Arkto-alpine Disjunktion
Arkto-alpine Disjunktion wird eine auffällige Arealaufspaltung einer Gruppe von Tieren und Pflanzen genannt, die in Lebensräumen der Tundra oder Kältesteppe, einerseits in Hochgebirgen der temperaten Zone (seltener subtropisch-mediterranen Gebirgen) und andererseits der Arktis, auftritt. In den zwei Teilarealen, die durch eine breite Lücke im mitteleuropäisch-osteuropäischen Flach- und Hügelland voneinander getrennt sind, kommen dann eine Vielzahl derselben Arten vor. Dieses Verbreitungsbild geht meist auf ein ehemals im Eiszeitalter zusammenhängendes Areal der kälteliebenden Arten zurück, das durch die seitherige Erwärmung verkleinert und dabei in Teilareale aufgespalten worden ist[1]. Vorkommen von Tieren und Pflanzen in isolierten, kleinen Habitaten, etwa einzelnen Berggipfeln, sind dann als Glazialrelikte der Eiszeiten aufzufassen.[2] Der Arealtyp, der sowohl in der geobotanischen als auch in der zoogeographischen Literatur sowie phylogenetisch gründlich untersucht wurde,[3] ist vor allem in Eurasien zu finden, da hier die großen Gebirgsketten latitudinal von West nach Ost verlaufen und zusätzlich zwischen den arktischen und temperaten Regionen Flachländer vorherrschen. In Nordamerika tritt die arkto-alpine Disjunktion nicht auf, da die Gebirgsketten überwiegend in longitudinaler Erstreckung verlaufen[4] sowie Borealer Nadelwald während der maximalen Vereisungsphase bis in unmittelbarer Nähe zum Laurentinischen Eisschild wuchs.[5]

Klimageschichte
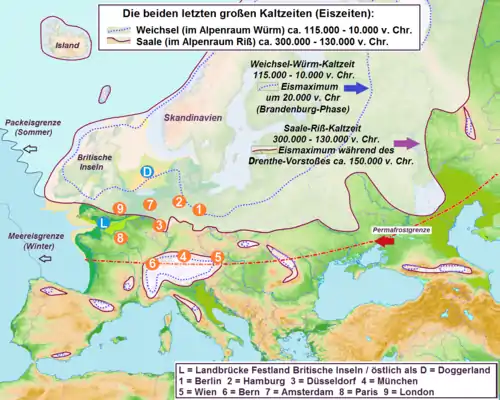
Die Gemeinsamkeit der Arealform bei verschiedenen Gruppen von Lebewesen kann auf einen gemeinsamen klimageschichtlichen Hintergrund zurückgeführt werden. Da sich bei vielen Artgruppen eine positive Korrelation zwischen der Ausdehnung der paläarktischen, eurasiatischen und holarktischen Teilareale feststellen lässt, muss in der Vergangenheit eine zonale Ausbreitungsmöglichkeit bestanden haben. Dies wird durch die Ausdehnung der Periglazialgebiete im Pleistozän erklärt, die in den Kaltzeiten des Glazials eine zusammenhängende Großlandschaft zwischen Atlantik und Pazifik gebildet hatten. In der über eine Million Jahre andauernden Periode der Eiszeiten wirkten sich mindestens sechs Glazialphasen mit abnehmenden Temperaturen auf den Zuwachs der Eismassen sowie auf Prozesse der Landschaftsdynamik, wie zum Beispiel die Bildung von Permafrostboden und weitflächigen Erscheinungen von Periglazialgebieten, aus. Dies hatte ebenfalls stark regressive Auswirkungen auf die vormals vorhandene Vegetation.[6]
Der Nachweis arkto-alpiner Elemente in den temperaten und meridionalen Gebirgsketten bildete neben quartärgeologischen Nachweisen eines der entscheidenden Argumente für die These, dass Inlandseis und Eiskappen zum Rückzug der arkto-tertiären Vegetation (u. a. Persischem Eisenholz oder Zelkova) geführt hatten. Nach deren Rückzug wurden sowohl im hohen Norden, als auch in höheren Bergregionen geeignete Lebensbedingungen vorgefunden und besiedelt. Als Leitart der arkto-alpinen Flora wird das Massenaufkommen von Dryas (Silberwurz) genommen. So kann mit dem fossilen Vorkommen von Dryas zwischen Süd- und Mittelengland, der Bretagne, den Benelux-Staaten, Mittel- und Osteuropa sowie Zentralrussland die Ausdehnung der ehemaligen kaltzeitlichen Tundra in Europa rekonstruiert werden.
