Beintastler
Die Beintastler (Protura) sind eine Ordnung der Sechsfüßer. Von den 788 im Jahr 2011[1] bekannten Arten der Gruppe sind etwa 200 auch in Mitteleuropa und 41 in Deutschland[2] verbreitet. Die Arten sind sich morphologisch extrem ähnlich und können weltweit nur von einer Handvoll Spezialisten unterschieden werden. Entdeckt wurden die Beintastler erst 1907, von einem Hobbyzoologen bei Genua.[3]
| Beintastler | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
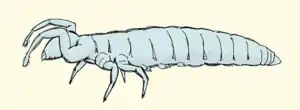
Beintastler (Protura) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Protura | ||||||||||||
| Silvestri, 1907 |
Beintastler leben vor allem im Boden, etwa bis in eine Tiefe von zehn Zentimetern. Allgemein sind sie feuchteliebend und saugen Pilzfäden im Boden aus.
Bau der Beintastler
Beintastler werden 0,5 bis 2,5 Millimeter lang. Sie sind meist weiß oder farblos durchscheinend, selten gelb oder orange, wobei das Hinterende oft kräftiger gefärbt ist. Nur die Gattung Sinentomon ist kräftig rotbraun gefärbt. Der Körper kann jeweils ein Paar einfacher und unverbundener, röhrenförmiger Tracheen besitzen, deren Öffnungen (Stigmen) am zweiten und dritten Rumpfsegment liegen. Diese können aber auch vollständig fehlen.
Kopf und Mundwerkzeuge
Der Kopf der Protura ist im Verhältnis zur Körpergröße klein, er ist rundlich bis birnenförmig, nach vorn etwas dreieckig zugespitzt. Die Mundwerkzeuge zeigen nach vorn (prognath). Es sind weder Augen noch Fühler ausgebildet. Auf der Kopfoberfläche sitzen nur zwei knopfförmig vorgewölbte, kuppelförmige Organe, die als Pseudoculi bezeichnet werden, diese sind vermutlich chemische Sinnesorgane. Die Mundwerkzeuge sind in einer röhrenförmigen Vorstülpung der Kopfwand eingeschlossen und nicht frei sichtbar (entognath), diese Röhre ist nicht vollkommen geschlossen, sondern lässt auf der Unterseite eine Naht (linea ventralis) erkennen. Die Mundwerkzeuge sind als stilettartige Stechborsten ausgebildet. Sie dienen dem Tier wohl vor allem dazu, Pilzfäden (Hyphen) anzustechen und auszusaugen. Als Stechborsten sind die Mandibeln, eine oder beide Laden der Maxillen und beide Laden (Ligula) des Labiums beteiligt. Die Taster (Palpen) der Maxillen sind recht lang und aus der Scheide der Mundwerkzeuge vorgestreckt, sie bestehen aus drei (selten vier) Segmenten. Die Labialpalpen sind klein und unauffällig und meist nur einsegmentig.
Im Inneren des Kopfs ist ein Innenskelett ausgebildet, bei dem umstritten ist, ob es dem Tentorium der anderen Hexapoda homolog ist.
Rumpf und Beine
Der Rumpf der Beintastler besteht hexapoden-typisch aus drei Segmenten, wobei das erste Segment deutlich kleiner ist als die beiden folgenden. Jedes Segment trägt eine Bauchplatte (Sternit), eine Rückenplatte (Tergit) und mehrere kleine, voneinander getrennte Sklerite (Pleurite) an den Seiten. Außerdem trägt jedes Segment ein Beinpaar. Dabei werden nur die beiden hinteren Beinpaare als Laufbeine eingesetzt. Die Vorderbeine werden im Leben angehoben und nach vorn gestreckt getragen, sie dienen als Sinnesorgane und haben im Wesentlichen die Funktionen übernommen, die bei anderen Arthropoden den Antennen zukommen. Die Beine weisen die typische Gliederung der Hexapoden-Beine in Coxa, Trochanter, Femur, Tibia und Tarsus auf. Die Tarsen sind einsegmentig. Der Prätarsus jedes Beins trägt eine einfache Klaue.
