Euploea core
Euploea core sensu stricto ist eine von mindestens fünf allopatrisch vorkommenden Schmetterlingsarten der Superspezies Euploea core, (Tagfalter), die in diesem Fall durch das Meer voneinander getrennt sind. Sie wurden bis in die 1990er Jahre zusammen als die Art Euploea core betrachtet und der Name wird weiterhin für die aus diesen fünf Arten gebildete Superspezies benutzt. Die Falter sind von Südasien bis Australien in unterschiedlichsten Lebensräumen weit verbreitet und häufig. Durch die Aufnahme von Herzglykosiden aus den Nahrungspflanzen im Raupenstadium können die Raupen, Puppen und schwarzbraunen Falter leicht giftig sein. Dadurch sind sie etwas vor den typischen Fressfeinden aus der Gruppe der Wirbeltiere wie Vögel, Mäuse und Echsen geschützt. Die Raupen ernähren sich von einer Vielzahl verschiedener Pflanzenarten, besonders von Apocynaceae, Seidenpflanzengewächsen (Asclepiadaceae) und Maulbeergewächsen (Moraceae).
| Euploea core | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
_on_Indian_Turnsole_(Heliotropium_indicum)_W_IMG_9747.jpg.webp)
Euploea core | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Euploea core | ||||||||||||
| (Cramer, 1780) |
_-_cprogrammer.jpg.webp)
Merkmale
Imago
Euploea core ist ein Schmetterling mit glänzenden dunkelbraunen bis schwarzen Flügeln und einer Flügelspannweite von sieben bis neun Zentimetern. Der schwarze Körper der Imago ist mit weißen Punkten gesprenkelt.
Die Flügeloberseite von Euploea core (s. str.) ist dunkelbraun und wird zum Außenrand, in der Diskalregion, heller. Entlang der Außenränder verlaufen zwei Reihen weißer Flecken. Auf dem Vorderflügel biegt sich die innere Reihe mit den größeren Flecken an der Flügelspitze (Apex) nach innen zum Vorderflügelrand. Die äußere Reihe mit den kleineren Flecken ist oft lückenhaft und erreicht oft nicht den Apex. Häufig ist ein kleiner Punkt am Vorderrand (Costa) und nur sehr selten ist ein Punkt im Apex der Zelle und in der Diskalregion vorhanden. Auf der Hinterflügeloberseite sind die Flecken der inneren Reihe länglich und die der Äußeren konisch. Die großen weißen Flecke sind oft nur durch die dunklen Flügeladern getrennt und bilden ein Band. Die Unterseite ist ähnlich, die Grundfarbe ist gleichmäßiger und die Punkte an der Costa, in der Zelle und der Diskalregion sind fast immer vorhanden. Die Männchen unterscheiden sich von den Weibchen durch einen länglichen Duftschuppenfleck in Zelle 2 auf der Vorderflügeloberseite.
Die Männchen haben auffällige ausstülpbare gelbe Haarbüschel am Abdomen, die Härchen sprießen über fast die gesamte Länge aus dem Haarbüschelschaft. Dies ist ein typisches Merkmal für die unter der Klade Euploeina zusammengefassten Gattungen Euploea, Anetia, Lycorea, Idea, Protoploea. Im Gegensatz dazu habe die anderen Gattungen der Danaini, die unter der Klade Danaina zusammengefasst werden, Härchen nur aus dem hinteren Drittel des Haarbüschelschafts sprießen. Die weiblichen Falter haben eine leuchtend gelbe Geschlechtsöffnung (Ostium), die die Haarbüschel nachahmt.[1][2]
Unterarten und Formen
Die Flecken der inneren Reihe auf den Vorderflügeln der Form E. core f. vermiculata sind größer und entweder quadratisch oder herzförmig. Der Übergang dieser in den Vorbergen und niederen Lagen des Himalaya lebenden Form zur Nominatform ist fließend. E. core f. godarti, Lucas, 1853, ist von Manipur über Cachar bis Burma und Indochina verbreitet und hat einen violetten Apex am Vorderflügel. In Burma tritt E. core f. layardi mit braunem Apex hat am Vorderflügel auf. Die Zwischenformen mit f. godarti werden als f. defigurata bezeichnet.[3]
Die Unterart E. core asela ist größer und unterscheidet sich von der Nominatform durch kleinere Flecken in beiden Reihen auf den Vorderflügeln, die mehr oder weniger verschwinden.[4]
Die auf Kalao beschränkte Unterart E. core kalaona ist schwarz mit nur kleinen weißen Flecken. Nur am Apex befinden sich zwei große bis zu den Flügeladern reichende Flecken. Nach der Zelle sind drei Flecken nur schwach angedeutet. Auf den Hinterflügeln fehlen die Flecken der inneren Reihe fast gänzlich. Der Duftschuppenfleck ist auffällig lang und gleicht dem von Euploea algea.[5]
E. core bauermanni ist auf Salajar beschränkt und ähnelt E. core kalaona. Die Unterart ist kleiner und die drei Flecken nach der Zelle sind gut ausgebildet.
