Epulopiscium fishelsoni
Epulopiscium fishelsoni (Epulopiscium: Gast beim Bankett eines Fisches, fishelsoni nach dem Entdecker) ist ein ungewöhnlich großes grampositives Bakterium. Es lebt symbiotisch im Darm von Doktorfischen, wo es beim Abbau von Nahrung hilft, die der Fisch aufgenommen hat.[1] Die 1985 entdeckte Art galt mit Längen von bis zu 0,6 Millimetern bis zur Entdeckung von Thiomargarita namibiensis 1999 als das größte bekannte Bakterium. Es gibt jedoch daneben auch Zellen, die nur 30 µm groß werden, so dass es zu 2000-fachen Volumenunterschieden zwischen großen und kleinen Vertretern der Art kommt[2].
| Epulopiscium fishelsoni | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Epulopiscium | ||||||||||||
| Montgomery & Pollak 1988 | ||||||||||||
| Wissenschaftlicher Name der Art | ||||||||||||
| Epulopiscium fishelsoni | ||||||||||||
| Montgomery & Pollak 1988 | ||||||||||||
Mittlerweile sind einige ähnliche Zellen auch in anderen Doktorfischen entdeckt worden. Die Artgrenzen sind beim jetzigen Stand der Forschung nicht eindeutig. Daher wird teilweise von Epulopiscium spp. oder Morphotypen gesprochen.[3]
Physiologie
Epulopiscium fishelsoni hat eine einzigartige Anatomie, deren Spezifika meist dazu dienen, klassische Begrenzungen der Größe eines Bakteriums zu überwinden. Die Gründe für die enorme Größe sind unbekannt. Eine mögliche Ursache ist der Schutz gegen bakterienfressende Protisten.
Die Zellwand ist vielfach eingefaltet, um die Oberfläche zu maximieren. Sie verfügt über einen Kortex mit Tubuli, Vesikeln und anderen Strukturen, die normalerweise nur bei Eukaryoten zu finden sind. Möglicherweise dienen diese Strukturen dem intrazellulären Transport. Damit wäre Epulopiscium fishelsoni ein ungewöhnliches Beispiel für konvergente Evolution in Dimensionen von Einzelzellen.
Epulopiscium fishelsoni Typ B lebt im Darmtrakt des Nasendoktorfisches Naso tonganus und wird 200–300 µm lang und 50–60 µm breit. Für diesen Typ wurde beschrieben, dass er hochgradig polyploid ist und bis zu 200.000 Kopien seines Genoms in sich trägt, in Abhängigkeit von der Zellgröße. Es ergab sich dabei ein Verhältnis von einer Kopie des Genoms auf 1,9 µm3 Cytoplasma. Im Vergleich zu Bacillus subtilis, einem normal großen Bakterium mit einem Genom auf 0,7 µm3 Cytoplasma liegt das Genom-Cytoplasma-Verhältnis also in einer ähnlichen Größenordnung.[4] Die DNA von Epulopiscium fishelsoni Typ B fand sich nahe der Zelloberfläche. Die Autoren der Studie spekulieren, dass dadurch eine schnellere Antwort auf äußere, örtliche Reize möglich sei, als wenn die Genome in der Mitte der Zelle lägen. Auch beim Riesenbakterium Thiomargarita namibiensis findet sich die DNA am Rand der Zelle. Hier ist das Stoffwechsel-aktive Cytoplasma jedoch durch eine große zentrale Vakuole, welche 98 % des Zellvolumens einnimmt, auf einen schmalen Rand begrenzt, während bei Epulopiscium fishelsoni das zentrale Cytoplasma anscheinend aktiv ist.[4]
Durch die Zellgröße und die hohe Anzahl an Erbgut-Kopien hat sich Epulopiscium manche Vorteile eukaryotischer Zellen angeeignet.[4]
Vermehrung
Einer der ungewöhnlichsten Aspekte von Epulopiscium fishelsoni ist sein Reproduktionsverhalten. Anders als die meisten Bakterien, die sich asexuell durch Zellteilung vermehren, reproduziert sich Epulopiscium auf eine Weise, die vermutlich aus der Sporulation entstanden ist: meistens zwei, in einem Morphotyp aber auch bis zu zwölf Tochterzellen wachsen in der Mutterzelle heran, bis diese sich auflöst und die neuen Individuen freigibt.[1]
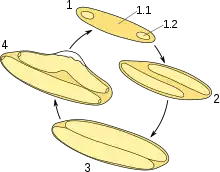
Obwohl Sporulation unter Bakterien nicht ungewöhnlich ist (siehe Bacillus subtilis), ist sie normalerweise eher ein Mittel in Mangel- bzw. Stresssituationen als der übliche Weg der Vermehrung. Auch sind die Tochterzellen üblicherweise Ruhestadien, wohingegen die freigesetzten Zellen von Epulopiscium bereits aktiv sind. Ähnliche Reproduktionswege gehen andere symbiotische Darmbakterien wie z. B. Metabacterium polyspora, die Epulopiscium phylogenetisch nahestehen. Möglicherweise ermöglicht Sporulation den Schutz der Tochterindividuen vor den rauen Bedingungen des Verdauungssystems.
Bei E. fishelsoni-ähnlichen Endosymbionten wurde auch Vermehrung durch Teilung oder durch Mischformen gefunden.[3]
Ökologie
Aus den Eingeweiden der meisten Doktorfisch-Arten konnten unterschiedliche Stämme von Epulopiscium isoliert werden, eine Reinkultur gelang bisher aber nicht.
