Insektenflug
Insekten waren in der Evolution die ersten Tiere, die Flügel entwickelt haben. Die ältesten Belege finden sich im Karbon vor ca. 320 Millionen Jahren.[1] Innerhalb der Wirbellosen sind sie die einzige Klasse mit Flügeln.

Die Flügel der Insekten bestehen aus Chitin, ihre Vorderkanten sind versteift, während der übrige Teil elastisch bleibt. Aufgrund ihrer im Vergleich zu Wirbeltiere geringeren Größe, ist die Schlagfrequenz der Flügel meist wesentlich höher.[2]
Fluginsekten sind die artenreichste Gruppe im gesamten Tierreich. Die meisten Arten haben zwei Paar Flügel.
Flugmuskulatur
Eine direkte Flugmuskulatur haben Libellen und Eintagsfliegen, die meisten Insektenordnungen besitzen dagegen eine indirekte Flugmuskulatur.
Als Flügelschlagfrequenzen für verschiedene Insekten wurden Werte von 0 (Libelle im Segelflug) bis zu 1046 Hz bei kleinen Mücken (Gattung Forcipomyia, Ceratopogonidae)[3] gemessen. Da ein Muskel in dieser Frequenz durch direkte Nervenimpulse nicht angesteuert werden kann – die Latenzzeit und absolute Refraktärzeit des Aktionspotentials eines Nervs die maximal mögliche Frequenz auf etwa 800 bis 1000 Hz limitiert – kontrahieren bei Insekten mit indirekter Flugmuskulatur die Muskeln selbstständig bei Überdehnung. Da sehr kleine Insekten nur mit entsprechend hohen Flügelschlagfrequenzen die für den Flug nötigen Luftkräfte erzeugen können, besitzen sie stets eine indirekte Flugmuskulatur. Die flugfähigen Vertreter des ursprünglicheren, direkten Typs besitzen alle eine gewisse Mindestgröße.[4]
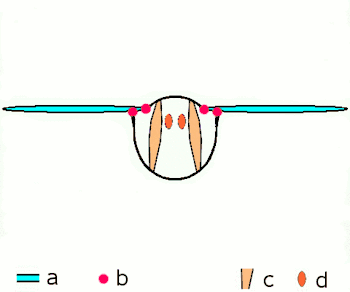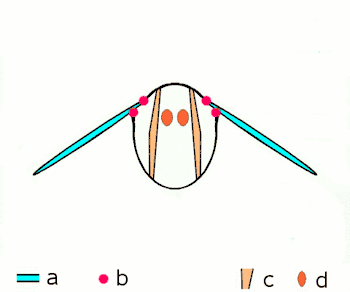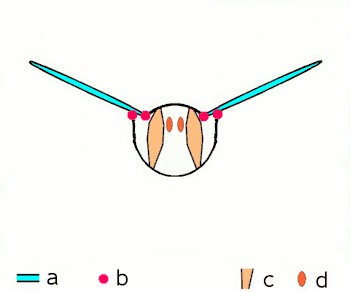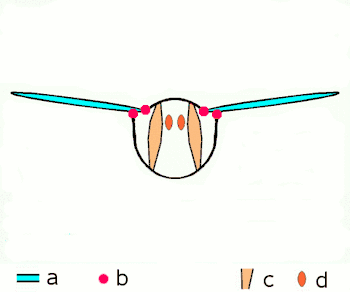
a Flügel
b primäre und sekundäre Flügelgelenke
c dorsoventrale Flugmuskulatur
d Muskulatur parallel zur Körperlängsachse
Indirekte Flugmuskulatur
Die Bezeichnung stammt daher, dass die Muskulatur nicht direkt an den Flügeln ansetzt, sondern die Muskelkraft indirekt über die Skelettelemente des Thorax auf die Flügel übertragen wird. Die für den Flügelabschlag zuständigen Muskeln sind längs des Thorax angeordnet (Longitudinalmuskeln), die für den Flügelaufschlag zuständigen Muskeln ziehen vom „Rücken“ des Thorax zu dessen „Bauch“ (Dorsoventralmuskeln). Die Verformungen des Thoraxskelettes übertragen die Muskelkraft auf die Flügel.
