.1111/j.1469-7998.2006.00222.x.
- Brigitte M. Raharivololona, Rakotondravao und Jörg U. Ganzhorn: Gastrointestinal parasites of Small Mammals in the Littoral Forest of Mandena. In: Jörg U. Ganzhorn, Steven M. Goodman und Manon Vincelette (Hrsg.): Biodiversity, Ecology, and Conservation of Littoral Ecosystems in the Region of Tolagnaro (Fort Dauphin), Southeastern Madagascar. Smithsonian Institution, Washington, D.C. 2007, ISBN 978-1-893912-00-7, S. 247–258.
- Jutta Schmid und Jörg U. Ganzhorn: Optional strategies for reduced metabolism in gray mouse lemurs. In: Naturwissenschaften 2009, Band 96, S. 737–741, doi:10.1007/s00114-009-0523-z.
- Jean-Baptiste Ramanamanjato und Jörg U. Ganzhorn: Effects of forest fragmentation, introduced Rattus rattus and the role of exotic tree plantations and secondary vegetation for the conservation of an endemic rodent and a small lemur in littoral forests of southeastern Madagascar. In: Animal Conservation 2001, Band 4, S. 175–183, doi:10.1017/S1367943001001202.
- Alain G. Chabaud, Edouard-R. Brygoo und Annie-J. Petter: Les nématodes parasites de lémuriens malgaches. VII. Description de six espèces nouvelles et conclusions générales. In: Annales de Parasitologie 1965, Band 40, Nr. 2, S. 181–214, Digitalisat.
- Brigitte M. Raharivololona: Gastrointestinal parasites of Cheirogaleus spp. and Microcebus murinus in the littoral forest of Mandena, Madagascar. In: Lemur News. The Newsletter of the Madagascar Section of the IUCN/SSC Primate Specialist Group Juni 2006, Nr. 11, S. 31–35, Digitalisat.
- Scott Hotaling et al.: Species discovery and validation in a cryptic radiation of endangered primates: coalescent-based species delimitation in Madagascar's mouse lemurs. In: Molecular Ecology 2016, Band 25, S. 2029–2045, doi:10.1111/mec.13604.
- Timothy M. Eppley et al.: Ecological Flexibility as Measured by the Use of Pioneer and Exotic Plants by Two Lemurids: Eulemur collaris and Hapalemur meridionalis. In: International Journal of Primatology 2017, Band 38, Nr. 2, S. 338–357, doi:10.1007/s10764-016-9943-8.
- B. Karina Montero: Challenges of next‐generation sequencing in conservation management: Insights from long‐term monitoring of corridor effects on the genetic diversity of mouse lemurs in a fragmented landscape. In: Evolutionary Applications 2019, Band 12, S. 425–442, doi:10.1111/eva.12723.
- Jörg U. Ganzhorn et al.: Population Genetics, Parasitism, and Long-Term Population Dynamics of Microcebus murinus in Littoral Forest Fragments of South-Eastern Madagascar. In: Judith Masters, Marco Gamba und Fabien Génin (Hrsg.): Leaping Ahead. Advances in Prosimian Biology. Springer Science+Business Media, New York 2013, ISBN 978-1-4614-4510-4, S. 61–69.
- Peter M. Kappeler et al.: Long-term field studies of lemurs, lorises, and tarsiers. In: Journal of Mammalogy 2017, Band 98, Nr. 3, S. 661–669, doi:10.1093/jmammal/gyx013.
- Casey M. Setash et al.: A biogeographical perspective on the variation in mouse lemur density throughout Madagascar. In: Mammal Review 2017, Band 47, S. 212–229, doi:10.1111/mam.12093.
- Giuseppe Donati et al.: Lemurs in Mangroves and Other Flooded Habitats. In: Katarzyna Nowak, Adrian A. Barnett, Ikki Matsuda (Hrsg.): Primates in Flooded Habitats. Cambridge University Press, Cambridge 2019, S. 29–32, doi:10.1017/9781316466780.006.
- David W. Weisrock et al.: Delimiting Species without Nuclear Monophyly in Madagascar’s Mouse Lemurs. In: PLoS ONE 2010, Band 5, Nr. 3, Artikel e9883, doi:10.1371/journal.pone.0009883.
- Frank-Thorsten Krell und Stephen A. Marshall: New Species Described From Photographs: Yes? No? Sometimes? A Fierce Debate and a New Declaration of the ICZN. In: Insect Systematics and Diversity 2017, Band 1, Nr. 1, S. 3–19, doi:10.1093/isd/ixx004.
- Birgit Kruse: Microcebus ganzhorni: Neue Affenart nach Biologen der Universität Hamburg benannt. Universität Hamburg, Pressemitteilung vom 15. April 2016 beim Informationsdienst Wissenschaft (idw-online.de), abgerufen am 15. April 2016.
- Anonymus: Seznam zvířat chovaných v zoo a bz města Plzně v roce 2017. Census of animals kept in Pilsen zoo by the end of 2017 year, S. 3. In: Zoo Pilsen (Hrsg.): Zoologická a botanická zahrada města Plzně / výroční zpráva. Zoological and Botanical Garden Pilsen / Annual Report 2017, Digitalisat, abgerufen am 25. März 2019.
- Appendices I, II and III valid from 4 October 2017, Website der Convention on International Trade in Endangered Species of Wild Fauna and Flora, abgerufen am 26. März 2019.
- Laza Andriamandimbiarisoa et al.: Habitat corridor utilization by the gray mouse lemur, Microcebus murinus, in the littoral forest fragments of southeastern Madagascar. In: Madagascar Conservation & Development 2015, Band 10, Nr. 3, S. 144–150, doi:10.4314/mcd.v10i3.7.