Altingia
Altingia ist eine Pflanzengattung in der Familie der Altingiaceae. Sie umfasst ungefähr zehn Arten von immergrünen Bäumen, die in Südchina, im östlichen Himalaya und in Südostasien verbreitet sind. Die bekannteste Art, Altingia excelsa, liefert unter dem Handelsnamen „Rasamala“ ein wertvolles Nutzholz.
| Altingia | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Altingia | ||||||||||||
| Noronha |
Beschreibung
Vegetative Merkmale
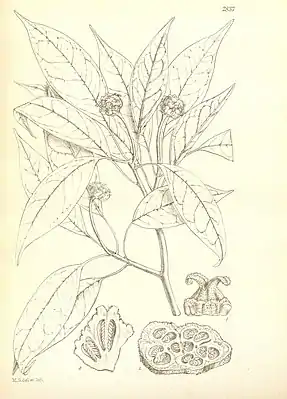
Die Altingia-Arten sind immergrüne Bäume. Die einzelnen Arten unterscheiden sich in den erreichbaren Dimensionen. Während Altingia excelsa Wuchshöhen von bis zu 50 m – in Einzelfällen sogar 60 m – erreichen kann, werden andere Arten, beispielsweise Altingia cambodiana, kaum höher als 10 m. Die Sprossachsen enthalten Sekretkanäle. Die schuppenbedeckten Knospen sind schmal eiförmig. Die schraubig angeordneten Laubblätter sind bis zu 4 cm lang gestielt. Die meist paarweise vorhandenen, sehr schmalen Nebenblätter sind bis zu 6 mm lang. Sie sind entweder hinfällig und hinterlassen kleine Narben oder sind mit dem Blattstiel verwachsen und bleiben länger erhalten. Die einfachen und ungeteilten, ledrigen, zweifarbigen Blattspreiten haben eine lanzettliche bis eiförmige oder verkehrteiförmige Gestalt. Sie sind 4–15(–17) cm lang und 1,5–7 cm breit. Die fiedrige Blattnervatur ist kamptodrom, das heißt, die bogig verlaufenden Seitennerven verästeln sich, bevor sie den Spreitenrand erreichen. Die Blättchen sind am Rand drüsig gekerbt-gesägt oder ganzrandig.
Das Holz von Altingia excelsa hat keine erkennbaren oder nur undeutliche Zuwachszonengrenzen. Das Kernholz ist braun bis rot gefärbt und ohne Farbstreifen, das Splintholz farblich davon deutlich abgesetzt. Die Rohdichte beträgt 0,6–0,85 kg/dm3.[1]
Blütenstände
Die Geschlechtsverteilung der Blüten ist einhäusig getrenntgeschlechtig (monözisch). Die männlichen und weiblichen Blütenstände sind gestielte Köpfchen, die anfangs von vier Hochblättern umhüllt sind. Die vielblütigen männlichen Köpfchen sind kugelig bis kurz zylindrisch. Zwischen den dicht stehenden Blüten befinden sich einige winzige Deckblätter. Diese Köpfchen sind zu traubigen bis rispigen Gesamtblütenständen angeordnet, die an den Enden der Zweige stehen oder nur wenig darunter. Die lang gestielten weiblichen Köpfchen, bestehend aus 5–30 miteinander verwachsenen Blüten, stehen einzeln oder zu Trauben angeordnet in der Nähe der Zweigenden oder im unteren Teil von überwiegend männliche Köpfchen enthaltenden Gesamtblütenständen.
Blüten
Die kleinen, eingeschlechtigen Blüten besitzen keine Blütenhülle. Die männlichen Blüten bestehen ausschließlich aus vier bis zehn freien Staubblättern mit sehr kurzen und dicken oder fehlenden Staubfäden. Die basifixen, also an ihrem Grund dem Staubfaden angehefteten Staubbeutel haben eine verkehrteiförmige bis eiförmige Form und sind an ihrem oberen Ende gestutzt. Die beiden Theken bestehen aus jeweils zwei Pollensäcken und öffnen sich mit einem Schlitz der Länge nach. In den weiblichen Blüten fehlen Staubblätter, aber es sind manchmal nadelähnliche Staminodien vorhanden. Den rudimentären Diskus repräsentiert eine wechselnde Anzahl von kleinen Lappen. Der halbunterständige bis unterständige Fruchtknoten besteht aus zwei miteinander verwachsenen, nur an der Spitze freien Fruchtblättern. Die beiden pfriemlichen Griffel sind oft stark nach außen zurückgekrümmt. Sie besitzen an ihrer Oberseite eine am Griffel herablaufende, papillöse Narbe. Jedes der beiden Fruchtknotenfächer enthält (12–)28–50 Samenanlagen, die in vier Reihen an der zentralwinkelständigen Plazenta auf der Scheidewand eingefügt sind.