Allopatrische Arten

Euploea andamanensis ist hellbraun und der kleine Punkt an der Costa, der Punkt im Apex der Zelle und die Punkte in der Diskalregion sind deutlich ausgeprägt und groß.
Euploea scherzeri ist ganz schwarz. Nur wenige Falter haben einige kleine weiße, unregelmäßig angeordnete, Flecken in der Diskalregion auf der Flügelunterseite.
Euploea corinna hat auffällige weiße Flecken in der Vorderflügelspitze und der Postdiskalregion der Vorderflügel. Auf den Flügelunterseiten befinden sich zusätzlich kleine, unregelmäßig angeordnete, weiße Flecke in der Diskalregion.
Die kleine Unterart Euploea charox renellensis unterscheidet sich von E. corinna durch rundere Flügel und größere Duftschuppenflecken. Auf der Hinterflügeloberseite ist nur die innere Reihe der submarginalen Flecken vorhanden. Diese sind deutlich größer als bei E. corinna und reichen fast bis zum Analwinkel. Die submarginalen Flecken auf den Vorderflügeln sind ebenfalls deutlich größer und Näher am Rand als bei E. corinna. In der Regel sind die Flecken bei den Weibchen größer als bei den Männchen.[5]
Ähnliche Arten
Die Männchen von Euploea sylvester unterscheiden sich durch zwei Duftschuppenflecken in Zelle 1b und 2 auf den Vorderflügeln statt nur einem. Bei den Weibchen ist die Form der Vorderflügel leicht unterschiedlich. Ansonsten gleichen sich die beiden Arten in ihrem Erscheinungsbild.[4]
Die Abbildungen aus Binghams The Fauna of British India including Ceylon and Burma von 1905 zeigen die Flügeloberseiten der Männchen von Euploea core (s. str.), Euploea core f. godarti (damals Euploea godarti) und Euploea sylvester coreta (damals Euploea coreta). Der eine Duftschuppenfleck und besonders im Falle von Euploea sylvester die zwei langen sind am Innenrand der Vorderflügel erkennbar, ebenso wie die große Variabilität der Fleckenmuster innerhalb der Art Euploea core.
_-_Bingham.png.webp) Euploea core (s. str.)
Euploea core (s. str.) Euploea core f. godarti
Euploea core f. godarti_-_Bingham.png.webp) Euploea sylvester coreta
Euploea sylvester coreta
Mimikry
Euploea core dient anderen Schmetterlingsarten als Vorbild für Batessche Mimikry. Die Nachahmer wollen vom Schutz vor Feinden, den die leicht giftigen Falter haben, profitieren.
