Amargasaurus
Amargasaurus ist eine Gattung sauropoder Dinosaurier, die während der Unterkreide in Südamerika lebten.
| Amargasaurus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettabguss im Foyer des Melbourne Museum | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Unterkreide (Barremium bis Unteres Aptium)[1] | ||||||||||||
| 130,7 bis 123 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Amargasaurus | ||||||||||||
| Salgado & Bonaparte, 1991 | ||||||||||||
| Art | ||||||||||||
|


Bisher ist ein einziges, nahezu vollständiges Skelett einschließlich eines fragmentarischen Schädels bekannt, das aus der La-Amarga-Formation in der argentinischen Provinz Neuquén stammt. Damit zählt Amargasaurus zu den am besten bekannten Sauropoden der Unterkreide.[2] Das auffälligste Merkmal war eine Doppelreihe aus langen Wirbelstacheln (gegabelte Dornfortsätze), welche über dem Hals und Rumpf verlief. Ihre Funktion ist jedoch ungeklärt.
Amargasaurus ist ein Vertreter der Dicraeosauridae und war mit den Gattungen des Oberjura Dicraeosaurus und Brachytrachelopan eng verwandt. Die einzige Art ist Amargasaurus cazaui.
Merkmale
Körperbau und Größe
Wie bei allen Sauropoden handelte es sich um einen Pflanzenfresser mit tonnenförmigem Körper und langem Hals und Schwanz. Amargasaurus war ein relativ kleiner Sauropode – Längenschätzungen des einzigen bekannten Exemplars belaufen sich auf 9[3][2] oder 10[4] Meter. Das Gewicht wird von Gerardo Mazzetta und Kollegen (2004) auf 2,6 Tonnen geschätzt.[3] Wie bei anderen Vertretern der Dicraeosauridae war der Hals mit knapp 2,4 Meter Länge[5] im Verhältnis kürzer als bei allen anderen Sauropoden. Laut Leonardo Salgado und José Bonaparte (1991) zeigte Amargasaurus, anders als einige Vertreter der Titanosauria, keinerlei Anpassungen an ein mögliches Aufrichten auf die Hinterbeine, und war daher vermutlich obligat vierbeinig.[6]
Schädel
Lediglich der hintere Bereich des Schädels, vor allem der Hirnschädel und die Schläfenregion, ist überliefert, während Schnauze und Kiefer fehlen.[7] Wahrscheinlich folgte der Schädel jedoch dem für verwandte Gattungen typischen Bauplan mit einer pferdeartig verlängerten und in Draufsicht annähernd rechteckigen Schnauze sowie schmalen, stiftartigen Zahnkronen.[8] Die Nasenöffnungen waren, wie bei verwandten Gattungen, weit nach hinten verschoben.[9] In den ersten Skelettrekonstruktionen wird Amargasaurus mit einem nur leicht nach unten geneigten Schädel dargestellt. Leonardo Salgado (1999) bemerkt jedoch, dass die langen Dornfortsätze der Halswirbel eine solche Schädelhaltung anatomisch unmöglich machen – stattdessen sei der Schädel, anders als bei verwandten Gattungen, stets annähernd vertikal gehalten worden.[10]