Früchte und Samen
Die Früchte eines Fruchtstands sind miteinander zu einem kugeligen bis halbkugeligen Köpfchen verwachsen, in dem die gut entwickelten Früchte mit abortierten vermischt sind. Die einzelnen Früchte sind holzige, verkehrteiförmige bis verkehrt-pyramidale Kapselfrüchte, die von einem Ring von vergrößerten, verhärteten Diskuslappen umgeben sind. Die Griffel beziehungsweise der größte Teil davon und die Staminodien sind im Fruchtzustand nicht mehr vorhanden. Die zweifächerigen Kapseln öffnen sich fachspaltig mit zwei zweiteiligen Klappen und enthalten in jedem Fach zahlreiche Samen. Die oberen Samen sind steril und ungeflügelt. Ein einzelner oder wenige der unteren Samen sind fruchtbar. Diese sind abgeflacht und den Rand entlang oder nur an der Spitze schmal geflügelt. Die Samenschale ist dick und hart. Es ist nur wenig Endosperm vorhanden.
Chromosomen
Beide bisher untersuchten Arten stimmen bezüglich der Chromosomenzahl miteinander überein. Sowohl Altingia excelsa als auch Altingia yunnanensis haben einen diploiden Chromosomensatz mit 2n = 32.[2]
Verbreitung und Lebensraum
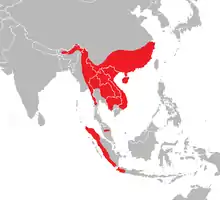
Das Hauptverbreitungsgebiet der Gattung Altingia reicht von der südlichen Abdachung des östlichen Himalaya (Bhutan, Nordost-Indien, Südost-Tibet) bis ins südliche Myanmar (Mergui) und von den südchinesischen Provinzen Zhejiang, Hunan, Guizhou und Yunnan bis ins nördliche und östliche Thailand, nach Kambodscha und ins südliche Vietnam. Altingia excelsa besitzt darüber hinaus auch noch kleinere Teilareale in Bergregionen im westlichen Malaysia (Pahang) sowie auf Sumatra und im Westen von Java. In China kommen acht Arten vor, vier davon sind dort endemisch.
Die Arten der Gattung Altingia wachsen insbesondere in feuchten immergrünen Wäldern und in Bergwäldern. Die Vorkommen liegen in der Regel oberhalb von 400 m Seehöhe. Altingia excelsa beispielsweise kommt in den unteren Lagen des östlichen Himalaya bis auf 1830 m hinauf vor.[3] Manche Arten werden für überdurchschnittlich feuchte Standorte angegeben, etwa Altingia cambodiana für feuchte Böden in der Umgebung von Wasserfällen.[4] Altingia siamensis (= A. takhtajanensis) ist charakteristisch für felsige Flussufer und kann auf granitischem Untergrund in engen Gräben Reinbestände bilden. Diese Art kommt im Süden Vietnams auch in relativ tiefen Lagen bis auf etwa 200 m Seehöhe hinab vor.[5]
Blütenbiologie
Der Bau der Blüten ohne Schauapparat, mit großen Narben und Staubbeuteln, sowie staubförmigem Pollen macht es sehr wahrscheinlich, dass die Altingia-Arten hauptsächlich durch den Wind bestäubt werden (Anemophilie).[6] Die Gattung ist regelmäßig im Pollenniederschlag in den entsprechenden Regionen im tropischen Asien vertreten.[7]
Taxonomie und Systematik
Die Erstbeschreibung von Altingia erfolgte 1790 durch den spanischen Botaniker Francisco Noroña in einer posthumen Publikation.[8] Er war der Typusart Altingia excelsa bei seinem Aufenthalt auf der Insel Java (1786–1787) begegnet. Sedgwickia Griff. ist ein Synonym.
Die Gattung Altingia wird traditionell meist zur Familie der Zaubernussgewächse (Hamamelidaceae) gestellt, etwa zusammen mit Semiliquidambar und den Amberbäumen (Liquidambar) in eine eigene Unterfamilie Altingioideae.[9] Molekularbiologische Untersuchungen[10] haben ergeben, dass die derart weit gefassten Zaubernussgewächse nicht monophyletisch sind. Seitdem hat sich die Abtrennung der drei genannten Gattungen in eine eigene Familie Altingiaceae durchgesetzt. Neuere Studien[11][12] haben die Altingiaceae als Teil einer aus Baum- und Straucharten bestehenden Klade („woody clade“) innerhalb der Steinbrechartigen (Saxifragales) gezeigt. Sie sind hier das Schwestertaxon einer Klade mit den drei übrigen Familien Zaubernussgewächse, Kuchenbaumgewächse (Cercidiphyllaceae) und Daphniphyllaceae.