- Euploea core ist das Vorbild für die weiblichen Falter von Hypolimnas bolina in deren westlichen Verbreitungsgebiet.[6]
- Chilasa clytia form clythia aus der Familie der Ritterfalter ahmt Euploea core im westlichen Verbreitungsgebiet nach.[7]
 Chilasa clytia aus Bingham (damals Papilio clytia)
Chilasa clytia aus Bingham (damals Papilio clytia) Mimikry eines Hypolimnas bolina Weibchens
Mimikry eines Hypolimnas bolina Weibchens
Ei
Die hellgelben Eier sind relativ groß, annähernd fassförmig, deutlich höher als breit und haben eine runde Spitze. Die Oberfläche ist durch Längs- und Querrippen strukturiert.
Raupe
Die Raupen werden bis zu 6 Zentimeter lang und haben acht dunkelviolette fühlerförmige fleischige Fortsätze, die paarweise am Mesothorax, Metathorax und dem zweiten und achten Abdominalsegment sitzen.
Die älteren Raupen von Euploea core (s. str.) sind blauviolett und haben das typische Muster aus dunklen Querstreifen mit hellen Rändern auf dem Rücken. Die dunklere Farbe wird mit jeder Häutung dunkler, die helle Farbe ist cremeweiß bis weiß. An den Seiten lösen sich die Bänder auf und gehen in ein längsgerichtetes Band aus der dunkleren Farbe über, in dem die schwarz umrandeten Tracheen deutlich sichtbar sind. Unterhalb dieses Streifens verläuft ein deutlich abgegrenztes cremefarbenes Band. Die Beine und die Kopfkapsel sind schwarz, wobei letztere zusätzlich weiße Linien hat.
Bei Euploea corinna haben die älteren Raupen normalerweise statt einer blauvioletten eine orange bis braune Grundfarbe. Als junge Raupen haben sie einen schwarzen Kopf und die Färbung des Körpers reicht von gelb bis braun. Nur im Norden findet sich häufig in trockenen Gegenden eine blauviolette Form, die den Raupen von E. core gleicht. Selten werden blasse Raupen mit pastellfarbener Zeichnung gefunden.[4][8]
Die Raupen von Euploea charox sind hellbraun mit schwarzen und weißen Bändern.
Puppe
Die Stürzpuppen der Unterfamilie der Danainae sind für ihre glänzende Oberfläche bekannt. Die plumpe und weich gerundete Puppe von Euploea core ist keine Ausnahme und hat einen goldenen Glanz, der durch Interferenz an bis zu 250 gleichmäßig dünnen Schichten hervorgerufen wird. Der goldene Farbton entsteht durch Karotin; Puppen von Raupen, die karotinfrei gezüchtet werden, haben einen silbernen Glanz. Die Puppen von Euploea charox sind manchmal auch in der freien Natur silberfarben und haben graubraune Male.
Die Puppen werden trotz ihres auffallenden Äußeren nicht von Feinden angegriffen. Einerseits spiegelt sich in der glänzenden Oberfläche die Umgebung, was zur Tarnung beitragen kann. Andererseits hält Bernard d'Abrera die metallische glänzende Puppe auf Sri Lanka für sehr auffällig und vermutet, dass es sich um eine Warnfarbe handelt.[9]
 Ei von Euploea core an Blattunterseite
Ei von Euploea core an Blattunterseite Raupe von Euploea core
Raupe von Euploea core Präpuppe
Präpuppe Ältere Puppe mit dem typischen Goldglanz
Ältere Puppe mit dem typischen Goldglanz
Verbreitung
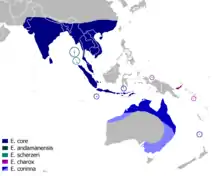
Euploea core (s. str.) ist von Indien, Sri Lanka, dem südlichen China über Indochina bis Sumatra, Java und den Kleinen Sunda-Inseln verbreitet. Durch Vorkommen auf Kalao (E. core kalaona) und Salajar (E. core bauermanni) reicht die Art bis an die südliche See vor Sulawesi heran. Auf den Kleinen Sunda-Inseln könnte auch schon Euploea corinna statt Euploea core vorkommen.
Euploea andamanensis ist auf die Andamanen beschränkt und Euploea scherzeri auf die südlich davon gelegenen Nikobaren.