Austausch von Flora und Fauna


Während der Eiszeiten wurde der arkto-tertiäre Formenbestand fast vollständig ausgeschaltet, die Wald- und Hochgebirgsflora in Refugialgebiete zurückgedrängt. Auf der anderen Seite wurde der Florenaustausch mit anderen Gebirgen und den nördlichen Regionen beträchtlich intensiviert. So sind arkto-alpine Arten von den Picos de Europa in Nordspanien bis in die Japanischen Alpen verbreitet.
Ein Beispiel einer eurasiatisch-amerikanisch-arktisch verbreiteten Art ist der Alpenapollo, dessen europäisches Teilareal nur die zentralen Alpen umfasst. Erst vom Ural an besiedelt er Sibirien, die Mongolei, Alaska, Kanada und die westlichen USA bis New Mexico. Jedoch fehlt er in Europa in Skandinavien oder Island. Nicht alle der kaltzeitlichen Arten konnten ein arkto-alpines Areal ausbilden. Ein Großteil der Tundrenarten besiedelt nur eines der Teilareale und wird dadurch jeweils in alpine oder arktische Sippenpaare getrennt. Beispiele der zwischen Arktis und temperaten alpinen Hochgebirgen divergenten Sippenpaare sind Salix retusa-Salix polaris,[7] oder Oxytropis dinarica-Oxytropis campestris.
Gleiches Schema findet sich auch in der Fauna der Paläarktis. Es lassen sich auch mehrere Arealformen arkto-alpiner Disjunktion beschreiben. So gibt es Verbreitungsareale zwischen europäischen und asiatischen Hochgebirgen und der Arktis oder solche die nur eines der Hochgebirgssysteme und die Arktis umfassen. Seltener sind Fälle rein europäischer arkto-alpiner Disjunktion.[8] Folgende Schmetterlinge besitzen ein europäisches arkto-alpines Areale: Pyrgus andromedae oder Pygmaeana fusca. In Asien umfasst es aber mehr Arten: u. a. Pyrgus centaureae, Colias hecla, Euphydras iduna, Boloria alaskensis oder Oeneis bore. Zum sibirischen arkto-alpinen Teilareal gehören Altai und Sajan sowie die sibirische Arktis. Auch bei Schmetterlingen finden sich Sippenpaare die sich in arktische und alpine Formen trennen lassen. Die südlichste Verbreitungsgrenze solcher alpinen Arten mit nächstverwandten arktischen liegt in den Hochgebirgen der Balkanhalbinsel. Es sind: Agriades glandon – Agriades aquilo, Colostygia lineolata – Colostygia turbata, Holoarctia cervini – Holoarctia puengeleri u. a.[9]
Durch reproduktive Isolation der stark zersplitterten Vorkommen einzelner Hochgebirge haben zahlreiche arkto-alpinen Arten oft Unterarten gebildet, die erst nach den Eiszeiten durch geographische Trennung entstanden sind. Auch unter diesen gibt es Disjunktionen nur zwischen den im gleichen Zonobiom liegenden Gebirgen und Landschaften. Diese Arealform wird alpine Verbreitung bezeichnet. Sie gilt für Arten, die in den alpinen Stufen einzelner Hochgebirge Eurasiens verbreitet sind, aber nie in der Arktis auftreten. Ein Beispiel einer rein oreotundralen Art ist Boloria pales. Es ist ein in alpinen Tundren lebender Schmetterling, der vom Kantabrischen Gebirge bis nach Taiwan für einzelne Hochgebirgen auftritt und noch die Gebirge Süd-Sibiriens erreicht (Altai, Sajangebirge), ohne in die Arktis vorzudringen.