Euthycarcinoidea
Die Euthycarcinoidea sind eine selten gefundene Gruppe ausgestorbener Gliederfüßer, von der Funde vom jüngsten Kambrium bis zur mittleren Trias vorliegen. Bisher sind 16 Arten beschrieben worden. Obwohl einige Fossilien mit exzellenter Erhaltung vorhanden sind, ist ihre genaue taxonomische Zuordnung bis heute rätselhaft, vorgeschlagen wurde unter anderem eine Zugehörigkeit zur Stammgruppe der Tausendfüßer, zu den Krebstieren oder als Schwestergruppe der Hexapoda. Während die ältesten bekannten Vertreter aus marinen Sedimenten stammen und wohl im Meer lebten, stammen alle späteren Fossilien aus limnischen oder terrestrischen Ablagerungen. Fossilierte Fußspuren (Spuren- oder Ichnofossilien) aus dem späten Kambrium und Ordovizium, die der Gruppe zugeordnet werden, gelten teilweise als die ältesten unzweideutigen Nachweise für Leben an Land.
| Euthycarcinoidea | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
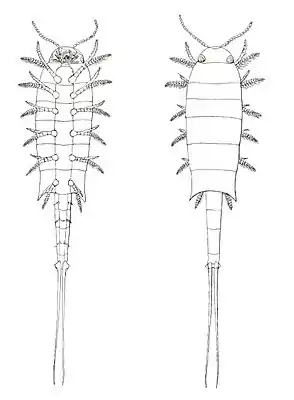
Euthycarcinus kessleri, Rekonstruktionszeichnung von Anton Handlirsch 1914 (heutige Rekonstruktionen in zahlreichen Details abweichend) | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Furongium bis Anisium | ||||||||||||
| 485 bis 247 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
|
Europa (Großbritannien, Frankreich, Deutschland), Nord- und Südamerika, Australien | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Euthycarcinoidea | ||||||||||||
| Gall & Grauvogel, 1964 | ||||||||||||
Körperbau
Es handelte sich um mittelgroße, abgeflachte Arthropoden. Kleinste Art war Heterocrania rhyniensis mit ca. 15 Millimeter Körperlänge.[1] Schramixerxes gerem aus Montceau-les-Mines erreichte 60 Millimeter Körperlänge. Größte bisher beschriebene Art ist Mosineia macnaughtoni mit etwa 10 Zentimeter.[2]
Der Körper der Euthycarcinoidea war in drei gegeneinander abgesetzte Körperabschnitte (Tagmata) gegliedert: Kopf, Präabdomen (manchmal auch Thorax oder Rumpf) und Postabdomen. Der Kopf besteht bei genauerer Betrachtung aus zwei Abschnitten. Der vordere, das Procephalon, trug zwei lange, einfache Gliederantennen und zwei halbkugelige "sphäroidale Fortsätze", von denen fast alle Bearbeiter meinen, dass es sich um Augen gehandelt hat (obwohl bis 2019 niemals Facetten beobachtet worden sind). Der hintere, Gnathocephalon genannt, trug die Mundgliedmaßen. Diese bestanden meist aus einem, selten zwei Paar meist plattenförmiger Strukturen, bei denen es sich nach der gängigen Interpretation um Mandibeln gehandelt hat, bei einigen Arten einer in der Kopfmitte (unpaarig) angeordneten Platte, die nach Ansicht einiger Bearbeiter dem Labrum der Insekten homolog sein könnte. Bei einigen Fossilien ist dies die einzige erkennbare Struktur, Mandibeln fehlen dann ganz; bei anderen Fossilien sind Mundwerkzeuge erhaltungsbedingt nicht erkennbar. Der mittlere Abschnitt, das Präabdomen, war auf der Ober- und Unterseite von Skleriten bedeckt. Die oberen Tergite waren dabei erheblich breiter als die unteren Sternite und bedeckten die Beine von oben fast vollständig. Seitlich an jedem Sternit saß ein Beinpaar, das vermutlich mit einem Apodem im Körper verankert war. Die Anzahl der Sternite, und damit der Beine, und die der Tergite stimmte dabei nicht überein, pro Tergit waren zwei oder bei den hinteren Segmenten manchmal drei Beinpaare vorhanden, häufig waren es elf Beinpaare. Die Beine waren einästig (uniram), sie waren in zahlreiche (bis zu 14) einfache, untereinander gleichförmige, ringförmige Segmente gegliedert, die zur Spitze hin etwas kleiner wurden, und endeten in einem Endstachel. Bei einigen Arten trugen sie feine lange Borsten (Kottyxerxes gloriosus)[3] oder je einen kurzen, stachelartigen Fortsatz.[1] Der hintere Körperabschnitt, das Postabdomen, war schmaler als das Präabdomen und bestand meist aus fünf, manchmal aus vier oder sechs Segmenten ohne Beine oder andere Anhänge und einem oft Telson genannten langen Endstachel variabler Länge. Bei den Gattungen Sottyxerxes und Pieckoxerxes (gefunden im Karbon der Lagerstätten Montceau-les Mines, Frankreich und Mazon Creek, USA) war das Präabdomen langgestreckt, relativ schmal und trug 14 Tergite und 28 Laufbeinpaare, die Tiere ähnelten dadurch rezenten Tausendfüßern. Bei den übrigen Gattungen war es wesentlich breiter als das Postabdomen, manchmal, so bei den Gattungen Euthycarcinus und Heterocrania seitlich hinten zahnförmig vorgezogen. Meist waren fünf Tergite erkennbar, bei manchen Gattungen (Mosineia) bis zu sieben; auch die Zahl der Beinpaare war variabel.