Euploea charox kommt auf Biak und dem Bismarck-Archipel vor. Auf Rennell und vielleicht auch auf der Nachbarinsel Bellona, die zu den Salomonen gehören, kommt die Unterart Euploea charox renellensis vor.
Die Verbreitung von Euploea corinna in Australien erstreckt sich entlang der Küste von Pilbara in Nordwestaustralien über das Northern Territory und entlang der ganzen Ostküste bis Tasmanien. Außerdem kommt die Art auf der südlich von Java liegenden Weihnachtsinsel und auf der östlich von Australien liegenden Norfolkinsel vor.
Euploea core scheint nicht auf Borneo, den Philippinen, den Molukken, Sulawesi und Neuguinea vorzukommen.[10]
Der Falter wird in vielen Regionen als häufig bis sehr häufig beschrieben. In Indien und Ceylon ist er sehr häufig und wird auf Ceylon auch in Dörfern und Städten oft angetroffen. Ebenso ist er in China sehr häufig und in Australien ist er einer der häufigsten Schmetterlinge und häufiger als alle anderen dort vorkommenden Arten der Gattung Euploea zusammen.[5][11][12]
Lebensraum
Der Falter lebt im Gegensatz zu den meisten Arten der Gattung, die auf Wälder oder große Waldgebiete beschränkt sind, in einer Vielzahl von Lebensräumen. Sie reichen von Wäldern und bewaldeten Wasserläufen über offene Landschaften bis zu städtischen Grünanlagen. Die vertikale Verbreitung reicht von Mangrovensümpfen auf Meeresniveau bis auf 2500 Meter Höhe im nordwestlichen Himalaya. In Australien leben die Falter auch in kühlen Sand- und Kalksteinschluchten landeinwärts und in der Umgebung von natürlichen Quellen, obwohl sie die tropische und subtropische Region und die Küsten bevorzugen. In heißen und trockenen Regionen können sie nicht überleben.
Lebensweise
Imago
_-_Hanumann.jpg.webp)
Euploea core fliegt in Bodennähe, nur wenige Meter über dem Boden. Bei seinen energielos wirkenden Flügen segelt er oft. Die Falter lassen sich von Menschen nicht stören, sie scheinen sogar menschliche Gesellschaft zu mögen und sind Kulturfolger in Gärten.[11] Oft sitzen sie an Blüten oder sie versammeln sich an feuchten Stellen, Wurzeln und verletzten oder abgestorbenen Pflanzenteilen.
Die Männchen stülpen ihr Haarbüschel oft im Flug aus, auch wenn keine Weibchen in der Nähe sind. Normalerweise dienen die Haarbüschel, die ein Merkmal der Unterfamilie der Danainae darstellen, der Werbung um Weibchen und werden nur in deren Nähe ausgestülpt, um einen auch für Menschen angenehmen süßlichen Duft zu verbreiten. Bei der Gattung Euploea scheint dieses Verhalten noch eine territoriale Funktion zu haben.
Von vielen Arten der Unterfamilie der Danainae ist bekannt, dass besonders die männlichen Falter von verwelkten und verletzten Pflanzenteilen angelockt werden, die Pyrrolizidinalkaloide enthalten. Bei faulenden oder frisch abgeknickten Blattstielen saugen die Tiere direkt an den Pflanzen. Um die Pyrrolizidinalkaloide mit dem Rüssel von trockenen Pflanzenteilen aufzunehmen, erbrechen die Falter oft Flüssigkeit auf die Oberfläche der Pflanzen und saugen diese wieder auf. Manchmal kratzen sie zusätzlich an den Pflanzen. Dieses Verhalten kann auch bei Euploea core an Tournefortia argentea (Raublattgewächse (Boraginaceae)) und Sonnenwenden (Heliotropium indicum) beobachtet werden, wobei von letzterer ebenso viele weibliche wie männliche Falter angelockt werden.[13] Die Männchen bauen die Alkaloide chemisch zu Danaidon und ähnlichen Pheromonen um, die auch in ihren Haarbüscheln gefunden werden. Die Alkaloide machen die Falter beider Geschlechter ungenießbar, oft sind sie bereits durch die Aufnahme von Giften aus den Raupennahrungspflanzen leicht giftig.
