Zwergelefant
Als Zwergelefanten bezeichnet man eine Reihe von überwiegend ausgestorbenen, kleinwüchsigen Rüsseltierformen, die sich im Eiszeitalter auf verschiedenen Inseln entwickelt haben. Sie gehören zumeist den Gattungen Palaeoloxodon und Mammuthus an, es sind aber auch aus anderen Rüsseltierlinien verzwergte Vertreter belegt. Zwei heute lebende Formen, der „Borneo-Zwergelefant“ und der afrikanische „Zwergelefant“, sind in ihrer taxonomischen Stellung umstritten.

Ausgestorbene Insel-Zwergelefanten
Als Zwergelefanten oder besser Insel-Zwergelefanten bezeichnet man eine Reihe von kleinwüchsigen Elefanten der Gattungen Palaeoloxodon und Mammuthus. Dabei ist die Zwergwüchsigkeit eine Anpassung an den verkleinerten Lebensraum auf den Inseln, das verringerte Nahrungsangebot und die Absenz von Raubtieren. Derartige verkleinerte Inselformen kamen als Ergebnis eines als Inselverzwergung bezeichneten Prozesses im Laufe der Rüsseltierevolution häufiger vor, wie zum Beispiel Stegodon auf den Inseln Südostasiens. Neben den Elefanten unterlagen auch andere großwüchsige Säugetiere, wie etwa das Flusspferd mit den Arten Hippopotamus melitensis, Hippopotamus pentlandi und Hippopotamus minor auf einigen Mittelmeerinseln, dem Prozess der Inselverzwergung. Solche Populationen sind jedoch anfällig gegen Änderungen der Umweltbedingungen wie Naturkatastrophen oder Einflüsse des Menschen. Nach einzelnen Untersuchungen bildet die Isolation der Insel keinen entscheidenden Faktor bei der Verzwergung. Dagegen haben die Größe der Insel, die Zeit, die seit der Trennung von der Hauptpopulation vergangen ist und Konkurrenz mit anderen Arten Einfluss auf den Prozess. Teilweise wirken diese gegenläufig. Auf Inseln mit einer Entfernung von 6 bis 10 km zum Festland erfolgte in der Regel kaum eine Verzwergung, da offensichtlich ein genügend großer genetischer Austausch zwischen den verschiedenen Populationen stattfand.[1][2] Für den Sizilianischen Zwergelefanten (Palaeoloxodon falconeri) von Sizilien und Malta wurde die Verzwergungsrate bestimmt. Ausgehend von einer Kolonialisierung der Inseln vor rund 70.000 Jahren liegt die Reduktion des Körpergewichts bei rund 18,45 kg und die der Schulterhöhe bei rund 3,41 mm je Generation. Eine frühere Trennung von der Hauptpopulation vor gut 520.000 Jahren ergibt entsprechend 0,79 kg und 0,15 mm je Generation.[3]
Inselelefanten des Mittelmeers
Auf einigen Inseln des Mittelmeers lebten im Pleistozän Zwergformen, die sich zu einem Großteil aus dem Europäischen Waldelefanten (Palaeoloxodon antiquus) entwickelten, welcher das Festland besiedelte. Weiterhin kamen, wenn auch seltener, Arten vor, die auf die Gattung Mammuthus zurückzuführen sind. Deren Ausgangsformen können einerseits der Südelefant (Mammuthus meridionalis), andererseits auch das Steppenmammut (Mammuthus trogontherii) sein. Abzüglich gewisser Schwankungen innerhalb der einzelnen Populationen kann für diese drei festländischen Arten ein Körpergewicht zwischen 10 und 15 t angegeben werden.[4][5]
Zwergelefanten