Lebensweise
Der Körperbau und die Fundumstände deuten darauf hin, dass die Euthycarcinoidea bodenlebende (benthische) Organismen waren, die in der flachen Uferzone von Gewässern lebten. Die Arten mit fein beborsteten Beinen waren möglicherweise auch schwimmfähig. Nach der Begleitflora und -fauna waren die besiedelten Gewässer in fast allen Fällen Süßwasser oder Brackwasser-Lebensräume. Bei der Art Apankura machu aus dem Kambrium Argentiniens[4] und den in Quebec gefundenen Arten,[2] also gerade bei den frühesten bekannten Vertretern der Gruppe, ist aber unzweifelhaft von einem marinen Lebensraum auszugehen. Die Gruppe stammt also ursprünglich aus dem Meer und scheint sich später in limnische Habitate zurückgezogen zu haben. Bei Tieren der Art Heterocrania rhyniensis aus dem Windyfeld Chert in Schottland (ganz nahe der berühmten Fossillagerstätte Rhynie) wurde ein fossilierter Darminhalt gefunden. Dieser besteht aus faserigem Material unzweifelhaft pflanzlicher Herkunft. Aufgrund der Dornfortsätze der Beine wird aber auch über eine mögliche räuberische Ernährung spekuliert. Für andere Arten wird aufgrund der feinen Haarborsten an den Beinen eine filtrierende Ernährung erwogen.[1]
Spurenfossilien
In fossil erhaltenen Böden von marinem Gezeitenwatt, zum Beispiel aus Kanada und Australien, sind Spurenfossilien bekannt, die aus seitlichen Abdrücken zahlreicher Beinpaare und einer zentralen Schleifspur bestehen. Diese sind als Parataxa je nach Ausprägung mit zahlreichen Namen versehen worden (Cruziana, Didymaulichus, Diplichnites, Protichnites). Diese können in vielen Fällen mit hoher Wahrscheinlichkeit auf Euthycarcinoidea bezogen werden.[5][6][7] Obwohl es nicht mit Sicherheit möglich ist, Spurenfossilien in sehr flachem Wasser von solchen an Land zu unterscheiden, ist die verbreitetste Interpretation diejenige, dass die Tiere das Wasser verlassen haben, entweder, um Algenwatten abzuweiden, oder um von einem flachen Gezeitentümpel zum nächsten zu wandern. Zu diesem Zeitpunkt liegen sonst keine Fossilien oder Spurenfossilien von landlebenden Tieren vor. Möglicherweise waren die Euthycarcinoidea daher die ersten Tiere, die das Wasser der Ozeane verlassen haben, um zumindest kurze Ausflüge an Land zu unternehmen. Zumindest bei zwei Arten, Synaustrus brookvalensis und Kottyxerxes gerem wurden aber auch einfache tracheenartige Strukturen beschrieben, die auf Luftatmung hindeuten könnten.
Aus dem Buntsandstein Deutschlands sind Ruhespuren von Arthropoden auf der Sedimentoberfläche als Tripartichnus triassicus beschrieben worden. Obwohl es naturgemäß nicht eindeutig möglich ist, den Spurenerzeuger zu identifizieren, sprechen die Indizien auch hier für einen Vertreter der Gruppe, möglicherweise Euthycarcinus kessleri.[8]
Verwandtschaft
Als Anton Handlirsch den ersten Vertreter der Gruppe, Euthycarcinus kessleri, aus dem Buntsandstein aus der Nähe von Saarbrücken beschrieb, nahm er an, dass er zu einer ausgestorbenen Ordnung der Krebstiere mit Beziehung zu den Ruderfußkrebsen gehörte.[9] Gall und Grauvogel beschrieben diese Art neu, nun als Vertreter einer eigenen Ordnung der Krebstiere, für die sie den Namen Euthycarcinoidea einführten. Spätere Bearbeiter vermuteten Beziehungen zu den Merostomata (Pfeilschwanzkrebse und Seeskorpione), die zu den Spinnentieren gehören, andere sahen sie unter anderem wieder als Krebstiere[10] oder als urtümliche Verwandte und mögliche Schwestergruppe der Hexapoda.[3]
Eine phylogenetische Analyse anhand morphologischer Merkmale[4] ergab eine Position basal innerhalb der Mandibulata, mit Krebstieren, Tausendfüßern und Sechsfüßern gemeinsam als Schwestergruppe. Ihre tatsächliche Position ist aber unklar und hängt in kritischer Weise vor allem von der Interpretation des Kopfs mit seinen Anhängen ab. Sie sind daher, dieser Analyse zufolge, als Arthropoda incertae sedis, vermutlich zu den Mandibulata gehörig, zu betrachten.