Die Weibchen legen ihre Eier in Gruppen auf der Unterseite junger Triebe im oberen Bereich der Nahrungspflanzen ab, wo sich auch die meisten junge Triebe und Blüten befinden. Pflanzen mit schattigem Standort werden bevorzugt, vorher abgelegte Eier anderer Falter haben keinen Einfluss auf das Eiablageverhalten. An Oleander legen sie ihre Eier bevorzugt an der Unterseite von jungen Trieben und Blütenblättern ab, wobei die Blütenfarbe keinen Einfluss auf das Eiablageverhalten hat. Im Labor wurden Seidenpflanzengewächse (Asclepiadaceae), Apocynaceae, Maulbeergewächse (Moraceae), Oleander und Parsonsia straminea stark mit Eiern belegt, manchmal wurde sogar auf den für die Raupen ungeeigneten Pflanzen der Gattung Asclepias stark abgelegt.[14]
Aktive Falter leben bis zu 13 Wochen lang, überwinternde Falter erreichten in Queensland eine Lebensdauer von 30 Wochen.
Wanderverhalten
Euploea core tritt in verschiedenen Regionen der Welt als Wanderfalter in Erscheinung. Die Falter sammeln sich immer erst in größeren Gruppen, bevor sie gemeinsam wandern.[15]
Im südlichen Indien wandern die Falter zeitweise in die feuchten Urwälder des Tieflandes. Während der Mittagshitze sitzen die Falter in schattigen Bäumen. Mit dem Einsetzen des Nordost-Monsuns in Sri Lanka leiten E. core asela und Danaus hamata die beginnende Wandersaison der Schmetterlinge ein.[16]
In Australien ähnelt das Wanderverhalten von Euploea corinna dem des Monarchfalters. Die Falter wandern im Winter aus dem Süden nach Norden und versammeln sich an verschiedenen Plätzen an der Küste zwischen Brisbane und Townsville. Die Wahl der Plätze hängt vom Wetter und Nektar- und Wasserquellen ab. Die Tiere sind kälte- und hitzeempfindlich und müssen sich an windgeschützten Stellen während dieser winterlichen Diapause vor Austrocknung schützen. Auf einigen Inseln des Great Barrier Reef sammeln sie sich besonders häufig in geschützten Senken. Es wird vermutet, dass die Bildung von Eiweißreserven maßgeblich zum Überleben der Ansammlungen während der Trockenzeit im nördlichen Australien beiträgt, da sie während des Winters nur selten Nahrung aufnehmen.[17]
In Hongkong überwintert Euploea core und Euploea midamus in Ansammlungen zusammen mit einzelnen Faltern von Ideopsis similis, Tirumala limniace, Parantica aglea und Danaus genutia.[18]
Raupe

Die Raupen von Euploea core ernähren sich oft von giftigen Pflanzen und speichern diese Gifte, so dass auch die Schmetterlinge teilweise ungenießbar und damit geschützt vor Prädatoren sind. Die Aufnahme der Glykoside ist viel schwächer als bei dem zur selben Tribus gehörenden Monarchfalter und die Aufnahme der Glykoside reicht oft nicht aus, um bei Vögeln einen Brechreiz auszulösen. Wenn sich eine Raupe von ungiftigen Pflanzen ernährt, kann sie keine Gifte synthetisieren.[19][20] Bevor die Raupe an einem Blatt frisst, beißt sie bei Pflanzen mit hohem Milchsaftanteil zuerst den Blattstiel an, so dass das Blatt nach unten knickt. Damit wird die Zufuhr von Milchsaft in das Blatt unterbrochen. Die Gefahr, dass die Mandibeln durch den Milchsaft verkleben, wird dadurch vermindert. Dieses Verhalten kann schon zwei Stunden nach dem Schlüpfen der Eiraupe beobachtet werden. Im ersten Raupenstadium beißt sie meist eine Reihe bogenförmiger Adern am Blattrand durch, manchmal wird aber auch die Hauptader durchtrennt, wie es die älteren Raupen machen.[21]
Der ideale Temperaturbereich für die Raupen liegt zwischen 18 °C und 30 °C, werden diese Werte unter-, beziehungsweise überschritten, sterben viele Raupen.