Zu den bekanntesten verzwergten Verwandten des Europäischen Waldelefanten gehört unter anderem der Sizilianische Zwergelefant (Palaeoloxodon falconeri) mit einer Schulterhöhe von etwa einem Meter und einem Gewicht von rund 170 kg. Diese Art kam sowohl auf Sizilien als auch auf Malta vor, die beide in den Kaltzeiten des Pleistozäns, als der Meeresspiegel deutlich niedriger lag als heute, einen wohl enger geschlossenen geographischen Raum bildeten. Die Art entstand im mittleren Pleistozän in einer ersten Phase der geographischen Isolierung. Eine zweite Phase der Isolierung führte im späten Pleistozän auf Sizilien und Malta zur Bildung der Art Palaeoloxodon mnaidriensis, die etwas weniger als 2 m an der Schulter maß und bis zu 2,5 t wog.[6][7] Dagegen ist der ehemals als Maltesischer Zwergelefant beschriebene Elephas melitensis identisch mit Palaeoloxodon mnaidriensis.[7][8] Etwa ähnlich groß wie Palaeoloxodon mnaidrensis wurde eine noch unbestimmte Form auf Favignana westlich von Sizilien, deren Alter auf rund 20.000 Jahre vor heute datiert und damit die wahrscheinlich jüngsten Belege von Zwergelefanten im westlichen Mittelmeer repräsentieren. Die heutige Insel war aber zu diesem Zeitpunkt mit Sizilien verbunden.[9]

Auch auf den heute teilweise winzigen ägäischen Inseln des Dodekanes wie Astypalea, Tilos und Rhodos sowie der Kykladen, etwa Delos und Naxos gab es ähnliche Formen. Genauer untersucht ist Palaeoloxodon tiliensis von Tilos[10] mit einer Schulterhöhe von 1,4 m und einem rekonstruierten Gewicht von 650 kg.[7] Eines der jüngsten datierten Skelette von der Insel Tilos weist ein Alter von 2400 v. Chr. auf,[11] der letzte Nachweis liegt bei etwa 1300 v. Chr.[7] Es ist daher möglich, dass bronzezeitliche Kulturen des Mittelmeerraums diesem Zwergelefanten begegnet sind und etwas mit deren Aussterben zu tun hatten; ob Darstellungen auf altägyptischen Wandmalereien solche Zwergelefanten zeigen, ist bis heute ein Streitpunkt.[12] Die Schädel der Zwergelefanten könnten im Altertum zur Entstehung der mediterranen Sage von den Zyklopen beigetragen haben, da Elefanten in der Mitte der Stirn eine einzige Nasenöffnung besitzen, die man leicht als Auge missdeuten kann.[13] Die meisten anderen Formen sind bis jetzt unbenannt, lediglich die Funde von Naxos wurden im Jahr 2014 als Palaeoloxodon lomolinoi beschrieben, das nur 10 % der Größe eines Europäischen Waldelefanten erreichte.[14][15][16][2] Während der maximalen Vereisungsphasen der Kaltzeiten des jüngeren Pleistozäns lag der Meeresspiegel teilweise rund 100 m tiefer als heute, wodurch einige Inseln der Kykladen eine größere Landmasse bildeten. Diese in Fachpublikationen als „Paläo-Kykladen“ bezeichnete Insel schloss unter anderem Naxos, Paros und Delos ein, auf denen Nachweise von Elefanten gelangen.[14]
Eine mit nur 250 kg sehr kleine Zwergform stellt Palaeoloxodon cypriotes von Zypern dar. Mehr als 200 Knochenreste wurden unter anderem in der Höhlenruine von Aetokremnos dokumentiert, gemeinsam mit unzähligen Resten eines verzwergten Flusspferdes. Da die Höhle auch Hinterlassenschaften menschlicher Nutzung barg und die Fundschichten auf rund 9500 v. Chr. datiert sind, gehen einige Wissenschaftler davon aus, dass die Tiere möglicherweise von den ersten Besiedlern der Insel ausgerottet wurden,[17][18][7] was aber nicht unumstritten ist.[19] Palaeoloxodon xylophagou repräsentiert eine zweite verzwergte Form auf Zypern. Sie erreichte etwas größere Ausmaße und datiert in das Mittelpleistozän.[20] Dem gegenüber war Palaeoloxodon creutzburgi aus dem Jungpleistozän von Kreta weniger stark verzwergt. Die Art erreichte etwa 69 % der Körpergröße der Festlandsformen, Gewichtsschätzungen reichen anhand der aufgefundenen Langknochen bis etwa 3 t. Funde von der östlich benachbarten Insel Kasos sind möglicherweise ebenfalls Palaeoloxodon creutzburgi zuzuweisen. Ebenfalls von Kreta wurde Palaeoloxodon chaniensis berichtet.