[10] Weitere Beispiele der alpinen Verbreitung in Europa sind der Eros-Bläuling sowie zahlreiche Erebien wie Erebia oeme, Erebia albergana, Erebia pronoe, Erebia manto, Erebia sudetica, Erebia tyndarus, Erebia pharte, Erebia epiphron oder Erebia gorge. Ein vikariierndes Sippenpaar der arkto-alpinen Fauna sowie den oro-Mediterranen xeromontanen Gebirgen zwischen Artkis der Alpen, der Iberischen und Balkanhalbinsel sowie Kleinasiens sind Agriades glandon – Agriades pyrenaicus – Agriades dardanus. Während Agriades glandon eine polyzentrische eurasiatische (zirkumpolare) tundro-alpine Art und Vertreter der alpiden Fauna ist, vollzog sich bei den anderen die jeweilige Artbildung in meridionalen Refugien während der Eiszeiten, die höchstwahrscheinlich in größter Nähe zu den heutigen stark zersplitterten Standorten lagen und Larvalhabitate mit dem Zottigen Mannsschild (Androsace villosa) anboten.[11][12]
Die Adamović-Linie
Eine strenge Grenze nach Süden besitzen die Arten, die zur arkto-alpinen Arealgruppen zählen, in der sogenannten Adamović-Linie. Diese scharfe Trennungslinie tritt auf der Balkanhalbinsel in Erscheinung. Sie ist die Trennungslinie zwischen der mitteleuropäischen und mediterranen Vegetationszone.[11] Südlich der Adamović-Linie treten praktisch keine arkto-alpinen Elemente mehr auf. De Lattin hatte diese von Lujo Adamović 1907 aufgrund pflanzengeographischer Grundzüge postulierte Grenze als Stauungszone sibirischer Faunenelemente erkannt. Der ungarische Entomologe Zoltán Varga hat die Adamović-Linie als Grenze zwischen alpinen und xeromontanen Schmetterlingsarealtypen weiter bekräftigt. Neben den klimatischen Ursachen sind Substrattypen, Geomorphologie und Bodenverhältnisse und dadurch bedingte Unterschiede in der Vegetation für diese faunistischen Unterschiede verantwortlich. Für die genetischen Linien der südosteuropäischen Hochgebirge mit typischen alpiner Zonierung konnte bei arkto-alpinen und alpinen Disjunktionen eine frühere genetische Trennungen festgestellt werden. Sie gehören älteren genetischen Linien an, die dort an Ort und Stelle die Eiszeiten überdauert haben und im Gegensatz zu den Alpinen, Relikte der älteren Eiszeitphasen stellen.[13]
Phylogenetische Zeugnisse

Die arkto-alpinen Arten stellen phylogenetisch gut untersuchte Gruppen. Sowohl in der Fauna (Pardosa saltuaria, Graubrauner Mohrenfalter und Nebria rufescens) als auch in der Vegetation konnten die vermuteten Zusammenhänge einer durchgehenden Besiedlung in Tundren zwischen den Eiszentren der Alpen und dem Nordischen Inlandeis durch phylogenetische Verwandtschaft belegt werden. Erst in den südlicher liegenden balkanischen Gebirgen wurden ältere Stammlinien gefunden, die nicht auf einer nacheiszeitlichen Einwanderung basieren.