Wenn die Raupe gestört wird, hebt sie ihr Vorderteil auf und neigt den Kopf nach unten. Dieses Verhalten ähnelt dem der Raupen aus der Familie der Schwärmer (Sphingidae).
Ein breites Spektrum von Pflanzen dient Euploea core als Nahrung, was wahrscheinlich mit ein Grund für seine Häufigkeit ist.[15] In Australien wird Oleander und Cryptostegia grandiflora, ein invasiver Neophyt mit starker Ausbreitungstendenz, erst in jüngster Zeit als Nahrungspflanze genutzt. Beide wurden erst durch die europäischen Einwanderer eingeschleppt und spielen in Australien eine große Rolle, da sie ein beständigeres Angebot als die ursprünglichen Nahrungspflanzen an für die Eiablage günstigen jungen Trieben liefern. Cryptostegia grandiflora ist häufig entlang von Straßen und Trockenflüssen zu finden und dort sind oft beachtliche Populationen von Euploea core anzutreffen.[8][22]
- Apocynaceae: Allemanda sp., Carissa ovatais, Holarrhena antidysenterica, Ichnocarpus frutescens, Mandevilla sp., Oleander (Nerium oleander), Parsonsia strarninea, Plumeria acutifolia, Rhyncospermum sp., Trachelospermum bowringii
- Seidenpflanzengewächse (Asclepiadaceae): Gomphocarpus physocarpus, Hoya australis, Ischnostemma carnosum, Leichardtia australis, Marsdenia leichhardtiana, Marsdenia rostrata, Marsdenia suaveolens, Sarcostemma australe, Secamone elliptica, Stephanotis sp., Toxocarpus wightianus
- Maulbeergewächse (Moraceae): Ficus australis, Banyan-Feige (Ficus benghalensis), Birkenfeige (Ficus benjamina), Ficus eugenioides, Ficus glomerata, Ficus indica, Chinesische Feige oder Lorbeer-Feige (Ficus microcarpa), Ficus muelleri, Ficus obliqua, Ficus pandurata, Ficus platypoda, Ficus religiosa, Ficus variolosa, Streblus asper
- Periplocaceae: Cryptolepis elegans, Cryptolepis pauciflora, Cryptostegia madagascariensis, Gymnanthera nitida, Hemidesmus indicus
- Rötegewächse (Rubiaceae): Morinda sp.
- Ulmengewächse (Ulmaceae): Girionniera reticulata
Teilweise werden die Eier an für die Raupen ungeeignete Pflanzen der Gattung Asclepias abgelegt. Die Raupen beginnen an den Pflanzen zu fressen, gehen aber nach wenigen Tagen ein. Die Ursache dafür ist nicht bekannt, nahe verwandte Pflanzen werden dagegen gut von den Raupen vertragen.[14] Unter Laborbedingungen überlebte von 50 Euploea core corinna-Raupen nur eine das erste Stadium an Gomphocarpus fruticosus, diese schaffte es aber dann bis zum Falter. Dagegen war die Mortalität an Oleander deutlich geringer, hier überlebten 94 Prozent das erste Stadium, wobei die Sterblichkeit in den Stadien vier und fünf stark zunahm und es etwa 50 Prozent bis zur Puppe schafften. An Birkenfeige überlebten mit 86 Prozent weniger Raupen das erste Stadium als an Oleander, die Sterblichkeit nahm allerdings nicht so stark zu, so dass sich etwa 75 Prozent verpuppten. An Parsonsia strarninea starben pro Stadium etwa 10 Prozent der Larven, so dass ebenso viele bis zur Puppe überlebten wie an Oleander.[23]
Parasiten
Mehrere Generalisten sind als Parasitoide der Raupen von Euploea core nachgewiesen. Dazu gehören die Raupenfliegen (Tachinidae) Paradrino laevicula, Sturmia flavohalterata und Winthemia neowinthemioides und der Hautflügler (Hymenoptera) Brachymeria lasus aus der Familie der Chalcididae. Die Eier werden nur sehr selten parasitiert, da sie wahrscheinlich wesentlich mehr Glykoside enthalten als die Raupen und damit besser geschützt sind.[8][24]
Systematik
Die Art wurde 1780 von Cramer als Papilio core anhand eines Falters von der ostindischen Küste erstbeschrieben. Der englische Trivialname des Falters lautet „Common Crow“ (Gemeine Krähe). In Indien wird er auch als „Common Indian Crow“ (Gemeine Indische Krähe) und in Australien als „Australian Crow“ (Australische Krähe) und „Oleander Butterfly“ (Oleanderfalter) bezeichnet.