[21] Dessen taxonomische Eigenständigkeit ist aber umstritten. Die Form übertraf Palaeoloxodon creutzburgi etwas in der Größe.[7][15][16] Von der ionischen Insel Kefalonia im Golf von Patras wurde im Jahr 2018 anhand eines einzelnen Oberkiefers die Art Palaeoloxodon cephallonicus beschrieben. Der Fund datiert in den Übergang vom Mittel- zum Jungpleistozän und verweist auf ein Tier mit Ausmaßen vergleichbar zu Palaeoloxodon creutzburgi oder Palaeoloxodon mnaidriensis.[22] Ein Jahr später wurde in einer Publikation angezweifelt, dass es sich bei dem Oberkieferrest um eine auf Kefalonia endemische Art handelt. Die Autoren der Studie ordnen stattdessen den Fund dem Europäischen Waldelefanten zu und nehmen eine mit den Festlandsformen übereinstimmende Körpergröße an. Der Europäische Waldelefant ist auf Kefalonia ebenfalls belegt.[2]
Zwergmammute

Zu den Vertretern der Mammute zählt unter anderem das im Spätpleistozän auf Sardinien vorkommende Mammuthus lamarmorai, welches etwa 1,5 m groß wurde und rund 800 kg wog. Das stark fragmentierte Fundmaterial stammt weitgehend vom westlichen Teil der Insel.[7][23] Ein weiterer Vertreter war das ursprünglich in seiner taxonomischen Stellung umstrittene, stark verzwergte Kreta-Zwergmammut (Mammuthus creticus) aus dem Frühpleistozän,[7] welches vor einigen Jahren mittels molekulargenetischer Untersuchungen (vergl. aDNA) als tatsächlich zu den Mammuten zu zählende Art identifiziert wurde.[24] Neueren Untersuchungen zufolge ist diese Mammutart mit einer Schulterhöhe von 1,1 m und einem Gewicht von rund 310 kg die bisher kleinste bekannte Form dieser Rüsseltiergattung. Bisher ist mit Kap Maleka auf der Halbinsel Akrotiri nur eine Fundstelle bekannt.[25][2]
Übersicht der Nachweise auf Mittelmeer-Inseln
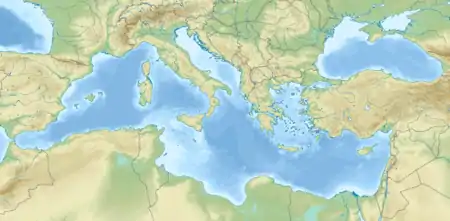 Zwergelefant (Mittelmeer)
| |||||||||
Mittelmeer-Inseln mit Nachweisen von Zwergelefanten der Gattung Palaeoloxodon ( |
Italien, Malta
| Insel | Taxon | Autor | Datierung |
|---|---|---|---|
| Sardinien | Mammuthus lamarmorai | (Major, 1883)[26] | Mittel-/Jungpleistozän |
| Favignana | Palaeoloxodon sp. | Palombo et al., 2020[9] | Jungpleistozän |
| Malta | Palaeoloxodon falconeri | (Busk, 1869)[27] | Mittel-/Jungpleistozän |
| Palaeoloxodon mnaidriensis | (Adams, 1874)[28] | Mittel-/Jungpleistozän | |
| Sizilien | Palaeoloxodon falconeri | (Busk, 1869) | Mittel-/Jungpleistozän |
| Palaeoloxodon mnaidriensis | (Adams, 1874) | Mittel-/Jungpleistozän |
Griechenland
| Insel | Taxon | Autor | Datierung |
|---|---|---|---|
| Astypalea | Palaeoloxodon sp.A1 | Athanassiou et al., 2019[2] | unklar |
| Kasos | Palaeoloxodon aff. creutzburgiA2 | Sen et al., 2014[15] | Jungpleistozän |
| Rhodos | Palaeoloxodon sp. | Symeonides et al., 1974 | Jungpleistozän |
| Tilos | Palaeoloxodon tiliensis | (Theodorou et al. 2007)[10] | Jungpleistozän/Holozän |
| Insel | Taxon | Autor | Datierung |
|---|---|---|---|
| Kreta | Mammuthus creticus | (Bate, 1907)[29] | Alt-/Mittelpleistozän |
| Palaeoloxodon chaniensisA3 | (Symeonides et al., 2000)[30] | Jungpleistozän | |
| Palaeoloxodon creutzburgi | (Kuss, 1965)[31] | Mittel-/Jungpleistozän |
| Insel | Taxon | Autor | Datierung |
|---|---|---|---|
| Delos | Palaeoloxodon sp. | Vaufrey, 1929 | unklar |
| Naxos | Palaeoloxodon lomolinoi | Van der Geer et al., 2014[14] | Jungpleistozän |