Der Schädel lässt sich nur eingeschränkt mit dem naher verwandter Gattungen vergleichen, da die Art Dicraeosaurus hansemanni der bislang einzige andere Vertreter der Dicraeosauridae ist, von dem ebenfalls Schädelmaterial vorliegt. Beide Vertreter zeigen jedoch gemeinsame Schädelmerkmale, die allen anderen bekannten Sauropoden fehlen: So war das paarige Stirnbein (Frontale) miteinander verschmolzen. Die Basipterygoid-Fortsätze, welche den Hirnschädel mit der Oberseite der Mundhöhle verbinden, waren extrem verlängert. Dieses Merkmal war bei Amargasaurus noch stärker ausgeprägt als bei Dicraeosaurus.[11] Des Weiteren war das Supratemporalfenster, ein Schädelfenster der Schläfenregion, stark verkleinert und seitlich ausgerichtet – im Gegensatz zu anderen diapsiden Reptilien, bei denen es stets nach oben ausgerichtet ist.[2] Im Unterschied zu Dicraeosaurus und anderen Sauropoden waren die basalen Tubera – knollenförmige Auswüchse, die sich auf der Unterseite des Hirnschädels befinden – miteinander verschmolzen.[9]
Rumpfskelett und Gliedmaßen
Die Wirbelsäule setzte sich aus 13 Hals-, 9 Rücken- und vermutlich 5 Kreuzwirbeln zusammen, die Anzahl der Schwanzwirbel ist unbekannt. Die Wirbelkörper der Hals- und vorderen Rückenwirbel waren opisthocoel, das heißt, sie waren auf der Vorderseite konvex und auf der Hinterseite konkav. Die übrigen Rückenwirbel dagegen waren amphiplat (beidseitig flach). Wie bei anderen Dicraeosauriden fehlten seitliche Aushöhlungen (Pleurocoele) der Rückenwirbel. Ein einzigartiges Merkmal waren die stark ausgebildeten Querfortsätze (Diapophysen) der vorderen Rückenwirbel, die auf stark ausgeprägte Rippen hinweisen.[2][12]
Das auffälligste Merkmal waren die extrem langen Dornfortsätze der Hals- und Rückenwirbel, die wie bei anderen Vertretern der Dicraeosauridae bis zum Dach des Wirbellochs in zwei Äste gegabelt waren. In ihrer beträchtlichen Länge und ihrer stachelartigen Form unterscheiden sie sich jedoch deutlich von jenen aller anderen Sauropoden. Die Dornfortsätze wiesen einen rundlichen Querschnitt auf und liefen nach oben spitz zu. Sie erreichten im mittleren Bereich des Halses ihre größte Höhe, wo sie am achten Halswirbel 60 cm Länge maßen. Im Bereich des Halses waren sie um etwa 50° nach hinten gebogen, sodass ihre Spitzen den jeweils nachfolgenden Wirbel überragten.[13][4] Die Dornfortsätze der letzten beiden Rückenwirbel, der Kreuzbeinwirbel und der vorderen Schwanzwirbel waren ebenfalls stark verlängert,[4] unterschieden sich jedoch deutlich von denen der vorangegangenen Wirbel: Diese Dornfortsätze waren nicht gegabelt und nach oben hin seitlich erweitert, was ihnen eine paddelartige Form verlieh.[2][12]
Schulter- und Beckengürtel sowie die Vorder- und Hintergliedmaßen glichen denen des verwandten Dicraeosaurus und weisen keine einzigartigen Merkmale auf.[14] Das Becken war relativ breit, da der Abstand zwischen den zu beiden Seiten des Kreuzbeins liegenden Darmbeine (Ilia) vergrößert war.[2] Hände und Füße sind nicht fossil überliefert, waren jedoch vermutlich wie bei allen Sauropoden fünfstrahlig, wobei die Finger der Hand stark zurückgebildet, die Zehen des Fußes jedoch vollständig ausgeprägt waren.