Unter den arkto-alpinen Pflanzenarten konnte für Ranunculus glacialis, Arabis alpina und Veronica alpina eine Besiedlung der Nordwest-Arktis aus den Alpen heraus gezeigt werden. Bei Dryas octopetala erfolgte die Kolonisierung von Nordeuropa und Skandinavien aus über die Stammpopulationen zwischen den beiden Vereisungszentren. Während bei Minuartia biflora wie Carex atrofusca die Populationen in Nordeuropa und den Alpen schon über längere Zeit separiert waren. Bei Ranunculus pygmaeus waren die nächstverwandten der Alpinen und Skandinavischen Populationen die der Taimyr-Halbinsel im nördlichen Sibirien und im Ural.[14]
Habitattypen
Arkto-alpine Arten sind immer an waldfreie Habitate gebunden.[15] Sie besiedeln oft Fels- und Schuttstandorte oder Hochmoore, die zumeist durch periglaziale Prozessdynamik, also ständigen Wechsel zwischen Tau- und Frostperioden, geprägt werden. Das Verbreitungsbild dieser Tundrenarten entstand durch Expansion während der Kaltzeiten mit anschließendem Zurückweichen in der nacheiszeitlichen Klimaerwärmung. Ursprünglich besiedelten die arkto-alpinen Arten weite Gebiete der zur Eiszeit baumfrei gewordenen Tundren in Mittel- und Osteuropa. Diese Gebiete zwischen den Vereisungszentren in den Alpen und Nordeuropas besaßen weite Urstromtäler mit periglazialen Frostschuttzonen, in denen sich die eiszeitlichen Tierarten aufhielten. Nach Abschmelzen der Inlandgletscher sowie dem Zurückweichen der Alpengletscher wichen die kälteliebenden Arten sowohl in höhere Breiten als auch, im Süden, in höhere Lagen aus.
Warmzeitliche Mikrorefugien arkto-alpiner Arten

Anders als die kaltzeitliche Fauna wurde keine der arkto-alpinen Pflanzenarten durch die klimatischen Oszillationen im Quartär ausgerottet.[5] Einige der wärmeempfindlichsten Arten wurden aber in den Interstadialen aus regionalen Floren verdrängt. Während des 1200 Jahre dauernden kaltklimatischen Rückfalls in volle Glazialbedingungen zur Jüngeren Dryas-Zeit kehrten u. a. Pedicularis hirsuta, Pecicularis lanata, Silene uralensis und Salix polaris nicht mehr in das kontinentale Europa oder auf die Britischen Inseln zurück. Interessanterweise besiedeln diese Arten in Europa nur noch hochliegende Standorte in Skandinavien.[16] Viele arkto-alpine Arten sind heute nach dem Abklingen des Glazialen Maximums oder dem Ende des Jüngeren Dryas in Großbritannien und Irland ausgestorben. Neben den schon erwähnten Pedicularis hirsuta, Pecicularis lanata, Silene uralensis und Salix polaris sind es Coriospermum pallasii, Papaver Sect. Scapiflora, Ranunculus hyperboreus, Ranunculus aconitifolius, Silene uralensis und Stellaria crassifolia. Reliktisch auf Mikrorefugien angewiesen sind auf den Britischen Inseln Arenaria ciliata, Arenaria norvegica ssp. anglica, Aregaria norvegica ssp. norvegica, Astragalus alpinus, Koenigia islandica, Linnaea boralis, Lychnis alpina, Minuartia stricta, Minuartia rubella, Polemonium caerulaeum, Saxifraga cespitosa u. a.