| Paros | Elephantidae indet.A4 | Georgalas, 1929 | unklar |
| Kythnos | Elephantidae indet. | Honea, 1975 | Jungpleistozän? |
| Milos | Elephantidae indet. | Papp, 1953 | unklar |
| Serifos | Elephantidae indet. | Papp, 1953 | unklar |
| Insel | Taxon | Autor | Datierung |
|---|---|---|---|
| Kefalonia | Palaeoloxodon cephallonicusA5 | (Theodorou et al., 2018)[22] | Jungpleistozän |
Zypern
| Insel | Taxon | Autor | Datierung |
|---|---|---|---|
| Zypern | Palaeoloxodon cypriotes | (Bate, 1903)[32] | Jungpleistozän/Frühholozän? |
| Palaeoloxodon xylophagou | Athanassiou et al., 2015[20] | Mittelpleistozän |
Inselverbund[9]
Inselverbund[14]
Inselelefanten im Arktischen Ozean
Lange Zeit wurde angenommen, dass das Wollhaarmammut (Mammuthus primigenius) eine verzwergte Inselvariante auf der sibirischen Wrangelinsel im Arktischen Ozean ausgebildet hatte. Dort überlebte die letzte Population dieser Art noch bis vor etwa 3700 Jahren, also lange, nachdem das Mammut auf dem Festland ausgestorben war. Erste Berechnungen an Zahnfunden sprachen den Tieren eine Schulterhöhe von rund 1,8 m bei einem Gewicht von gut 2 t zu.[33] Analysen des Körperskeletts erbrachte keine Abweichungen von den spätpleistozänen Tieren des Festlandes, für die eine Körperhöhe von rund 2,2 m rekonstruierbar ist.[34] Auch für die Vertreter des Wollhaarmammuts auf den Pribilof-Inseln vor der Küste Alaskas, die ebenfalls erst im Holozän, aber etwas früher als die Mammuts der Wrangel-Insel ausstarben, wird eine frühe Phase der Inselverzwergung postuliert.[35]
Inselelefanten im Pazifischen Ozean
Auf den Kanalinseln vor der Küste Kaliforniens lebte im Jungpleistozän Mammuthus exilis. Diese Zwergform wurde zwischen 1,2 und 1,8 m groß und geht vermutlich auf das Präriemammut (Mammuthus columbi) des nordamerikanischen Festlandes zurück. Für die Zwergform wird ein Gewicht von rund 1,35 t rekonstruiert, die Ausgangsform hingegen wog zwischen 9,5 und 12,5 t. Beide Mammutarten starben am Ende des Pleistozäns aus.[36][5]
Zwergelefanten
Von Sulawesi, dem früheren Celebes, wurde eine Zwergelefantenart aus dem Alt- bis Mittelpleistozän Südostasiens berichtet. Ursprünglich als Elephas celebensis ausgewiesen, ordnen sie einige Wissenschaftler heute aufgrund des Zahnbaus zur eigenständigen Gattung Stegoloxodon. Zu dieser gehören zusätzlich einige Funde von Java.[37][38] Ein einziger Zahn von Luzon, der im Jahr 1956 als Elephas beyeri vorgestellt wurde,[39] ist in seinen tatsächlichen Verwandtschaftsverhältnissen unsicher und gehört entweder in die Linie des Asiatischen Elefanten oder zu Palaeoloxodon.[1]
Zwergstegodonten

Daneben sind auf verschiedenen Inseln der Region auch verzwergte Verwandte der Elefanten belegt, die zu den Stegodonten gezählt werden. Zeitgleich zu Stegoloxodon bewohnte so unter anderem Stegodon sompoensis die Insel Sulawesi. Die Form brachte wohl zwischen 350 und 950 kg auf die Waage.[37] Auf Flores erschien im ausgehenden Altpleistozän vor rund 900.000 Jahren Stegodon sondaari, das mit einem ein Gewicht von rund 300 kg der bislang kleinste Repräsentant der Stegodonten ist. Benannt wurde die Form im Jahr 1999 unter Berufung auf einen Unterkiefer und zugehörigen oberen Zähnen. Im Übergang zum Mittelpleistozän vor 850.000 bis 700.000 Jahren wurde die Art durch Stegodon florensis ersetzt. Diese Form war mit rund 850 kg merklich größer. Im weiteren Verlauf reduzierte sich ihre Größe aber offensichtlich wieder, da Funde aus dem Jungpleistozän auf rund 30 % kleinere Individuen verweisen. Sie starb dann aber in einem Zeitraum von vor 95.000 bis 12.000 Jahren aus.