Hinterleib
Der Hinterleib der Protura ist langgestreckt-walzenförmig, zum Hinterende hin etwas zugespitzt. Er besteht aus zwölf deutlich erkennbaren Segmenten und besitzt damit ein Segment mehr als alle anderen Hexapoda. Das zwölfte Segment wird oft als Telson bezeichnet, diese Bezeichnung geht auf eine Hypothese zu seiner Entwicklung und Homologie bei anderen Arthropoden zurück, es unterscheidet sich aber in seinem Bau nicht von den anderen. Am Hinterende trägt es einen zungenförmigen Anhang, der manchmal als Furca bezeichnet wird. Paarige Cerci fehlen. Die Segmente acht bis zwölf können teleskopartig ins Hinterende eingezogen und wieder ausgestülpt werden. Die ersten drei Abdominalsegment tragen auf der Bauchseite kurze Extremitäten (sogenannte Styli). Das erste Paar davon besteht aus zwei Segmenten, es besitzt am Ende jeweils ein ausstülpbares Bläschen. Diejenigen des zweiten und dritten Segments können ähnlich gebaut oder zu knospenartigen Rudimenten reduziert sein. Am achten Segment mündet ein Paar große Drüsen aus, die als Wehrdrüsen zur Abwehr von Feinden gedeutet werden, sie geben ein zähes, klebriges Sekret ab. Direkt nachgewiesen wurde dies aber bisher nur von einer Art (Acerentulus sp.).
Die äußeren Genitalien der Protura sind in beiden Geschlechtern sehr ähnlich gebaut. Sie bestehen aus zwei Basalarmen, die jeweils einen Anhang (Stylus) tragen). Bei den Männchen liegen die Geschlechtsöffnungen dabei auf den Armen (sie sind also paarig), beim Weibchen liegt die einzige Geschlechtsöffnung auf dem Basalkörper zwischen ihnen. Sie sind in Ruhelage in eine Genitalkammer zurückgezogen, die sich auf der Bauchseite zwischen den Bauchplatten (Sterniten) elf und zwölf öffnet, und können vorgestreckt werden. Über die Paarung und Befruchtung in dieser Gruppe ist nichts bekannt. Es ist noch nicht einmal klar, ob es zu einer direkten Befruchtung kommt, oder das Männchen eine Spermatophore absetzt, die erst dann vom Weibchen aufgenommen wird (indirekte Befruchtung).[4] Der Bau der Spermien (mit immobilem Flagellum[5]) spricht dabei eher für indirekte Befruchtung.
Fortpflanzung und Entwicklung
Die Weibchen der Protura legen Eier ab, die entweder glatt sind oder eine raue Oberfläche aufweisen, die eventuell dazu dient, bei Überflutung Luft festzuhalten (Plastron). Aus dem Ei schlüpft eine Prälarve mit neun Hinterleibssegmenten, die keine Nahrung aufnimmt. Diese häutet sich zum ersten Larvenstadium mit ebenfalls neun Hinterleibssegmenten. Das zweite Larvenstadium besitzt dann zehn Hinterleibssegmente. Diese häutet sich in ein "Maturus junior" genanntes Stadium mit zwölf Hinterleibssegmenten. Auf dieses Stadium folgt entweder das geschlechtsreife Tier, oder es ist noch ein Subimaginalstadium eingeschoben, das sich von der Imago nur in den noch nicht funktionstüchtigen Geschlechtsorganen unterscheidet. Der Entwicklungsgang, bei dem postembryonale Larvenstadien bei Häutungen noch Segmente hinzugewinnen, wird Anamerie oder Anamorphose genannt. Die Protura sind die einzigen Hexapoda mit Anamorphose.
Die Entwicklungsdauer der einzelnen Stadien und ihr Jahresgang sind offensichtlich ziemlich variabel. Meist wird von einem einjährigen Entwicklungszyklus ausgegangen. Imaginale Tiere sind ganzjährig vorhanden. Im Winter ziehen sie sich nicht in tiefere Bodenschichten zurück.