Während des Fluges beobachtet man in einigermaßen regelmäßigen Abständen Aktionspotentiale von Motoneuronen, die in den Flugmuskeln zur Ausschüttung von Calciumionen aus dem sarkoplasmatischen Retikulum führen. Die Calciumkonzentration im Cytoplasma der Muskelzellen (Myoplasma) ist bei diesem speziellen Muskeltyp jedoch nicht der Auslöser der Kontraktionen. Ausgelöst wird die Kontraktion einer Muskelzelle durch ihre Dehnung, die im Flugbetrieb durch ihren Antagonisten erfolgt. Das Vorhandensein einer bestimmten myoplasmatischen Calciumkonzentration ist eine notwendige, jedoch nicht hinreichende Bedingung für die Kontraktion der indirekten Flugmuskeln. Möglicherweise wird beim ruhenden System jedoch die erste Muskelkontraktion durch eine initiale Calciumausschüttung ausgelöst, die durch ein Aktionspotential eines Motoneurons hervorgerufen wird.
Angehalten wird das System durch das Ausbleiben von Aktionspotentialen der Motoneurone, da dann die Calciumkonzentration im Myoplasma durch Ionenrückaufnahme so stark absinkt, dass Kontraktionen nicht mehr erfolgen können.
Die Kontraktionen der Muskelzellen verlaufen anders als bei Muskelzellen üblich, da die Myosinköpfchen durch eine Konformationsänderung zwar umknicken, sich jedoch nicht vom Aktin ablösen um an einem weiteren Aktinabschnitt neu anzusetzen („biologisches Zahnrad“ der Muskeln). Daher ist die Verkürzung des Flugmuskels extrem gering und muss durch eine mechanische Übersetzung auf den Flügel übertragen werden. Diese Übersetzung wird unter anderem durch eine Kette von einarmigen Hebeln erreicht, indem die Kraft vom vorderen Teil des Thoraxrückens, dem Scutum, auf den hinteren Rückenteil (Scutellum) übertragen wird, um von dort über die auf beiden Seiten am Scutellum ansitzenden Tergalhebel wieder nach vorne zu den Flügelgelenken gebracht zu werden.
Schon wegen dieses Mechanismus', bei dem beide Flügel nur gleichzeitig auf- bzw. abgeschlagen werden können, hat die Fliege nicht die Möglichkeit, die Vortriebskräfte, die beim Kurvenflug auf beiden Seiten unterschiedlich sein müssen, durch eine unterschiedliche Schlagfrequenz zu erzeugen. Stattdessen kann sie die Amplitude des Flügelschlages, sowie den Anstellwinkel des Flügels auf beiden Seiten separat ändern. Dies wird durch eine hochkomplizierte Flügelgelenkkonstruktion und eine Reihe von dünnen Stellmuskeln erreicht, die das Gelenk jeweils passend einstellen können.
Die Flügelschlagfrequenz von Insekten mit indirekter Flugmuskulatur wird von den Luft- und Masseträgheitskräften bestimmt, die der Muskelkontraktion entgegenstehen. Die Flugmuskulatur kontrahiert aufgrund der dehnungsausgelösten Kontraktion entweder vollständig, oder gar nicht. Bei Änderungen von Flügelschlagamplitude oder Flügelstellung, die für die Flugsteuerung eingesetzt werden, ändert sich jedoch der Luftwiderstand am Flügel, der der Kontraktionskraft entgegensteht. Dadurch ändert sich dann die Flügelschlagfrequenz des Insektes. Im Experiment kann man an thoraxfixierten Fliegen durch Kürzen oder durch Beschweren des Flügels mit Wachs die Abhängigkeit der Schlagfrequenz von den mechanischen Eigenschaften des gesamten Systems, das man als Resonanzsystem auffassen kann, beobachten.
Die Steuerung der Flugmuskeln kann äußerst primitiv sein und an die Meldung von Bodenkontakt der Tarsen der Insektenbeine gekoppelt sein (wie zum Beispiel bei der Fliege). Zum Start springt die Fliege einfach vom Boden weg, alle Beine melden keinen Bodenkontakt und der Flugmuskel wird daraufhin gestartet.
Bei der Landung meldet ein oder mehrere Beine Bodenkontakt und die Flügelmuskeln werden gestoppt. Befestigt man eine Fliege an ihrem Thorax, ohne dass ihre Beine Kontakt zum Boden haben, schlagen ihre Flügel oft sehr lange.
