Heterocyste
Heterocysten (griechisch ἕτερος héteros „andersartig“, κύστις kýstis „Blase“) sind spezialisierte Zellen in manchen Zellketten (=Filament) bildenden Cyanobakterien (s. Abbildung dort), in denen die enzymatische Fixierung des Luftstickstoffs durch das Enzym Nitrogenase stattfindet (biologische Stickstofffixierung).[1]
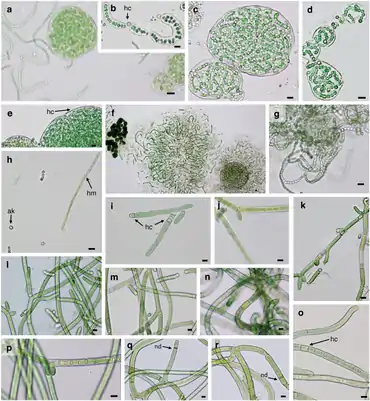
a—f: Nostoc commune, g—h: Nostoc calcicola
i—m: Tolypothrix distorta, n—r: Scytonema hyalinum
Balkenlänge 10 µm
Diese Zellen werden gebildet, sobald gebundener Stickstoff (Nitrat, Ammonium) in der Umgebung fehlt. Etwa jede zehnte vegetative Zelle eines Filaments wird in einem zehn bis 15-stündigen Entwicklungsprogramm irreversibel differenziert. Unter dem Mikroskop fallen diese oft auch etwas größeren Zellen auf durch ihre verdickte Zellwand, die hellgrüne bis fast gelblich-transparente Erscheinung und die bei älteren Heterocysten auffälligen, aus dem Reserve-Polymer Cyanophycin (Arginin-Aspartat-Copolymer) bestehenden Polkörper an den Verbindungsstellen zu den vegetativen Zellen.
Am Ende des Zelldifferenzierungsprogramms der Heterocysten steht die Expression eines zelltypspezifischen Stickstofffixierungssystems (entwicklungsgesteuertes Nif1-System). In vegetativen Zellen mancher Cyanobakterien kann zudem (schon nach circa zwei Stunden) ein eigenes Stickstofffixierungssystem angeschaltet werden, jedoch ist bei diesem die Abwesenheit von gebundenem Stickstoff UND von Sauerstoff in der Umgebung erforderlich (umweltgesteuertes Nif2-System).
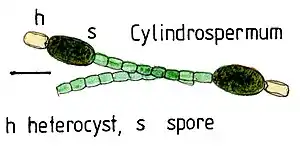
Da alle Nitrogenasen äußerst sauerstoffempfindlich sind, ist der sauerstoffentwickelnde Teil der Photosynthese (Photosystem II) in Heterocysten nicht aktiv. Photosystem I arbeitet jedoch und kann daher – neben der Atmung – im Licht dazu beitragen, den sehr hohen Energiebedarf (ATP) der Nitrogenase zu gewährleisten. Vor von außen eindringendem Sauerstoff schützt die Heterocysten eine extra aufgelagerte Zellwandschicht aus Glycolipid- und Polysaccharidschichten, deren Schichtdicke sich dem Sauerstoffgehalt der Umgebung anpasst (Sauerstoff wird auch bei der Synthese dieser Schicht benötigt). In den umgebenden vegetativen Zellen des Filaments wird durch sauerstoffentwickelnde Photosynthese Zucker erzeugt (wahrscheinlich vor allem Saccharose). Diese werden in die Heterocysten transportiert und dort so weit oxidiert, dass einerseits genügend Elektronen zur Reduktion des Luftstickstoffs zu Ammonium zur Verfügung stehen und andererseits die entstehenden teiloxidierten Kohlenstoffgerüste zum Einbau des Ammoniums zur Verfügung stehen (Endprodukt: Glutamin). Der im Glutamin gebundene Luftstickstoff wird in die benachbarten Zellen transportiert und versorgt so den gesamten Zellfaden mit gebundenem Stickstoff. Der in den Heterocysten ablaufende Stoffwechsel ergänzt die in den vegetativen Zellen ablaufende photosynthetische CO2-Fixierung somit um eine photosynthetische N2-Fixierung. Auf diese Weise wird in Heterocysten bildenden Cyanobakterien aus den „unerschöpflichen“ Grundstoffen H2O, CO2 und N2 unter maximaler Ausnutzung von Lichtenergie Biomasse erzeugt. Es handelt sich hier um die effektivste bekannte Art der Primärproduktion.
Evolution der Heterozyste
Es ist anzunehmen, dass die Entwicklung der gleichzeitigen Photofixierung von Kohlendioxid und Distickstoff zur Primärproduktion in einem Organismus schon sehr früh in der Erdgeschichte „in der Luft lag“: als Reaktion auf den Rückgang anderer ergiebiger organischer und anorganischer Energie-, Kohlenstoff- und Stickstoffquellen, verursacht durch die Stoffwechseltätigkeit konkurrierender Bakterien. Anzeichen von Heterocysten gibt es jedenfalls schon aus Fossilien in Sedimenten, die ein Alter von etwa 2,2 Milliarden Jahre aufweisen. In dieser Zeit war die Atmosphäre praktisch noch frei von Sauerstoff. Dieser Befund unterstützt die Schlussfolgerung, dass die Entwicklung der Heterocysten zuerst auf den Schutz der Nitrogenase vor Sauerstoff zurückgeht, der in den eigenen vegetativen Zellen durch zeitgleiche Photosynthese produziert wird, und der zu hohen lokalen Überkonzentrationen von Sauerstoff in der Mikroumgebung der Zellfilamente führen kann.
Einzelnachweise
- C. Peter Wolk, Anneliese Ernst, Jeff Elhai: Heterocyst Metabolism and Development. In: The Molecular Biology of Cyanobacteria. Advances in Photosynthesis and Respiration. Band 1, 2004, S. 769–823. ISBN 978-0-7923-3222-0.