Obwohl es möglich ist, mit speziellen Techniken Beintastler über längere Zeit im Labor am Leben zu halten, und es auch vereinzelt Nachweise für Eier und Jungtiere unter solchen Bedingungen gibt[6] sind alle Versuche einer Zucht über mehrere Generationen bisher fehlgeschlagen.[4]
Ökologie
Beintastler sind Bodenorganismen. Sie leben im Lückensystem des Bodens, einschließlich des Auflagehumus (einzelne Beobachtungen liegen auch aus verrottetem Totholz oder feuchten, teilzersetzten Pflanzenrückständen vor). Sie kommen normalerweise nie freiwillig an die Bodenoberfläche. Sie bleiben aber auf die oberen Bodenschichten beschränkt, kein Tier wurde bisher tiefer als etwa 50 Zentimeter unter der Oberfläche gefunden.
Aus den bisherigen Beobachtungen wird eine Ernährung vorzugsweise von Pilzhyphen angenommen. Die Tiere sind in der Lage, mit ihren stechborsten-artigen Mundwerkzeugen die Zellwand zu durchstoßen und anschließend das Innere auszusaugen. Zahlreiche Autoren nehmen eine enge Bindung zumindest der meisten Arten an Mykorrhiza-Pilze an; dies sind Pilzarten die in Symbiose mit Pflanzenwurzeln leben. Inwieweit eine andere Ernährungsweise möglich ist und vorkommt, ist umstritten. Frühere Beobachtungen für eine räuberische Ernährung gelten heute als unglaubwürdig.[4] Die Tiere können lange hungern, im Labor überlebten sie bis zu sieben Wochen ohne Nahrungsaufnahme.
Protura leben in Böden aller Art. Ihre Populationsdichte soll aber in sauren Böden höher liegen als in basischen. Ihre Populationsdichte in verschiedenen Böden wurde bei zahlreichen Studien (vorwiegend aus Waldböden) auf zwischen etwa 2.000 und 18.000 Individuen pro Quadratmeter Boden ermittelt. Sie sind damit eine Größenordnung seltener als die ähnlich kleinen Milben und Springschwänze. Über ihre ökologische Bedeutung im Lebensraum ist nahezu nichts bekannt. In ihren Lebensräumen kommen in der Regel zahlreiche Arten vergesellschaftet vor, die bisher höchste an einer Stelle gefundene Artendichte waren 23 Arten in einer Grünanlage der Stadt Wien.[4]
Beintastler sind im Boden bei direkter Nachsuche mit bloßem Auge fast unmöglich zu finden. Sie werden gewöhnlich zur Untersuchung durch Hitzeextraktion (Berlese-Extraktion) aus dem Boden ausgetrieben. Möglich, aber weniger verbreitet ist es, sie über Flotation abzutrennen.
Phylogenie
Die Beintastler werden klassisch innerhalb einer gemeinsamen Gruppe mit den Springschwänzen (Collembola) und Doppelschwänzen (Diplura) als Sackkiefler (Entognatha) zusammengefasst und den Freikieflern (Ectognatha) oder Insekten als Schwestergruppe gegenübergestellt. Als Begründung für diese Taxon-Bildung gilt die Mundtasche, in der die Mundwerkzeuge liegen und die bei allen drei Gruppen vorhanden ist. Diese Gruppierung ist mehrfach aus morphologischen Gründen und aufgrund von Analysen der mitochondrialen DNA bezweifelt worden, gilt aber nach wie vor als wahrscheinlichste Hypothese. Innerhalb der Entognatha werden meist zwei Hypothesen gegenübergestellt: Entweder die Protura und Collembola bilden eine gemeinsame Gruppe, die dann "Ellipura" genannt wird. Oder die Protura und die Diplura bilden eine gemeinsame Gruppe, die in diesem Fall als "Nonoculata" (nach den fehlenden Augen) bezeichnet wird.