Die indirekte Flugmuskulatur von Fliegen und anderen Insekten ist eines der Gewebe, die den höchsten Energieumsatz besitzen. Elektronenmikroskopische Aufnahmen zeigen eine extreme Dichte der Mitochondrien, die allen Zellen als „Energiekraftwerke“ dienen. Durch die enorme Abwärmeproduktion beim Flug wird ein effizientes Kühlsystem benötigt. Bienen und wahrscheinlich auch Fliegen verwenden hier – ebenso wie Säugetiere – die sehr effiziente Verdunstungskühlung. Die Insekten pumpen die von den Flugmuskeln im Thorax erhitzte Hämolymphe in den Kopf und würgen einen Flüssigkeitstropfen aus, der am Rüssel austritt und dann verdunstet. Bei Fliegen, die auf der Thoraxoberseite befestigt werden, kann man diesen Tropfen nach einiger Zeit sehr schön beobachten. Die Neigung der Insekten, zu fliegen, geht bei hoher Luftfeuchtigkeit stark zurück.
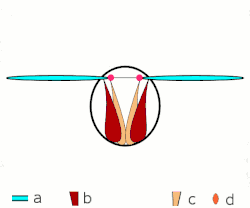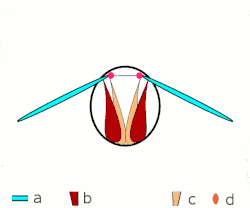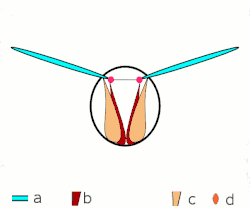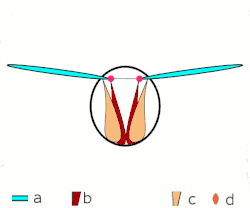
a Flügel
b, c dorsoventrale Flugmuskulatur
d Flügelgelenke
Direkte Flugmuskulatur
Bei ursprünglicheren Insekten, wie beispielsweise Libellen, findet man einen direkten Antrieb (im Gegensatz zu den übrigen Fluginsekten). Die Flügel sind hier direkt mit den antagonistischen Muskeln verbunden. Die Steuerung der Bewegung ist bei dem direkten Ansatz an den Flügeln analog zur Steuerung der Skelettmuskulatur bei Wirbeltieren (z. B. Beuger/Strecker bei Armmuskeln). Die Kontraktionen werden dabei direkt vom Nervensystem über Aktionspotentiale der Motoneurone ausgelöst. Wie bei Laufbewegungen sind im zentralen Nervensystem solcher Insekten bestimmte Nervenverschaltungen vorhanden, die auch Mustergeneratoren genannt werden und das Kontraktionsverhalten der Flugmuskulatur als Aktionspotential-Sequenzen erzeugen. Der Antrieb mit direkter Flugmuskulatur erlaubt eine unabhängige Bewegung der vier Flügel. Libellen sind dadurch extrem wendige Flieger, was den Beutefang in der Luft ermöglicht.
Bedeutung der Viskosität
Abhängig von seiner Größe und Fluggeschwindigkeit ist die Viskosität der Luft für das Insekt von unterschiedlicher Bedeutung. Diese drei Größen können zur Reynoldszahl zusammengefasst werden, die das Verhältnis von Trägheits- zu viskosen Kräften angibt.
Besonders für kleine Insekten sind die viskosen Kräfte dominant, die Luft ist für sie aufgrund ihrer Größe und Fluggeschwindigkeit so zäh wie Wasser (siehe auch Weblinks: „Wenn die Luft klebrig wird“). Sie haben daher auch keine strömungsgünstigen Flügel, sondern z. T. nur pinselartige Widerstandserzeuger wie zum Beispiel die Borsten- oder Fransenflügler.
Größere oder schneller fliegende Insekten besitzen meist durchsichtige Hautflügel. Hiervon kann das hintere Paar zu sogenannten Schwingkölbchen reduziert sein, die lediglich zum Massenausgleich gegen die Flügel rotieren und als Kreiselinstrument genutzt werden (bei den Dipteren, wie den Stechmücken und Fliegen).

Bei Fliegen erfolgt der Flügelabschlag von hinten oben nach vorne unten mit horizontaler Flügelstellung. Dadurch ist der Luftwiderstand in vertikaler Richtung maximal und in horizontaler Richtung minimal. So wird ein hoher Auftrieb und ein minimaler Rücktrieb erzeugt. Beim Flügelaufschlag wird der Flügel von vorne unten nach hinten oben bei vertikaler Flügelstellung bewegt. Daraus resultiert ein minimaler Luftwiderstand in vertikaler Richtung und ein entsprechend geringer Abtrieb, sowie ein maximaler Widerstand in horizontaler Richtung und entsprechend ein maximaler Vortrieb. Diese Aufgabenteilung der antagonistischen Muskelgruppen in Auf- und Vortrieb hat ein ungefähres Kräftegleichgewicht zur Folge, das für die gegenseitige Kontraktionsauslösung der Antagonisten bei der indirekten Flugmuskulatur durch Dehnung von nicht unerheblicher Bedeutung ist.