Eine Arbeitsgruppe um Gregory D. Edgecombe vom Londoner Natural History Museum hat 2019 Heterocrania rhyniensis aus dem Schottischen Rhynie Chert neu untersucht. Sie fanden bisher übersehene morphologische Merkmale: So besaß die Art eindeutig Facettenaugen. Am Kopf waren neben den Mandibeln als einzigen echten Mundwerkzeugen und dem Hypopharynx paarige Fortsätze, die als Superlinguae anzusprechen sind; Auswüchsen des Sternums des Mandibelsegments bei Tausendfüßern und primitiven Hexapoda. Im Kopfinneren wurden Teile des Tentoriums, einer Struktur der Kopfinnenskeletts, nachgewiesen. Einen weiteren Sklerit interpretieren sie als mögliches Labrum. Auf Basis der neuen Erkenntnisse favorisieren die Autoren ein Schwestergruppen-Verhältnis der Euthycarcinoidea mit den Myriapoda oder Tausendfüßern. Eine Zugehörigkeit zur Stammgruppe der Tausendfüßer hätte wichtige Konsequenzen zu unserer Kenntnis von deren Evolution. Damit wäre nicht nur die Existenz dieser Klade bis ins Kambrium erstmals klar nachgewiesen, sondern auch die lange gesuchte marine, aquatische Stammgruppe der Myriapoda eingegrenzt.[11]
Quellen
- Javier Ortega-Hernández, David A. Legg, Jonathan Tremewan, Simon J. Braddy (2010): Euthycarcinoids (Fossils explained 59). Geology Today Vol. 26, No. 5: 195–198.
- Patrick R. Racheboeuf, Jean Vannier, Frederick R. Schram, Dominique Chabard, Daniel Sotty (2008): The euthycarcinoid arthropods from Montceau-les-Mines, France: functional morphology and affinities. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 99: 11–25. doi:10.1017/S1755691008006130
Einzelnachweise
- Lyall I.Anderson, & Nigel H. Trewin (2003): An Early Devonian arthropod fauna from the Windyfield cherts, Aberdeenshire, Scotland. Palaeontology, Volume 46, Number 3: 467-509.
- Joseph H. Collette: Three-Dimensionally Preserved Arthropods from the Cambrian (Furongian) of Quebec and Wisconsin: Systematics, Phylogeny, Ichnology, and Taphonomy. Masters Thesis 1896 - February 2014. Paper 322. download
- Frederick R. Schram & W. D. Ian Rolfe (1982): New Euthycarcinoid Arthropods from the Upper Pennsylvanian of France and Illinois. Journal of Paleontology Vol. 56, No. 6: 1434-1450.
- N.E. Vaccari, G.D. Edgecombe, C. Escudero (2004): Cambrian origins and affinities of an enigmatic fossil group of arthropods. Nature vol. 430: 554-557.
- Gregory J. Retallack (2008): Cambrian–Ordovician non-marine fossils from South Australia. Alcheringa 33 (4): 355-391. doi:10.1080/03115510903271066
- Joseph H. Collette, James W. Hagadorn, Mario A. Lacelle (2010): Dead in their tracks — Cambrian arthropods and their traces from intertidal sandstones of Quebec and Wisconsin. Palaios 25: 475–486. doi:10.2110/palo.2009.p09-134r
- Joseph H. Collette, Kenneth C. Gass,James W. Hagadorn (2012): Protichnites eremita Unshelled? Experimental Model-Based Neoichnology and New Evidence for A Euthycarcinoid Affinity for This Ichnospecies. Journal of Paleontology vol. 86: 442-454. doi:10.1666/11-056.1
- Lothar H. Vallon & Martin Röper (2008): Spuren lesen - Vom Abdruck zum Erzeuger. Fossilien 2008: 230-234. download (Memento des Originals vom 19. Oktober 2014 im Internet Archive) Info: Der Archivlink wurde automatisch eingesetzt und noch nicht geprüft. Bitte prüfe Original- und Archivlink gemäß Anleitung und entferne dann diesen Hinweis.
- Anton Handlirsch (1914): Eine interessante Crustaeeenform aus der Trias der Vogesen. Verhandlungen der Zoologisch-Botanischen Gesellschaft in Wien 64: 1-8. PDF
- Jean-Claude Gall & Léa Grauvogel-Stamm (2005): The early Middle Triassic ‘Grès à Voltzia’ Formation of eastern France: a model of environmental refugium. Comptes Rendus Palevol 4: 637–652. doi:10.1016/j.crpv.2005.04.007
- Gregory D. Edgecombe, Christine Strullu-Derrien, Tomasz Góral, Alexander J. Hetherington, Christine Thompson, Markus Koch (2020): Aquatic stem group myriapods close a gap between molecular divergence dates and the terrestrial fossil record. PNAS Proceedings of the National Academy of Sciences USA 117 (16): 8966–8972. doi:10.1073/pnas.1920733117