In ihrem großen Verbreitungsgebiet wurde die Art, auch aufgrund ihrer variablen Erscheinung, mehrfach beschrieben. Viele dieser Beschreibungen werden als Unterarten betrachtet. Die ehemals unter der Art Euploea core zusammengefassten Populationen haben keine ausgeprägten gemeinsamen Merkmale, stattdessen waren das Fehlen von Merkmalen, die eine Zuordnung zu anderen Arten erlauben würden, und die getrennten Verbreitungsgebiete der Grund für deren Einordnung in diese Art, was aber immer wieder in Frage gestellt wurde. 1993 erhob Richard I. Vane-Wright vier der geographisch abgetrennten Unterarten zu Arten, womit die neue Superspezies aus den fünf Arten E. core (s. str.), E. andamanensis, E. scherzeri, E. charox und E. corinna besteht. E. core renellensis wird als Unterart von E. charox betrachtet, könnte jedoch auch eine weitere Art sein.[10]
Nach der Aufteilung in fünf Arten ergibt sich folgende Systematik mit den auch hier erwähnten Unterarten:
- E. core Cramer, 1780
- E. core asela (Moore), 1877
- E. core kalaona (Fruhstorfer), 1898
- E. core bauermanni (Röber), 1885
- E. andamanensis, Atkinson, [1874]
- E. scherzeri C. Felder, 1862
- E. charox Kirsch, 1877
- E. charox renellensis (Carpenter), 1953
- E. corinna (Macleay), 1826,
Quellen
- Milkweed Butterflies, S. 76
- Milkweed Butterflies, S. 40
- Milkweed Butterflies, S. 54
- C. T. Bingham: Butterflies. In: The Fauna of British India including Ceylon and Burma. Band 1. London 1905, S. 32 f.
- Bernard d'Abrera: Butterflies of the Australian Region. 3. Auflage. Hill House Publishers, Melbourne/London 1990, ISBN 0-7018-1003-3, S. 181–183.
- N. A. Marsh, C. A. Clarke, Miriam Rothschild & D. N. Kellett: Hypolimnas bolina (L.), a mimic of danaid butterflies, and its model Euploea core (Cram.) store cardioactive substances. In: Nature. Band 268, 25. August 1977, S. 726–728, doi:10.1038/268726a0.
- Reginald Crundall Punnett: Mimicry in Butterflies. Cambridge University Press, 1915, S. 25.
- Biology of the Australian Butterflies, S. 191ff
- Milkweed Butterflies, S. 72
- R.I. Vane-Wright & R. de Jong: The butterflies of Sulawesi: annotated checklist for a critical island fauna. In: Zool. Verh. Band 343. Leiden 2003, ISBN 90-73239-87-7, S. 227 (naturalis.nl [PDF; 3,5 MB; abgerufen am 28. Februar 2008]).
- Bernard d'Abrera: The Butterflies of Ceylon. Hill House Publishers, Armadale (Victoria) 1998, ISBN 0-947352-35-X, S. 112.
- Paul Lau: Butterflies of Hong Kong. Hongkong 1997, S. 156.