Nicht alle der Arten wanderten in den Interglazialen ebenso nach Norden in hohe Breiten oder in hoch liegende Stufen in Gebirgen ab. Solche als Mikrorefugien (engl. microrefugia oder "cryptic refugia") bezeichnete Standorte werden auch heute in der nachglazialen Periode beobachtet.[17] Mikrorefugien in niederen Höhenstufen oder niedrigen Breiten treten in Europa dort auf, wo die standörtlichen Verhältnisse wettbewerbsfähigere, wärmeliebende (thermophile) Arten benachteiligen.[5][18][19] In Südeuropa sind Glazialrelikte nur noch in den höchsten Gebirgen und dort oft nur noch in topographischen Gunsträumen anzutreffen. Insbesondere trifft dies auf die Regionen an der Grenze zur mediterranen Florenregion zu. In Bulgarien beschränkt sich der Alpenspitzkiel im fast 3000 m hohen Pirin auf periglaziale Schuttdecken oder Solifluktionböden und Fließerden in Höhen um 2600 m.[20] Beispiel für extrem niedrige liegende Refugien in Gebirgen die nicht mehr hoch genug aufragen, als auf ihnen noch einen alpine Stufe gebildet ist, finden sich in Dolinen, in denen sich Kaltluft ansammeln kann. Diese sind in verschiedenen Regionen wie dem Apennin oder den Dinariden Rückzugsorte der Glazialflora. So finden sich im Orjen-Gebirge die mittelmeernächsten reliktischen Standorte von Dryas octopetala nur noch innerhalb der Begrenzung eines Kaltluftsees in einer Höhenlage von nur 1575 m. Auch hier ist die Ausbildung von periglazialen Nischen für sein auftreten notwendig.[21]
Beispiele

Bekannte Beispiele sind im Pflanzenreich unter anderen Silberwurz (Dryas octopetala), Kraut-Weide (Salix herbacea), Einköpfiges Berufkraut (Erigeron uniflorus), Schnee-Enzian (Gentiana nivalis), Dreiblatt-Binse (Juncus trifidus), Nacktried (Kobresia myosuroides), Alpen-Spitzkiel (Oxytropis campestris), Alpen-Rispengras (Poa alpina), Knöllchen-Knöterich (Bistorta vivipara), Stängelloses Leimkraut (Silene acaulis) oder Zweiblütiges Veilchen (Viola biflora). Insgesamt sind in Europa 136 Pflanzenarten als arkto-alpine Elemente einzustufen. 131 kommen in den Alpen, 110 in den Karpaten, 92 in den Pyrenäen, 77 auf der Balkanhalbinsel und 58 im Apennin vor.[4]
Im Tierreich kommen in fast allen Gruppen arkto-alpine Arten vor. Insbesondere sind sie bei den Schmetterlingen gut untersucht. Darunter fallen unter anderen Pyrgus andromedae, Boloria napaea und Erebia pandrose.[22] Für viele Schmetterlingsarten mit arkto-alpiner Disjunktion sind enge Verbindungen zwischen Larvalhabitat, Futterpflanze sowie biogeomorphologischer Interaktionen anzunehmen. Die Raupen von Pyrgus andromedae, ernähren sich monophag von der Silberwurz. Daher ist diese Schmetterlingsart schon aufgrund der Verbreitung der Futterpflanze kaum außerhalb periglazialer Schuttstandorte anzutreffen.[11]
Bei den Vögeln ist das Alpenschneehuhn in den Hochgebirgen Pyrenäen und Alpen sowie disjunkt zirkumpolar verbreitet. Ähnlich, aber ohne ein Teilareal in den Pyrenäen, ist bei Säugetieren der Schneehase verbreitet. Neben der zirkumpolaren Verbreitung besteht bei ihm in den Alpen ein glaziales Reliktareal.
Hauptzentren der arkto-alpinen Disjunktion sind die Alpen, Karpaten, Pyrenäen und die höchsten Gebirge der Balkanhalbinsel (ausschließlich des Olymp). Auf der Balkanhalbinsel treten, da es dort keine durchgehenden Hochgebirge gibt, arkto-alpine Elemente nur lokal auf, insbesondere in Montenegro (Durmitor, Prokletije), Kosovo (ebenfalls Prokletije), im nördlichen Nordmazedonien (Šar Planina) sowie im westlichen Bulgarien (Rila und Pirin).
Literatur
- Vladimir Stevanović, Snežana Vukojičić, Jasmina Šinžar-Sekulić (2009): Distribution and diversity of Arctic-Alpine species in the Balkans. Plant Systematics and Evolution 283 (3/4): 219-235.