Aus dem vorderen Flügelpaar kann auch ein verhornter Deckflügel gebildet werden, der als Starrflügel im Flug Auftrieb erzeugt, während das größere, hautige hintere Flügelpaar für den Vortrieb zuständig ist (Beispiel Coleoptera, Käfer).

Aber es gibt auch zahlreiche Arten, bei denen beide Flügelpaare voll entwickelt sind. Der Libellenflügel der Großlibellen (Anisoptera) gilt dabei als Meisterstück der Natur. An jedem der vier Flügel formt sich mit Hilfe der Form der Tracheen und Membranen im Luftstrom ein durch eingelagerte Wirbelstromwalzen entstehendes widerstandsarmes Laminarprofil (also ein Flügelprofil mit hoher Dickenrücklage), das für Segelflug, stationären Schlagflug und zum Schnellflug genutzt werden kann. Trimmtanks, die in den Flügeln eingelagert sind (Pteristigmata) ermöglichen eine zusätzliche Steuerung. Obwohl der Flügel nur 2 Prozent des Gesamtgewichts der Libelle ausmacht, ermöglicht er unvergleichliche Stabilität und hervorragende Flugeigenschaften, wobei er mehr als 50 Prozent der Oberfläche des Tieres ausmacht. Dieses Verhältnis von Tragflächengewicht und Oberflächenanteil zu Gesamtgewicht ist in der gesamten menschlichen Luftfahrt unerreicht, bei der jedoch auch andere Mechanismen zum Fliegen genutzt werden.

Kleinere Insekten nutzen zur Auftriebsgewinnung im Schlagflug meist die Technik des leading edge vortex. Durch den Auf- und Abschlag ihrer scharfkantigen Flügelvorderkante wird entlang dieser Kante ein Wirbel erzeugt, der direkt für den Auftrieb genutzt werden kann, indem der hintere Teil dieser entstehenden Wirbelwalze beim Abschlag den Flügel nach oben saugt, beim Aufschlag den Flügel nach oben drückt. Dies erklärt die hervorragenden Flugeigenschaften von Schmetterlingen (Lepidoptera) von denen einige in ihrem Leben mehrere Interkontinentalflüge von bis zu 4000 km im Schlagflug absolvieren können, wie zum Beispiel der Monarchfalter.
Literatur
- Nachtigall, Werner: Insektenflug. Springer Berlin, 2003, ISBN 354000047X
- Nachtigall, Werner und Rolf Nagel: Im Reich der Tausendstel-Sekunde, Faszination des Insektenflugs. Gerstenberg, 1988, ISBN 3806720436
- Hagemann & Steininger (Hrsg.): Alles was fliegt – in Natur, Technik und Kunst. Kleine Senckenbergreihe Nr. 23, 1996, ISBN 3-7829-1143-1
Weblinks
- „Wenn die Luft klebrig wird“ (PDF) (134 kB)
- Flattern für die Wissenschaft – Bionische Forschung an Insekten (PDF; 305 kB). Von Dörte Ostersehlt In: Unterricht Biologie 332 2008. S. 22 bis 27
- University of Florida Book of Insect Records. Sammlung von gemessenen Fluggeschwindigkeiten verschiedener Insekten.
- "Fotos von Insekten im Flug"
Einzelnachweise
- Carsten Brauckmann, Brigitte Brauckmann, Elke Gröning: The stratigraphical position of the oldest known Pterygota (insecta. Carboniferous, Namurian). In: Annales de la Société Géologique de Belgique. 117 (Hommage à Maurice Streel) Fascicule 1, 1994, S. 47–56.
- Schülerduden Biologie. Bibliographisches Institut & F. A. Brockhaus AG, München 2009. S. 192f.
- C. W. Scherer: Book of Insect Records, Univ. Florida, Kap. 9, 1995
- Konrad Dettner, Werner Peters: Lehrbuch der Entomologie. 2. Auflage. Elsevier, München, ISBN 3-8274-1102-5, S. 249.