- Milkweed Butterflies, S. 78
- G. Daglish, M. P. Zalucki, I. Gynther, H. Rahman, E. Scheermeyer & M. Arura: Field and Laboratory Observations on Oviposition by Euploea core (Insecta, Lepidoptera). In: Australian Journal of Zoology. Band 34, Nr. 6. CSIRO PUBLISHING, Collingwood, Victoria 1986, S. 827–836, doi:10.1071/ZO9860827.
- Gay, Kehimkar & Punetha: Common Butterflies of India. Hill House Publishers, Oxford 1992, ISBN 0-19-563164-1, S. 26.
- Williams, Carrington Bonsor: Die Wanderflüge der Insekten: Einführung in das Problem des Zugverhaltens der Insekten unter besonderer Berücksichtigung der Schmetterlinge. Paul Parey, Hamburg, Berlin 1961, S. 55.
- Milkweed Butterflies, S. 228
- Milkweed Butterflies, S. 102
- Miriam Rothschild, Neville Marsh & Brian Gardiner: Cardioactive substances in the Monarch butterfly and Euploea core reared on leaf-free artificial diet. In: Nature. Band 275, 19. Oktober 1978, S. 649–650, doi:10.1038/275649a0.
- Milkweed Butterflies, S. 81
- Clarke, Anthony R.; Zalucki, Myron P.: Foraging and vein-cutting behaviour of Euploea core corinna (W. S. Macleay) (Lepidoptera: Nymphalidae) caterpillars feeding on latex-bearing leaves. In: Australian Journal of Entomology. Band 39, Nr. 4. Blackwell Publishing, Oktober 2000, S. 283–290, doi:10.1046/j.1440-6055.2000.00191.x (HTML [abgerufen am 20. Oktober 2008]).
- Milkweed Butterflies, S. 229
- Habib Ur Rahman, Myron P. Zaluckai & Elly Scheermeyer: The Effect of Host Plant on the Development and Survival of the Immature Stages of Euploea core corinna (Lepidoptera Nymphalidae). In: Australian Journal of Entomology. Band 24, Nr. 2, Mai 1985, S. 95–98, doi:10.1111/j.1440-6055.1985.tb00195.x.
- J. McAuslane & Fred D. Bennett: Parasitoids and Predators Associated with Syntomeida epilais (Lepidoptera: Arctiidae) on Oleander Heather. In: Florida Entomologist. Band 78, Nr. 3, September 1995, S. 543 ff. (fcla.edu [PDF]).
Literatur
- Richard Irwin Vane-Wright & P. R. Ackery (Hrsg.): Milkweed Butterflies. Their Cladistics and Biology. Cornell University Press, Ithaca and London 1984, ISBN 0-8014-1688-4.
- Michael F. Braby: Butterflies of Australia, CSIRO Publishing, Melbourne 2000, Band 2, S. 605–607.
- R. L. Kitching & Myron P. Zalucki: Observations on the Ecology of Euploea core corinna (Nymphalidae) with special reference to an overwintering population, Journal of the Lepidoptera Society, Volume 35, 1981, S. 106–119.
- R. L. Kitching, Dr. E. Scheermeyer, R. E. Jones and N. E. Pierce (Hrsg.): The Biology of the Australian Butterflies, CSIRO Publishing, Melbourne 1999, Kapitel 9.
- Elly Scheermeyer: Overwintering of Three Australian Danaines: Tirumala hamata, Euploea tulliolus tulliolus and Euploea core corinna in S. B. Malcolm and M. P. Zalucki (Hrsg.): Biology and Conservation of the Monarch Butterfly, Natural History Museum of Los Angeles County, Los Angeles 1993, S. 345–354.
Weblinks
- Common Australian Crow – Euploea core corinna (englisch)
- Butterflies take a winter break – Interview mit Myron Zalucki (englisch)
- Euploea core in der Roten Liste gefährdeter Arten der IUCN 2013.2. Eingestellt von: Muller, C.J. & Tennent, W.J., 2009. Abgerufen am 18. Februar 2014.