[37][40][41][7] Die südlich von Flores gelegene Insel Sumba barg gleichfalls die Reste eines extrem verzwergten Stegodonten, der als Stegodon sumbaensis bezeichnet wird. Es sind mehrere Unterkiefer dokumentiert worden, deren Altersstellung vergleichbar ist zu der von Stegodon sondaari.[42] Etwa zeitgleich zu den frühen Funden auf Flores trat auf Java mit Stegodon trigonocephalus eine verzwergte Form auf, die zwischen 1000 und 1700 kg schwer wurde. Von ihr ist ein recht umfangreiches Fossilinventar unter anderem von der bedeutenden Fundstelle von Trinil im Osten von Java überliefert. Es gehört der alt- bis mittelpleistozänen Kedung-Brubus-Fauna an. Die Art war dann noch in der nachfolgenden Ngandong-Fauna anwesend. Die Insel beherbergte aber verschiedene kleine Stegodontenformen, die wohl aufgrund der zerklüfteten Landschaft entstanden waren. So ist bei Sambungmacan, ebenfalls im östlichen Java gelegen, mit Stegodon hypsilophus eine weitere Art nachgewiesen, belegt über mehrere Zähne und Unterkiefer. Ihr Alter entspricht weitgehend den Funden von Trinil.[43][44][41] Eine weitere Zwergform bildete sich mit Stegodon timorensis auf Timor heraus. Erste Funde wurden bereits 1964 entdeckt,[45] die Art erhielt fünf Jahre später ihren Namen basierend auf einem einzelnen Zahn. Heute sind zahlreiche Funde von mehreren Fundstellen belegt, die auf ein Tier etwas größer als Stegodon sondaari schließen lassen. Das Alter der Funde liegt bei 168.000 bis 130.000 Jahren vor heute.[46][37] Von Mindanao stammen Zahnfunde eines kleinen Vertreters der Stegodonten, der im Jahr 1890 zur Art Stegodon mindanensis gestellt wurde.[47] Wiederum einzelne Zähne von Luzon werden mit der Zwergform Stegodon luzonensis verbunden.[39][37] Festlandsvertreter der Gattung Stegodon, die hochvariable Angehörige vereint, konnten ein Körpergewicht von bis zu 12,7 t erreichen.[5]
Übersicht der Nachweise auf südostasiatischen Inseln
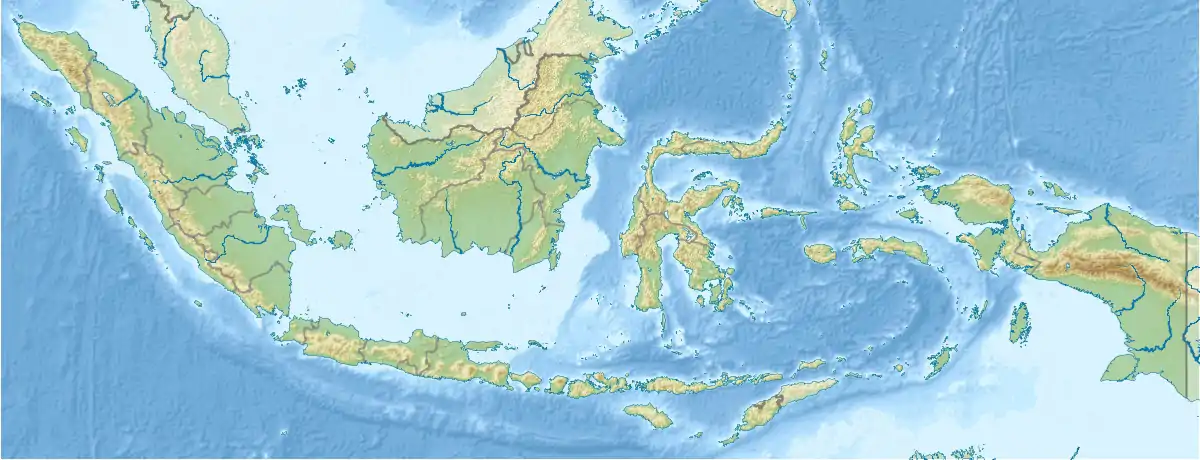 Zwergelefant (Indonesien)
| |||||
Inseln Südostasiens (Indonesien) mit Nachweisen von Zwergrüsseltieren der Gattung Stegodon ( |
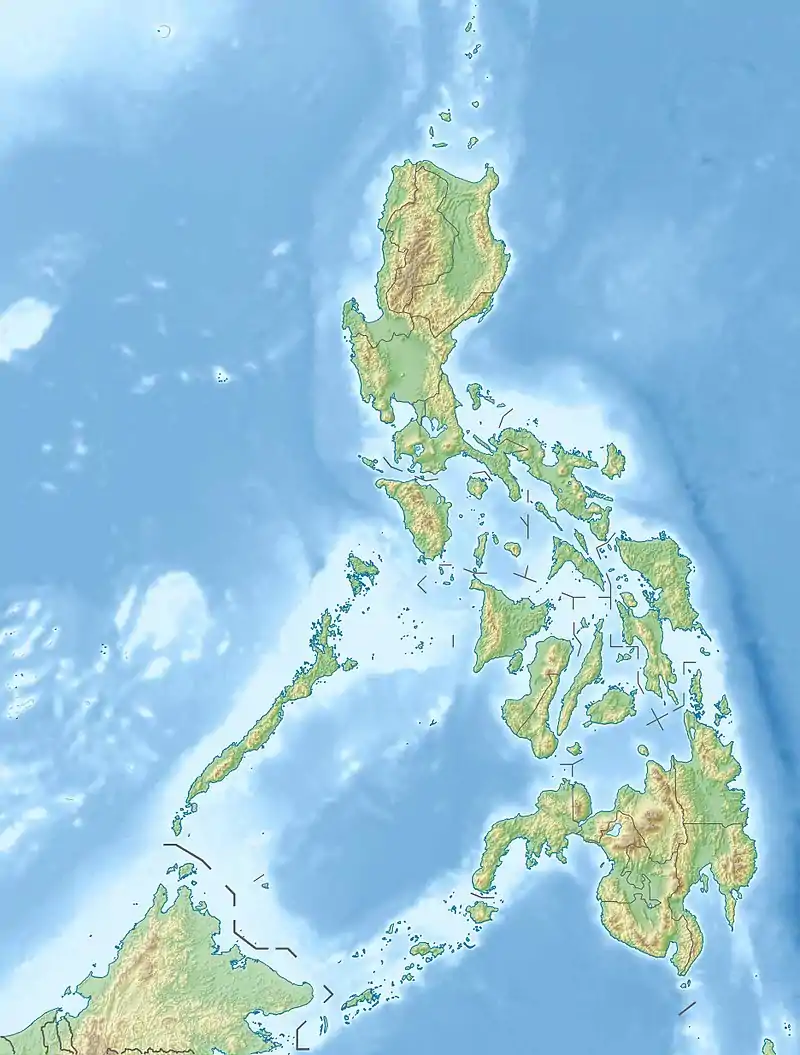 Zwergelefant (Philippinen)
| ||
Inseln Südostasiens (Philippinen) mit Nachweisen von Zwergrüsseltieren der Gattung Stegodon ( |
Indonesien
| Insel | Taxon | Autor | Datierung |
|---|---|---|---|
| Java | Stegodon hypsilophus | Hooijer, 1954[43] | Altpleistozän |
| Stegodon trigonocephalus | Martin, 1887[48] | Alt-/Mittelpleistozän | |
| Stegoloxodon indonesicus | Kretzoi, 1950[49] | Altpleistozän | |
| Sulawesi | Stegodon sompoensis | Hooijer, 1964[50] | Alt-/Mittelpleistozän |
| Stegoloxodon celebensisB1 | (Hooijer, 1949)[51] | Alt-/Mittelpleistozän |
| Insel | Taxon | Autor | Datierung |
|---|---|---|---|
| Flores | Stegodon florensis | Hooijer, 1957[52] | Mittel-/Jungpleistozän |
| Stegodon sondaari | van den Bergh, 1999[37] | Altpleistozän | |
| Stegodon cf. sondaari | van den Bergh, 1999[37] | Altpleistozän | |
| Sumba | Stegodon sumbaensis | Sartono, 1979 | Altpleistozän |
| Timor | Stegodon timorensis | Sartono, 1969 | Mittelpleistozän |
Philippinen
| Insel | Taxon | Autor | Datierung |
|---|---|---|---|
| Luzon | Stegodon luzonensis | von Koenigswald, 1956[39] | Pleistozän |