Zu den Verwandtschaftsverhältnissen innerhalb der Altingiaceae gibt es widersprüchliche Ergebnisse. Während eine kladistische Analyse auf der Grundlage von morphologischen Merkmalen[13] sowohl Altingia als auch Liquidambar mit guter statistischer Absicherung als monophyletisch bestätigt hat, haben phylogenetische Untersuchungen, die auf molekularbiologischen Daten basierten,[14][15] alle drei Gattungen der Familie als para- bzw. polyphyletisch gezeigt. Das nachfolgende Kladogramm präsentiert ein Beispiel für die Verwandtschaftsbeziehungen, wie sie sich mit molekularbiologischen Methoden darstellen:[14]
| |||||||||||||||||||||||||||||||||||||||||||
Etymologie
Die Gattung ist zu Ehren von Willem Arnold Alting (1724–1800) benannt, dem Generalgouverneur von Niederländisch-Indien zur Zeit, als der Erstbeschreiber Francisco Noroña Java besuchte.[16]
Arten
Die Gattung Altingia umfasst etwa zehn – je nach Autor fünf bis zwölf – Arten. Die Gattung bedarf einer taxonomischen Revision.[17]
| Wissenschaftlicher Name | Verbreitung |
|---|---|
| Altingia cambodiana Lecomte | Kambodscha (Elefantenberge) |
| Altingia chinensis (Champ. ex Benth.) Oliv. ex Hance | in Süd-China weit verbreitet, Vietnam |
| Altingia excelsa Noronha | Bhutan, Assam, China (Südost-Tibet, Yunnan), Myanmar, Malaiische Halbinsel, Sumatra, West-Java |
| Altingia gracilipes Hemsl. | Südost-China (Zhejiang bis Guangdong, Hainan) |
| Altingia multinervis W.C.Cheng | Süd-China (Guizhou) |
| Altingia obovata Merr. & Chun | Süd-China (Hainan) |
| Altingia poilanei Tardieu | Nord-Vietnam (Lào Cai) |
| Altingia siamensis Craib | Süd-China (Yunnan, Guangdong), Vietnam, Laos, Kambodscha, Thailand |
| Altingia tenuifolia Chun ex Hung T.Chang | Süd-China (Guizhou, Jiangxi) |
| Altingia yunnanensis Rehder & E.H.Wilson | Süd-China (Yunnan), Vietnam (unsicher) |
Quellen
- Endress P. K. 1993: Hamamelidaceae. In: Kubitzki K., Rohwer J. G., Bittrich V. (Hrsg.): The families and genera of Vascular Plants. Vol. II: Flowering Plants: Dicotyledons: Magnoliid, Hamamelid and Caryophyllid families. Springer, Berlin/ Heidelberg/ New York, ISBN 3-540-55509-9, S. 322–331.
- Tardieu-Blot M.-L. 1965: Hamamelidaceae. In: Flore du Cambodge, du Laos et du Vietnam. Fasc. 4. Muséum National d'Histoire Naturelle, Paris, S. 75–116.
- Vink W. 1957: Hamamelidaceae. In: Flora Malesiana. Ser. I, Vol. 5(3). Botanic Gardens of Indonesia, Bogor, Rijksherbarium, Leyden, S. 363–379. – Altingia – Online
- Zhang Zhiyun, Zhang Hongda & Endress P. K. 2003: Hamamelidaceae. In: Flora of China. Vol. 9. Science Press, Beijing, Missouri Botanical Garden Press, St. Louis, ISBN 1-930723-14-8, S. 18–42. – Altingia – Online
Einzelnachweise
- Richter H. G., Dallwitz M. J.: Altingia excelsa. In: Handelshölzer. Abgerufen am 16. Januar 2013.
- Goldblatt P., & Johnson D. E. (Hrsg.): Altingia. In: Tropicos.org: Index to Plant Chromosome Numbers (IPCN). Missouri Botanical Garden, St. Louis, abgerufen am 16. Januar 2013.
- Long D. G. 1987: Family 65. Hamamelidaceae. In: Flora of Bhutan. Vol. 1(3). Royal Botanic Garden, Edinburgh, ISBN 0-9504270-6-3, S. 468–471.