Systematik
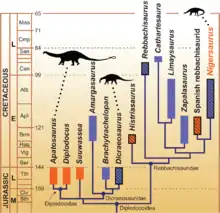
Amargasaurus ist ein Vertreter der Dicraeosauridae, einer von drei Gruppen innerhalb der Diplodocoidea. Der Dicraeosauridae werden derzeit drei Gattungen mit insgesamt vier Arten zugeordnet – neben Amargasaurus cazaui sind dies Dicraeosaurus hansemanni und Dicraeosaurus sattleri aus dem Oberjura Tansanias sowie der erst kürzlich beschriebene Brachytrachelopan mesai aus dem Oberjura Argentiniens. Amargasaurus ist damit der einzige benannte Vertreter aus der Unterkreide, obwohl ein unbenannter Fund aus Brasilien zeigt, dass Vertreter dieser Gruppe noch am Ende der Unterkreide existierten.[1]
Die Verwandtschaftsbeziehungen innerhalb der Dicraeosauridae sind relativ unumstritten. So kommen drei jüngere Analysen aus den Jahren 2005,[16] 2007[15] und 2011[17] übereinstimmend zu dem Ergebnis, dass Dicraeosaurus und Brachytrachelopan näher miteinander verwandt sind als mit Amargasaurus – somit bildet Amargasaurus das Schwestertaxon einer von Brachytrachelopan und Dicraeosaurus gebildeten Klade. Zu einem abweichenden Ergebnis kommt eine Studie aus dem Jahr 1999,[10] welche nahelegt, dass Dicraeosaurus paraphyletisch ist und dass Dicraeosaurus sattleri tatsächlich eine zweite Art von Amargasaurus darstellt.[17][18]
Fund, Forschungsgeschichte und Namensgebung
Das einzige Skelett wurde im Februar 1984 von Guillermo Rougier während einer Expedition unter der Leitung von José Bonaparte geborgen. Diese Expedition war die achte in Patagonien durchgeführte paläontologische Expedition eines Projekts über die jurassische und kreidezeitliche terrestrische Wirbeltierfauna Südamerikas, das bereits 1976 startete.[7] Dieselbe Expedition entdeckte außerdem das Skelett des Theropoden Carnotaurus.[19]
Der Fundort befindet sich im Wadi La Amarga im Departamento Picún Leufú in der argentinischen Provinz Neuquén. Das Skelett (Holotyp, Exemplarnummer MACN-N 15) ist größtenteils im ursprünglichen anatomischen Verbund erhalten: So artikulierte die aus 22 Wirbeln bestehende Rumpf- und Halswirbelsäule mit dem Schädel und mit dem Kreuzbein. Vom Schädel ist lediglich die Schläfenregion sowie der Hirnschädel überliefert, während das Kreuzbein zwar vollständig, aber noch vor der Zuschüttung des Kadavers mit Sedimenten zum Teil erodiert ist. Von den Schwanzwirbeln sind lediglich drei vordere, drei mittlere, ein hinterer sowie Fragmente weiterer Wirbelkörper erhalten. Vom Schultergürtel ist das Schulterblatt (Scapula) mit Rabenbein (Coracoid) vorhanden, vom Becken lediglich das Darmbein (Ilium). Die Gliedmaßen sind ebenfalls nicht vollständig bekannt, so fehlt die Hand und – bis auf zwei Mittelfußknochen – der Fuß. Das Skelett wird in der Sammlung des Museo Argentino de Ciencias Naturales Bernardino Rivadavia in Buenos Aires aufbewahrt.[7]
Die erste, jedoch inoffizielle Erwähnung des neuen Dinosauriers veröffentlichte Bonaparte 1984 in dem italienischen Buch Sulle Orme dei Dinosauria. Hier bezeichnete Bonaparte die Art als Amargasaurus groeberi, obwohl der Artname in der 1991 erschienenen, offiziellen Erstbeschreibung in Amargasaurus cazaui geändert wurde.[20] Die Erstbeschreibung erfolgte durch Leonardo Salgado und José Bonaparte in spanischer Sprache in der Fachzeitschrift Ameghiniana.[21] Ein Jahr später folgte eine weitere, auf den Schädel fokussierte Beschreibung.[22]
Der Name Amargasaurus (Amarga; gr. sauros – „Echse“) weist auf den Wadi La Amarga, in welchem das Skelett gefunden wurde. Der zweite Teil des Artnamens, cazaui, ehrt den bei der argentinischen Ölgesellschaft YPF Sociedad Anónima angestellten Wissenschaftler Luis B. Cazau, der auf die paläontologische Bedeutung der La-Amarga-Formation aufmerksam gemacht hat.[7]
Paläohabitat