| Stegodon cf. trigonocephalus | von Koenigswald, 1956[39] | Pleistozän | |
| „Elephas“ beyeriB2 | von Koenigswald, 1956[39] | unklar | |
| Mindanao | Stegodon mindanensis | Naumann, 1890[47] | Pleistozän |
Heutige „Zwergelefanten“
„Borneo-Zwergelefant“
Der auf Borneo lebende „Borneo-Zwergelefant“ (Elephas maximus borneensis) ist eine angenommene Unterart des Asiatischen Elefanten (Elephas maximus). Ihr systematischer Status ist unsicher, ebenso wird diskutiert, ob es sich um eine endemische Population oder um vom Menschen eingeschleppte Tiere handelt. In der Körpergröße unterscheiden sie sich kaum von ihren Verwandten des südostasiatischen Festlandes.[53][54]
Afrikanischer „Zwergelefant“
Im Jahr 1906 beschrieb Theophil Noack ein kleines Individuum eines Afrikanischen Elefanten unter der wissenschaftlichen Bezeichnung Elephas africanus pumilio.[55] Das Tier lebte im Hamburger Tierpark, stammte aber ursprünglich aus dem Gebiet der heutigen Republik Kongo. Die geringe Größe des Tieres veranlasste Noack, seine neue Form als „Zwergelefanten“ anzusehen. Später wurde der „Zwergelefant“ in Loxodonta pumilio umbenannt. Über seine tatsächliche Existenz wurde lange Zeit eine kontroverse Debatte geführt. Befürwortern zufolge lebte der „Zwergelefant“ in den tropischen Regenwäldern des zentralen Afrikas und teilte sich dort seinen Lebensraum mit dem Waldelefanten (Loxodonta cyclotis).[56][57] Unterstützt wurde dies durch die gelegentliche Sichtung kleinerer Tiere mit ausgebildeten Stoßzähnen. Gegner dagegen lehnten seine Existenz ab und begründeten dies unter anderem damit, dass das Typusexemplar von Loxodonta pumilio zu seinen Lebenszeiten noch weiter gewachsen war und zu seinem Todeszeitpunkt eine Schulterhöhe von über 2 m aufwies. Sie sahen es als Jungtier des Waldelefanten an.[58] In der Wende von 20. zum 21. Jahrhundert erbrachten durchgeführte anatomische Vergleiche des Waldelefanten mit dem „Zwergelefanten“ keine Unterschiede zwischen den beiden Formen. Ebenso zeigten molekulargenetische Studien, dass der „Zwergelefant“ identisch mit dem Waldelefanten ist. Heute sehen Forscher die gelegentlichen Sichtungen kleinerer Elefanten im Kongobecken als verwaiste Jungtiere oder als pathologisch verzwergte Individuen des Waldelefanten an.[59][60]
Literatur
- Jordi Augusti: Mammoths, Sabertooths and Hominids 65 Million Years of Mammalian Evolution in Europe. Columbia University Press, New York NY u. a. 2002, ISBN 0-231-11640-3
- Erich Thenius: Grundzüge der Faunen- und Verbreitungsgeschichte der Säugetiere. Gustav Fischer, Stuttgart 1980, ISBN 3-437-30312-0
- Harald Gebhardt und Mario Ludwig: Von Drachen, Yetis und Vampiren – Fabeltieren auf der Spur. BLV-Verlag, München, 2005, ISBN 3-405-16679-9
Einzelnachweise
- Alexandra A. E. van der Geer, Gerrit D. van den Bergh, George A. Lyras, Unggul W. Prasetyo, Rokus Awe Due, Erick Setiyabudi und Hara Drinia: The effect of area and isolation on insular dwarf proboscideans. Journal of Biogeography 43 (8), 2016, S. 1656–1666, doi:doi:10.1111/jbi.12743
- Athanassios Athanassiou, Alexandra A. E. van der Geer und George A. Lyras: Pleistocene insular Proboscidea of the Eastern Mediterranean: A review and update. Quaternary Science Reviews 218, 2019, S. 306–321