- Tardieu-Blot M.-L. 1965, S. 95.
- Tardieu-Blot M.-L. 1965, S. 99.
- Endress P. K. 1993, S. 324. – Vorschau bei der Google-Buchsuche
- Corlett R. T. 2004: Flower visitors and pollination in the Oriental (Indomalayan) Region. Biological Reviews 79: 497–532. – doi:10.1017/S1464793103006341
- Noroña F. 1790: Relatio plantarum Javanensium iter factione usque in Bandong recognitarum a Dne. F. Noron(h)a. Verhandelingen van het Bataviaasch Genootschap van Kunsten en Wetenschappen 5: 1. – siehe auch: Altingia. In: Tropicos.org. Missouri Botanical Garden, St. Louis, abgerufen am 16. Januar 2013.
- Endress P. K. 1989: A suprageneric taxonomic classification of the Hamamelidaceae. Taxon 38: 371–376. – JSTOR 1222267
- Chase M. W., Soltis D. E., Olmstead R. G., Morgan D., Les D. H., Mishler B. D., Duvall M. R., Price R. A., Hills H. G., Qui Yin-Long, Kron K. A., Rettig J. H., Conti E., Palmer J. D., Manhart J. R., Sytsma K. J., Michaels H. J., Kress W. J., Karol K. G., Clark W. D., Hedrén M., Gaut B. S., Jansen R. K., Kim Ki-Joong, Wimpee C. F., Smith J. F., Furnier G. R., Strauss S. H., Xiang Qiu-Yun, Plunkett G. M., Soltis P. S., Swensen S. M., Williams S. E., Gadek P. A., Quinn C. J., Eguiarte L. E., Golenberg E., 21 Learn G. H., Graham S. W., Barrett, S. C. H., Dayanandan S., Albert V. A. 1993: Phylogenetics of seed plants: an analysis of nucleotide sequences from the plastid gene rbcL. Annals of the Missouri Botanical Garden 80: 528–580. – Online
- Jian Shuguang, Soltis P. S., Gitzendanner M. A., Moore M. J., Li Ruiqi, Hendry T. A., Qiu Yin-Long, Dhingra A., Bell C. D., Soltis D. E. 2001: Resolving an ancient, rapid radiation in Saxifragales. Systematic Biology 57: 38–57. – doi:10.1080/10635150801888871
- Soltis D. E., Smith S. A., Cellinese N., Wurdack K. J., Tank T. C., Brockington S. F., Refulio-Rodriguez N. F., Walker J. B., Moore M. J., Carlsward B. S., Bell C. D., Latvis M., Crawley S., Black C., Diouf D., Xi Zhenxiang, Rushworth C. A., Gitzendanner M. A., Sytsma K. J., Qiu Yin-Long, Hilu K. W., Davis C. C., Sanderson M. J., Beaman R. S., Olmstead R. G., Judd W. S., Donoghue M. J., Soltis P. S. 2011: Angiosperm phylogeny: 17 genes, 640 taxa. American Journal of Botany 98: 704–730. – doi:10.3732/ajb.1000404
- Ickert-Bond S. M., Pigg K. B., Wen J. 2007: Comparative infructescence morphology in Altingia (Altingiaceae) and discordance between morphological and molecular phylogenies. American Journal of Botany 94: 1094–1115. – doi:10.3732/ajb.94.7.1094
- Shi S., Huang Y., Zhong Y., Du Y., Zhang Q., Chang H., Boufford D. E. 2001: Phylogeny of the Altingiaceae based on cpDNA matK, PY-IGS and nrDNA ITS sequences. Plant Systematics and Evolution 230: 13–24. – doi:10.1007/s006060170002
- Ickert-Bond S. M., Wen J. 2006: Phylogeny and biogeography of Altingiaceae: Evidence from combined analysis of five non-coding chloroplast regions. Molecular Phylogenetics and Evolution 39: 512–528. – doi:10.1016/j.ympev.2005.12.003
- Hayne F. G. 1830: Getreue Darstellung und Beschreibung der in der Arzneykunde gebräuchlichen Gewächse. Vol. 11. Berlin. – Vorschau bei der Google-Buchsuche
- Endress P. K. 1993, S. 330. – Vorschau bei der Google-Buchsuche
Weblinks
- Altingia. In: Germplasm Resources Information Network (GRIN). United States Department of Agriculture (USDA), ARS, National Genetic Resources Program, National Germplasm Resources Laboratory, Beltsville, Maryland, abgerufen am 16. Januar 2013.