Amargasaurus stammt aus der La-Amarga-Formation, einer zum Neuquén-Becken gehörenden Gesteinsabfolge, die auf das Barremium bis Untere Aptium datiert wird. Die Fossilien wurden, ebenso wie die meisten anderen Wirbeltierfunde der La-Amarga-Formation, im untersten der drei Schichtglieder (Subformationen) entdeckt, dem Puesto-Antigual-Member. Dieses Schichtglied weist eine Mächtigkeit von etwa 29 Metern auf und besteht aus Sandsteinen, die innerhalb eines Zopfstromsystems unter hochenergetischen Strömungsbedingungen zur Ablagerung kamen.[23] Die Fossilien von Amargasaurus selbst wurden aus einer sandigen Konglomerat-Lage geborgen.[7] Andere Dinosaurierfunde der Formation schließen Pflanzenfresser wie den ursprünglichen Diplodocoidea Zapalasaurus, den Titanosaurier Amargatitanis, Überreste von ursprünglichen Vertretern der Titanosauriformes sowie von Rebbachisauriden sowie den bislang einzigen Fund eines Stegosauriers in Südamerika mit ein. Fleischfressende Dinosaurier waren mit dem kleinwüchsigen Ceratosaurier Ligabueino vertreten, außerdem weisen Zahnfunde auf die Präsenz eines großen Vertreters der Tetanurae. Neben Dinosauriern ist mit Amargasuchus ein Krokodilverwandter nachgewiesen worden – der Oberkiefer dieses Tieres fand sich zusammen mit den Knochen des Amargasaurus-Skeletts.[24] Wissenschaftlich besonders bedeutsam ist Vincelestes neuquenianus aus der Gruppe der Cladotheria, das bislang einzige aus der Unterkreide Südamerikas beschriebene Säugetier.[25]
Paläobiologie
Funktion der verlängerten Dornfortsätze
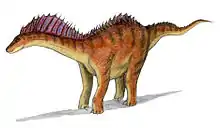
Sowohl das Erscheinungsbild der Dornfortsätze der Hals- und Rückenwirbel als auch ihre Funktion sind ungeklärt. Vermutlich ragten diese stachelartigen Auswüchse aus dem Hals heraus und waren somit von außen sichtbar. Ihre im oberen Abschnitt raue Oberfläche deutet darauf hin, dass die Dornfortsätze beim lebenden Tier möglicherweise mit Horn überzogen waren.[26] Unbekannt ist, ob sie getrennt voneinander standen oder durch ein Segel aus Haut miteinander verbunden waren.[2]
Leonardo Salgado und José Bonaparte (1991) vermuten, dass es sich um Anpassungen zur Verteidigung gegen Prädatoren handelte, worauf die nach oben spitz zulaufende Form der Stacheln hinweise. Andere mögliche Funktionen könnten laut diesen Forschern in der Zurschaustellung gegenüber Artgenossen bestehen, so für die Brautwerbung oder zur Einschüchterung von Rivalen.[6] Jack Bailey (1997) weist darauf hin, dass die stachelartigen Dornfortsätze Ähnlichkeiten mit denen der segeltragenden Pelycosaurier wie Dimetrodon zeigen, und sieht dies als Hinweis darauf, dass auch Amargasaurus ein solches Segel besessen hatte. Dieses Segel könnte laut Bailey der Zurschaustellung gedient haben. Anders als bei den Pelycosauriern waren die Dornfortsätze bei Amargasaurus allerdings gegabelt und bildeten somit eine Doppelreihe. Da der Abstand zwischen beiden Reihen lediglich 3 bis 7 cm betrug, hält es Bailey für unwahrscheinlich, dass zwei separate Segel existierten – stattdessen hätten die Reihen ein Gerüst gebildet, das von einer einzelnen Haut vollständig umspannt wurde. Außerdem weist Bailey darauf hin, dass die Dornfortsätze ab dem vorletzten Rückenwirbel deutlich anders aufgebaut waren als diejenigen der vorangegangenen Wirbel. Laut dem Forscher gleichen diese paddelförmigen Fortsätze denen heutiger höckertragender Huftiere wie den Bisons, was darauf hinweisen würde, dass das Segel im hinteren Rumpfbereich in einen fleischigen Höcker überging.[13]
Laufgeschwindigkeit
Leonardo Salgado und José Bonaparte (1991) vermuten, dass sich Amargasaurus nur langsam fortbewegte, da die Unterarme und Unterschenkel proportional kurz waren, ähnlich wie bei anderen, nicht auf schnelles Laufen ausgelegten Tieren.[27] Gerardo Mazzetta und Richard Fariña (1999) dagegen kommen zu dem Ergebnis, dass Amargasaurus zum schnellen Laufen fähig war: Diese Forscher argumentieren, dass die Beinknochen beim Laufen besonders stark durch Biegemomente beansprucht werden und dass ihre Widerstandsfähigkeit gegen Biegemomente ein limitierender Faktor für die Höchstgeschwindigkeit eines Tieres sei. Die Forscher erklären, dass die Beinknochen von Amargasaurus sogar kräftiger seien als jene des heutigen Breitmaulnashorns, eine an das Galoppieren angepassten Spezies.[28]
Belege
Hauptliteratur
- Jack Bowman Bailey: Neural spine elongation in dinosaurs; sailbacks or buffalo-backs? In: Journal of Paleontology. Bd. 71, Nr. 71, 1997, ISSN 0022-3360, S. 1124–1146, JSTOR 1306608.