- Sina Baleka, Victoria L. Herridge, Giulio Catalano, Adrian M. Lister, Marc R. Dickinson, Carolina Di Patti, Axel Barlow, Kirsty E.H. Penkman, Michael Hofreiter und Johanna L. A. Paijmans: Defining the island dwarfing rate of an extinct Sicilian elephant using ancient DNA. Current Biology, 2020, doi:10.2139/ssrn.35268913
- Per Christiansen: Body size in proboscideans, with notes on elephant metabolism. Zoological Journal of the Linnean Society 140, 2004, S. 523–549
- Asier Larramendi: Shoulder height, body mass, and shape of proboscideans. Acta Palaeontologica Polonica 61 (3), 2016, S. 537–574, doi:10.4202/app.00136.2014
- George Zammit Maempel: Għar Dalam. Cave and deposits. Malta, 1989
- Maria Rita Palombo: Elephants in miniature. In: Harald Meller (Hrsg.): Elefantenreich – Eine Fossilwelt in Europa. Halle/Saale, 2010, S. 275–295
- Lucia Caloi, Tassos Kotsakis, Maria Rita Palombo und Carmelio Petronio: The Pleistocene dwarf elephants of Mediterranean islands. In: Jeheskel Shoshani und Pascal Tassy (Hrsg.): The Proboscidea. Evolution and palaeoecology of the Elephants and their relatives. Oxford, New York, Tokyo, 1996, S. 234–239
- Maria Rita Palombo, Fabrizio Antonioli, Carolina Di Patti, Presti Lo Valeria und Matthew E. Acarborough: Was the dwarfed Palaeoloxodon from Favignana Island the last endemic Pleistocene elephant from the western Mediterranean islands? Historical Biology, 2020, S. 1–19, doi:10.1080/08912963.2020.1772251
- George Theodorou, Nikolaos Symeonidis und Elizabeth Stathopoulou: Elephas tiliensis n. sp. from Tilos (Dodecanese, Greece). Hellenic Journal of Geosciences 42, 2007, S. 19–32
- G. E. Theodorou: Die fossilen Zwergelefanten der Höhle „Charkadio“ auf der Insel Tilos, Athen 1983 (= Dissertation an der Dodekanes Univ.).
- Baruch Rosen: Mammoths in ancient Egypt? Nature 369, 1994, S. 364–365
- Hans Dietrich Kahlke: Das Eiszeitalter. Leipzig, Jena, Berlin, 1981
- Alexandra A. E. van der Geer, George A. Lyras, Lars W. van den Hoek Ostende, John de Vos und Hara Drinia: A dwarf elephant and a rock mouse on Naxos (Cyclades, Greece) with a revision of the palaeozoogeography of the Cycladic Islands (Greece) during the Pleistocene. Palaeogeography, Palaeoclimatology, Palaeoecology 404 (15), 2014, S. 133–144
- Sevket Sen, Eric Barrier und Xavier Crété: Late Pleistocene Dwarf Elephants from the Aegean Islands of Kassos and Dilos, Greece. Annales Zoologici Fennici 51 (1/2), 2014, S. 27–42
- Sevket Sen: A review of the Pleistocene dwarfed elephants from the Aegean islands, and their paleogeographic context. Fossil Imprint 73 (1/2), 2017; S. 76–92
- Rolfe D. Mandel und Alan H. Simmons: Geoarchaeology of the Akrotiri Aetokremnos Rockshelter, Southern Cyprus. Geoarchaeology: An International Journal 12 (6), 1997, S. 567–605
- Alan H. Simmons: Akrotiri-Aetokremnos (Cyprus) 20 years later: An Assessment of its significance. Eurasian Prehistory 10 (1–2), 2014, S. 139–156
- Antoine Zazzo, Matthieu Lebon, Anita Quiles, Ina Reiche und Jean-Denis Vigne: Direct Dating and Physico-Chemical Analyses Cast Doubts on the Coexistence of Humans and Dwarf Hippos in Cyprus. PLoS ONE 10 (8), 2015, S. e0134429, doi:10.1371/journal.pone.0134429
- Athanassios Athanassiou, Victoria Herridge, David S. Reese, George Iliopoulos, Socrates Roussiakis, Vassiliki Mitsopoulou, Efthymios Tsiolokis und George Theodorou: Cranial evidence for the presence of a second endemic elephant species on Cyprus. Quaternary International 379, 2015, S. 47–57