Molekulare Studien (aufgrund homologer DNA-Sequenzen) haben mit wenigen Ausnahme die Nonoculata-Hypothese bestätigt.[7][8] Diese Resultate sind aber vorläufig und noch nicht ausreichend abgesichert. Problematisch ist, dass bisher nur wenige Beintastler-Arten sequenziert worden sind. Außerdem ist ihre Sequenz sehr verschieden von derjenigen der anderen Gruppen und zeigt ein ungewöhnliches Übergewicht zugunsten der Basen Guanin und Cytosin in der Zusammensetzung. Die als einziges vollständig analysierte Mitochondriale DNA weist auch strukturell zahlreiche Abweichungen und Besonderheiten auf.[9] Diese Faktoren können eine sogenannte "long-branch-attraction" (in etwa: Anziehung langer Äste) auslösen. Dabei werden ungewöhnliche Sequenzen in der Analyse quasi zum Rand hin "abgedrängt" und können so als Artefakt in der Analyse eine künstliche Gruppierung eingehen. Ein Hindernis der Bearbeitung war bisher, dass die Beintastler-Individuen zur sicheren morphologischen Bestimmung (unter dem Durchlichtmikroskop) auf Objektträger montiert werden müssen, wodurch es nicht mehr möglich war, Material für die DNA-Analyse zu gewinnen. Aufgrund der großen Ähnlichkeit der Arten untereinander konnte man so entweder die Art bestimmen, oder das Tier sequenzieren – aber nicht beides. Dieses Problem ist erst vor kurzer Zeit gelöst worden.[10]
Systematik
_micrograph.jpg.webp)
Die Beintastler werden gewöhnlich in drei Gruppen gegliedert, die im klassischen System den Rang von Unterordnungen erhalten.[8] Innerhalb dieser gibt es insgesamt sieben Familien.[11]
- Acerentomata
- Acerentomidae
- Dies ist die artenreichste Familie der Protura mit über 50 Gattungen und zahlreichen Arten
- Hesperentomidae
- Hesperentomon Price, 1960
- Huhentomon Yin, 1977
- Huhentomon plicatunguis Yin, 1977
- Ionescuellum Tuxen, 1960
- Protentomidae
- Condeellum Tuxen, 1963
- Hinomotentomon Imadaté, 1974
- Neocondeellum Tuxen & Yin, 1982
- Paracondeellum Yin, Xie & Zhang, 1994
- Protentomon Ewing, 1921
- Proturentomon Silvestri, 1909
- Acerentomidae
- Eosentomata
- Antelientomidae
- Antelientomon Yin, 1974
- Antelientomon guilinicum Zhang & Yin, 1981
- Antelientomon prodromi Yin, 1974
- Antelientomon xizangnicum Yin, 1974
- Antelientomon Yin, 1974
- Eosentomidae
- Anisentomon Yin, 1977
- Eosentomon Berlese, 1908
- Isoentomon Tuxen, 1975
- Madagascarentomon Nosek, 1978
- Neanisentomon Zhang & Yin, 1984
- Osientomon Nakamura, 2010
- Paranisentomon Zhang & Yin, 1984
- Pseudanisentomon Zhang & Yin, 1984
- Styletoentomon Copeland, 1978
- Zhongguohentomon Yin, 1979
- Antelientomidae
- Sinentomata
- Fujientomidae
- Fujientomon Imadaté, 1964
- Fujientomon dicestum Yin, 1977
- Fujientomon primum Imadaté, 1964
- Fujientomon Imadaté, 1964
- Sinentomidae
- Sinentomon Yin, 1965
- Sinentomon chui Tuxen & Paik, 1982
- Sinentomon erythranum Yin, 1965
- Sinentomon yoroi Imadaté, 1977
- Sinentomon Yin, 1965
- Fujientomidae
Arten aus Deutschland
Die Systematik der in Deutschland vorkommenden Arten sieht wie folgt aus:[12]
- Acerentomidae
- Acerella Berlese, 1908
- Acerella muscorum (Ionescu, 1930)
- Acerella tiarnea (Berlese, 1908)
- Acerentomon Silvestri, 1907
- Aus dieser Gattung sind 5 Arten in Deutschland nachgewiesen worden
- Acerentulus Berlese, 1908
- Aus dieser Gattung sind 8 Arten in Deutschland nachgewiesen worden
- Berbentulus Tuxen, 1963
- Berbentulus polonicus Szeptycki, 1968
- Gracilentulus Tuxen, 1963
- Gracilentulus europeus Szeptycki, 1993
- Gracilentulus gracilis (Berlese, 1908)
- Nosekiella Rusek, 1974
- Nosekiella danica (Conde, 1947)
- Acerella Berlese, 1908
- Hesperentomidae
- Hesperentomon
- Hesperentomon ulmiacum (Rusek & Stumpp, 1989)
- Hesperentomon
- Protentomidae
- Protentomon
- Protentomon thienemanni Strenzke, 1942
- Proturentomon
- Proturentomon minimum (Berlese, 1908)
- Proturentomon noseki Rusek, 1975
- Proturentomon perpusillum (Berlese, 1909)
- Proturentomon picardi Conde, 1960
- Protentomon
- Eosentomidae
- Eosentomon
- Aus dieser Gattung sind 21 Arten in Deutschland nachgewiesen worden
- Eosentomon
Quellen
- Heinz Janetschek: Ordnung Protura (Beintastler). In: Handbuch der Zoologie. Eine Naturgeschichte der Stämme des Tierreiches. 14. Lieferung. De Gruyter Verlag, 2. Auflage 1970. ISBN 3-11-006373-5.