- Fernando E. Novas: The age of dinosaurs in South America. Indiana University Press, Bloomington IN 2009, ISBN 978-0-253-35289-7.
- Leonardo Salgado, José F. Bonaparte: Un nuevo sauropodo Dicraeosauridae, Amargasaurus cazaui gen. et sp. nov., de la Formacion La Amarga, Neocomiano de la Provincia del Neuquén, Argentina. In: Ameghiniana. Bd. 28, Nr. 3/4, 1991, S. 333–346.
- Leonardo Salgado, Jorge O. Calvo: Cranial osteology of Amargasaurus cazaui Salgado and Bonaparte (Sauropoda, Dicraeosauridae) from the Neocomian of Patagonia. In: Ameghiniana. Bd. 29, Nr. 4, 1992, S. 337–346.
Ergänzende Literatur
- Sebastián Apesteguía: The sauropod diversity of the La Amarga Formation (Barremian), Neuquén (Argentina). In: Gondwana Research. Bd. 12, Nr. 4, 2007, ISSN 1342-937X, S. 533–546, doi:10.1016/j.gr.2007.04.007.
- José F. Bonaparte, Fernando E. Novas, Rodolfo A. Coria: Carnotaurus sastrei Bonaparte, the horned, lightly built carnosaur from the Middle Cretaceous of Patagonia (= Contributions in Science. Nr. 416, ISSN 0459-8113). Natural History Museum of Los Angeles County, Los Angeles CA 1990, Digitalisat (PDF; 4,99 MB).
- Luis M. Chiappe: A new trematochampsid crocodile from the Early Cretaceous of north-western Patagonia, Argentina and its palaeobiogeographical and phylogenetic implications. In: Cretaceous Research. Bd. 9, Nr. 4, 1988, ISSN 0195-6671, S. 379–389, doi:10.1016/0195-6671(88)90009-2.
- Donald F. Glut: Dinosaurs. The Encyclopedia. McFarland & Company, Jefferson NC u. a. 1997, ISBN 0-89950-917-7.
- Héctor A. Leanza, Sebastián Apesteguía, Fernando E. Novas, Marcelo S. de la Fuente: Cretaceous terrestrial beds from the Neuquén Basin (Argentina) and their tetrapod assemblages. In: Cretaceous Research. Bd. 25, Nr. 1, 2004, S. 61–87, doi:10.1016/j.cretres.2003.10.005.
- Gerardo V. Mazzetta, Richard A. Fariña: XIV jornadas Argentinas de paleontologia de vertebrados. Estimacion de la capacidad atlética de Amargasaurus cazaui Salgado y Bonaparte, 1991, y Carnotaurus sastrei Bonaparte, 1985 (Saurischia, Sauropoda-Theropoda). In: Ameghiniana. Bd. 36, Nr. 1, 1999, S. 105–106.