Einzelnachweise
- Lexikon der Biologie: arktoalpine Formen, Spektrum.de
- Akira S. Hirao, Mikio Watanabe, Shiro Tsuyuzaki, Ayako Shimono, Xuefeng Li, Takehiro Masuzawa, Naoya Wada 2017: Genetic diversity within populations of an arctic–alpine species declines with decreasing latitude across the Northern Hemisphere. Journal of Biogeography, vol. 44: 2740-2751. (Wiley Online Library:PDF)
- Schmitt, T.; Muster, C. & Schönswetter, P. 2010: Are Disjunct Alpine and Arctic-Alpine Animal and Plant Species in the Western Palearctic Really “Relics of a Cold Past”? In: Relict Species Phylogeography and Conservation Biology, 2010: 239-252
- Vladmir Stevanović, Snežana Vukojičić, Jasmina Šinžar-Sekulić, Maja Lazarević, Gordana Tomović, Kit Tan: Distribution and diversity of Arctic-Alpine species in the Balkans. Plant Systematics and Evolution, Dezember 2009, 283.219, (PDF)
- Hilary H. Birks 2008: Late-Quaternary history of arctic and alpine plants. Plant Ecology & Diversity. Vol. 1/2: November 2008: 135-146
- Thorsten Englisch 1999: Multivariate Analysen zur Synsystematik und Standortsökologie der Schneebodenvegetation (Arabidetalia caerulea) in den Nördlichen Kalkalpen. In: Stapfia. Band 59, Linz 1999, S. 30–31, ISSN 0252-192X, zobodat.at [PDF]
- Thorsten Englisch 1999: S. 31
- Varga, Zoltan und Schmitt, Thomas 2008: Types of oreal and oreotundral disjunctions in the western Palearctic. Biological Journal of the Linnean Society, vol. 93: 415-430.
- Varga, Zoltan und Schmitt, Thomas 2008: S. 422–423
- Yu-Feng Hsu 1997: Notes on Boloria pales yangi, ssp. nov., a remarkable disjunction in butterfly biogeography (Lepidoptera: Nymphalidae). Journal of Research on the Lepidoptera, vol. 34/1-4: 142-146 (researchgate:PDF)
- Zoltan Varga (2014): Biogeography of the high mountain Lepidoptera in the Balkan Peninsula. Ekologija Montenegrina 1: 140-168. (PDF)
- Varga, Zoltán 1977: Zoogeographische Gliederung der paläarktischen Orealfauna. In: Verhandlungen des Sechsten Internationalen Symposiums über Entomofaunistik in Mitteleuropa (Hrsg. Malicky, Hans), Springer Netherlands, 263-284.
- Schmitt, T.; Muster, C. & Schönswetter, P. 2010: Are Disjunct Alpine and Arctic-Alpine Animal and Plant Species in the Western Palearctic Really “Relics of a Cold Past”? In: Relict Species Phylogeography and Conservation Biology, 2010: 239-252
- Schmitt, T.; Muster, C. & Schönswetter, P. 2010: Are Disjunct Alpine and Arctic-Alpine Animal and Plant Species in the Western Palearctic Really “Relics of a Cold Past”? In: Relict Species Phylogeography and Conservation Biology, 2010: 239-252
- Eintrag arktisch-alpin in Lexikon der Biologie, Spektrum.de
- R.M.M. Crawford 2008: Cold climate plants in a warmer world. Plant Ecology & Diversity, 1/2: November 2008: 285-297. (PDF)
- Richard J. Abbott 2008: History, evolution and future of arctic and alpine flora: overview. Plant Ecology & Diversity. Vol. 1/2: November 2008: 129-133 (PDF)
- H. John. B. Birks und Katherine Willis 2008: Alpines, trees, and refugia in Europe. Plant Ecology & Diversity. Vol. 1/2: November 2008: 147-160
- R.M.M. Crawford 2008: Cold climate plants in a warmer world. Plant Ecology & Diversity, 1/2: November 2008: 285-297. (PDF)
- Kozuharova, E., Richards, A., Hale, M., Benbassat, N. und Wolff, K. 2015: Another brick in the wall of the oxytropis campestris complex with an emphasis of three members of this group from Pirin Mts, the Balkans. In: Fabaceae (Hrsg. Wayne Garza), Nova Science Publishers, 61-108. ISBN 978-1-63482-200-8
- Pavle Cikovac & Ingo Hölzle 2018: Glacial relicts in the Mediterranean Dinarides - A phenomenon of cold-air poll microclimates?. Botanica Serbica, vol. 42/1: 74. (Botanica Serbica:PDF)
- Zoltan Varga (1996): Biogeography and evolution of boreal Lepidoptera in the Palaearctic. Acta Zoologica Academiae Scientiarum Hungaricae 42: 289-330.