- O.W. Richards & R.G. Davies: Protura. In: Imms’ General Textbook of Entomology. Springer Verlag, 1977. ISBN 978-0-412-15230-6
- S.L. Tuxen: Protura (Insecta). In: C. T. Duval (editor): Fauna of New Zealand, Number 9. 1985. ISBN 0-477-06765-4
Einzelnachweise
- Günther Pass & Nikolaus Urban Szucsich (2011) 100 years of research on the Protura: many secrets still retained. Soil Organisms 83(3):309–334. Link
- Balkenhol, B. & A. Szeptycki 2003: Verzeichnis der Beintastler (Protura) Deutschlands. – Entomologische Nachrichten und Berichte, Beiheft 8: 7–10.
- http://oe1.orf.at/programm/395430 Vom Leben der Natur. Unbekannte Ur-Insekten. Der Zoologe Günther Pass spricht über die Beintastler. Teil 1: Flügellose Winzlinge. Ö1 (ORF) Radiosendung vom 26. Januar 2015, 08h55 (Serie hat 5 Teile)
- Günther Pass & Nikolaus Urban Szucsich (2011): 100 years of research on the Protura: many secrets still retained. Soil Organisms Volume 83 (3): 309-334.
- B. Bacchetti, R. Dallai, B. Frattello (1973): The spermatozoon of Arthropoda XXII: the 12+0, 14+0 or aflagellate sperm of Protura. Journal of Cell Science 13: 321-335.
- Ryuichiro Machida, Ichiro Takahashi (2004): Rearing technique for proturans (Hexapoda: Protura). Pedobiologia 48: 227–229. doi:10.1016/j.pedobi.2003.01.001
- Dell’Ampio, E., N. U. Szucsich, A. Carapelli, F. Frati, G. Steiner, A. Steinacher, G. Pass (2009): Testing for misleading effects in the phylogenetic reconstruction of ancient lineages of hexapods: influence of character dependence and character choice in analyses of 28S rRNA sequences. – Zoologica Scripta 38: 155–170.
- Emiliano Dell’Ampio, Nikolaus Urban Szucsich, Günther Pass (2011): Protura and molecular phylogenetics: status quo of a young love. Soil Organisms Volume 83 (3): 347-358.
- Wan-Jun Chen, Yun Bu, Antonio Carapelli, Romano Dallai, Sheng Li, Wen-Ying Yin, Yun-Xia Luan (2011): The mitochondrial genome of Sinentomon erythranum (Arthropoda: Hexapoda: Protura): an example of highly divergent evolution. BMC Evolutionary Biology 11: 246 open access
- Alexander Böhm, Daniela Bartel, Nikolaus Urban Szucsich, Günther Pass (2011): Confocal imaging of the exo- and endoskeleton of Protura after nondestructive DNA extraction. Soil Organisms Volume 83 (3): 335-345.
- Protura in GBIF Secretariat (2019). GBIF Backbone Taxonomy. Checklist dataset abgerufen via GBIF.org am 23. Januar 2021.
- Spelda J. Taxon list of Protura from Germany compiled in the context of the GBOL project. Staatliche Naturwissenschaftliche Sammlungen Bayerns. Checklist dataset abgerufen via GBIF.org am 23. Januar 2021.
Weblinks
- Protura Eintrag in Tree of Life Web Project
- Protura
- Beintastler (Protura)
- Beintastler - Proturans (Memento vom 17. Mai 2008 im Internet Archive)