- Gerardo V. Mazzetta, Per Christiansen, Richard A. Fariña: Giants and Bizarres: Body Size of Some Southern South American Cretaceous Dinosaurs. In: Historical Biology. Bd. 16, Nr. 2/4, 2004, ISSN 0891-2963, S. 71–83, doi:10.1080/08912960410001715132, Digitalisat (PDF; 5747,66 kB).
- Oliver W. M. Rauhut, Kristian Remes, Regina Fechner, Gerardo Cladera, Pablo Puerta: Discovery of a short-necked sauropod dinosaur from the Late Jurassic period of Patagonia. In: Nature. Bd. 435, Nr. 7042, 2005, S. 670–672, doi:10.1038/nature03623.
- Leonardo Salgado: The macroevolution of the Diplodocimorpha (Dinosauria; Sauropoda): a developmental model. In: Ameghiniana. Bd. 36, Nr. 2, 1999, S. 203–216.
- Daniela Schwarz, Eberhard Frey, Christian A. Meyer: Pneumaticity and soft-tissue reconstructions in the neck of diplodocid and dicraeosaurid sauropods. In: Acta Palaeontologica Polonica. Bd. 52, Nr. 1, 2007, ISSN 0567-7920, S. 167–188, online.
- Philip Senter: Necks for sex: sexual selection as an explanation for sauropod dinosaur neck elongation. In: Journal of Zoology. Bd. 271, Nr. 1, 2007, ISSN 0952-8369, S. 45–53, doi:10.1111/j.1469-7998.2006.00197.x.
- Paul C. Sereno, Jeffrey A. Wilson, Lawrence M. Witmer, John A. Whitlock, Abdoulaye Maga, Oumarou Ide, Timothy A. Rowe: Structural Extremes in a Cretaceous Dinosaur. In: PLoS ONE. Bd. 2, Nr. 11, 2007, e1230, doi:10.1371/journal.pone.0001230.
- Michael P. Taylor, Darren Naish: The phylogenetic taxonomy of Diplodocoidea (Dinosauria: Sauropoda). In: PaleoBios. Bd. 25, Nr. 2, 2005, ISSN 0031-0298, S. 1–7.
- Paul Upchurch, Paul M. Barrett, Peter Dodson: Sauropoda. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 259–324.
- John A. Whitlock: A phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda). In: Zoological Journal of the Linnean Society. Bd. 161, Nr. 4, 2011, ISSN 0024-4082, S. 872–915, doi:10.1111/j.1096-3642.2010.00665.x.
- Jeffrey A. Wilson: Overview of Sauropod Phylogeny and Evolution. In: Kristina Curry Rogers, Jeffrey A. Wilson (Hrsg.): The Sauropods. Evolution and Paleobiology. University of California Press, Berkeley CA u. a. 2005, ISBN 0-520-24623-3, S. 15–49.
Einzelnachweise
- Novas 2009, S. 151–152
- Novas 2009, S. 172–174
- Mazzetta et al. 2004, S. 4
- Upchurch et al. 2004, S. 304
- Senter 2007, S. 5–6
- Salgado und Bonaparte 1991, S. 344–346
- Salgado und Bonaparte 1991, S. 333–336
- Wilson 2005, S. 36–37
- Salgado und Calvo 1992, S. 341, 345–346
- Salgado 1999, S. 213–214
- Salgado und Bonaparte 1991, S. 342
- Salgado und Bonaparte 1991, S. 338–340
- Bailey 1997, S. 1124, 1139
- Salgado und Bonaparte 1991, S. 335
- Sereno 2007, S. 5
- Rauhut et al. 2005
- Whitlock 2011, S. 17
- Taylor und Naish 2005, S. 3
- Bonaparte et al. 1990, S. 2
- Glut 1997, S. 123
- Salgado und Bonaparte 1991
- Salgado und Calvo 1992
- Leanza 2004 et al., S. 65–66
- Chiappe 1988, S. 381
- Apesteguía 2007, S. 533–534, 540
- Schwarz 2007, S. 174, 180
- Salgado und Bonaparte 1991, S. 344
- Mazzetta und Fariña 1999