Die Beziehung der Doktorfische und Bakterien zueinander wird als Mutualismus eingestuft. Der genaue Charakter der Beziehung ist noch unbekannt, es wird jedoch als sicher angenommen, dass Epulopiscium fishelsoni den Fischen dabei hilft, Nährstoffe aus gefressenen Algen und aus Detritus zu gewinnen.
Der Tageszyklus von Epulopiscium fishelsoni verläuft synchronisiert mit den täglichen Aktivitäten seines Wirtes. Über Tag, während der Doktorfisch Algen aufnimmt, wandern die rundlichen, kompakten Nukleoide an die Enden der Zelle und beginnen sich zu strecken. Wenn sie sich am späten Nachmittag / frühen Abend auf rund 50 bis 75 % der Länge der Zelle ausgedehnt haben, beginnt die Sporulation.
Geschichte und Systematik
1985 wurde vom israelischen Wissenschaftler Lev Fishelson von der Universität Tel Aviv und Kollegen die Entdeckung eines großen, zigarrenförmigen, zunächst unbenannten Einzellers veröffentlicht, den sie im Darm von Goldtupfen-Doktorfischen (Acanthurus nigrofuscus) aus dem Roten Meer fanden.[5] Er konnte eine Größe von bis zu 0,5 mm erreichen, rund die dreifache Länge eines Pantoffeltierchens, über 85 % der Zellen waren jedoch kleiner als 0,2 mm. Sie hielten ihn aufgrund seiner Größe für einen Protisten, den sie jedoch keiner bekannten Protistengruppe zuordnen konnten. Weitere Gründe für die Einordnung als Protist waren haarähnliche Strukturen an der Zelloberfläche und eine Struktur im Zellinneren, die für einen Zellkern gehalten wurde. Der Einzeller kam mit einer Dichte von 20.000 bis 100,000 Zellen pro Milliliter im Darm dieses pflanzenfressenden Doktorfisches vor. Die Autoren hielten ihn für einen symbiotisch mit dem Fisch lebenden Organismus. Für diese Annahme sprach, dass sie ihn in jedem von mehreren hundert Exemplaren von Acanthurus nigrofuscus, finden konnten. Auch in Acanthurus sohal wurde er entdeckt, nicht jedoch in mehreren anderen verwandten Arten.[5]
Die Benennung in Epulopiscium fishelsoni erfolgte in einer Veröffentlichung 1988.[2][6]
Eine rRNA-Analyse durch Pace 1993 ergab, dass es sich um ein Bakterium handelt. Zwar konnte es in der Folge der Ordnung der Clostridiales zugeordnet werden, seine genaue systematische Position darin ist allerdings noch ungeklärt. Der Gattungsname verweist auf die symbiotische Lebensweise und bedeutet so viel wie „Gast beim Bankett eines Fisches“; das Artepitheton ehrt den Entdecker Lev Fishelson.
Bei einer Untersuchung der Darmflora von Riff-Fischen im Pazifik wurden in Doktorfischen eine Reihe ähnlicher Einzeller gefunden. Sie wurden aufgrund ihrer Größe und ihres Reproduktionsverhaltens klassifiziert. Große Zellen ähnlich Epulopiscium fishelsoni wurden zum Beispiel wiederum im Goldtupfen-Doktorfisch A. nigrofuscus und im Blaustreifen-Doktorfisch Acanthurus lineatus gefunden. Sie werden als A-Morphotyp bezeichnet.[3] Mittlerweile sind über zehn Morphotypen bekannt. Da die Zuordnung zu Arten noch unklar ist, wurden sie in ihrer Gesamtheit hilfsweise als ‚Epulos‘ bezeichnet.[2]
Einzelnachweise
- Esther R. Angert: Alternatives to binary fission in bacteria. Nature Reviews Microbiology 3, 214-224 (March 2005) doi:10.1038/nrmicro1096
- V. Bresler, W. L. Montgomery L. Fishelson and P. E. Pollak (1998): Gigantism in a bacterium, Epulopiscium fishelsoni, correlates with complex patterns in arrangement, quantitiy and segregation of DNA. In: J Bacteriol. 180 (21); 5601–11; PMID 9791108; PDF (freier Volltextzugriff)
- Website von Esther Angert an der Cornell University, hier
- Mendell, JE. et al. (2008): Extreme polyploidy in a large bacterium. In: Proc Natl Acad Sci USA 105 (18); 6730–4. PMID 18445653 doi:10.1073/pnas.0707522105
- Lev Fishelson, W. Linn Montgomery und Arthur A. Myrberg, Jr. (1985). A Unique Symbiosis in the Gut of Tropical Herbivorous Surgeonfish (Acanthuridae: Teleostei) from the Red Sea. Science 229(4708):49 - 51. doi:10.1126/science.229.4708.49
- W. L. Montgomery, P. E. Pollak (1988): Epulopiscium fishelsoni, N.G., N. Sp., a protist of uncertain taxonomic affinities from the gut of an herbivorous reef fish. In: J. Protozool. 35:565–569. Zitiert nach Bresler et al. 1998.
Weblinks
- Dr. Esther Angerts Seite über Epulopiscium
- Jean Euzéby, Aidan C. Parte: Some prokaryotic names without standing in nomenclature: „Epulopiscium fishelsoni“. In: List of Prokaryotic names with Standing in Nomenclature (LPSN). Abgerufen am 4. Januar 2014.
- Taxonomy Browser Epulopiscium fishelsoni. In: Webseite des National Center for Biotechnology Information (NCBI). Abgerufen am 4. Januar 2014.
- Eintrag zu Epulopiscium im MicrobeWiki des Kenyon College