- N. K. Symeonides, G. E. Theodorou und V. I. Giannopoulos: New data on Elephas chaniensis (Vamos cave, Chania, Crete). In: G. Cavarretta, P. Gioia, M. Mussi und M. R. Palombo (Hrsg.): La terra degli Elefanti, The World of Elephants. Consiglio Nazionale delle Ricerche, Rom, 2001, S. 510–513
- George Theodorou, Yiannis Bassiakos, Evangelos Tsakalos, Evyenia Yiannouli und Petros Maniatis: The Use of CT Scans and 3D Modeling as a Powerful Tool to Assist Fossil Vertebrate Taxonomy. In: Marinos Ioannides, Eleanor Fink, Raffaella Brumana, Petros Patias, Anastasios Doulamis, João Martins und Manolis Wallace (Hrsg.): Digital heritage. Progress in cultural heritage: documentation, preservation, and protection. 7th International Conference, EuroMed 2018, Nicosia, Cyprus, October 29–November 3, 2018, Proceedings, Part I. Springer, 2018, S. 79–89
- R. Melis, Maria Rita Palombo und M. Mussi: Mammuthus lamarmorae (Major, 1883) remains in the pre-Tyrrhenian deposits of San Giovanni in Sinis (Western Sardinia, Italy). In: G. Cavarretta et al. (Hrsg.): The World of Elephants – International Congress. Consiglio Nazionale delle Ricerche. Rom, 2001, S. 481–485
- Nikos Poulakakis, Aris Parmakelis, Petros Lymberakis, Moysis Mylonas, Eleftherios Zouros, David S Reese, Scott Glaberman und Adalgisa Caccone: Ancient DNA forces reconsideration of evolutionary history of Mediterranean pygmy elephantids. Biology Letters 2 (3), 2006, S. 451–454, doi:10.1098/rsbl.2006.0467
- Victoria L. Herridge und Adrian M. Lister: Extreme insular dwarfism evolved in a mammoth. Proceedings of the Royal Society series B 279 (1741), S. 3193–3200, doi:10.1098/rspb.2012.0671
- Charles Immanuel Forsyth Major: Die Tyrrhenis: Studien über geographische Verbreitung von Tieren und Pflanzen im westlich Mittelmeergebiet. Kosmos 13, 1883, S. 1–17 ()
- George Busk: Description of the remains of three extinct species of elephant, collected by Capt. Spratt, C.B., R.N., in the ossiferous cavern of Zebbung, in the island of Malta; partly from the notes of the late H. Falconer. Transactions of the Zoological Society 6 (5), 1869; S. 227–306 (S. 251–271) ()
- A. Leith Adams: On the dentition and osteology of the Maltese fossil elephant, being a description of the remains discovered by the author in Malta between the years 1860 and 1866. Transactions of the Zoological Society of London 9 (1), 1874, S. 1–124 ()
- Dorothea M. A. Bate: On Elephant Remains from Crete, with Description of Elephas creticus, sp. n. Proceedings of the Zoological Society of London 77 (2), 1907, S. 238–250 ()
- Νικόλαος Κ. Συμεωνίδης, Γ. Θεοδώρου und Β. Γιαννόπουλος: Το νέο είδος Elephas chaniensis n. sp. στις υποθασάσσιες πλειστοκαινικές αποθέσεις του σπηλαίου Βάμου Χανίων Κρήτης. Δελτίον Ελληνικής Σπηλαιολογικής Εταιρίας 22, 2000, S. 95–108 ()
- Siegfried E. Kuss: Eine Pleistozäne Säugetierfauna der Insel Kreta. Berichte der Naturforschenden Gesellschaft zu Freiburg im Breisgau 55, 1965, S. 271–348 (S. 287–308)
- Dorothea M. A. Bate: Preliminary note on the discovery of a pigmy elephant in the Pleistocene of Cyprus. Proceedings of the Royal Society of London 71, 1903, S. 498–500 ()
- Adrian M. Lister: Mammoths in miniature. Nature 362, 1993, S. 288–289
- Sergey L. Vartanyan, Khikmat A. Arslanov, Juha A. Karhu, Göran Possnert und Leopold D. Sulerzhitsky: Collection of radiocarbon dates on the mammoths (Mammuthus primigenius) and other genera of Wrangel Island, northeast Siberia, Russia. Quaternary Research 70, 2008, S. 51–59, doi:10.1016/j.yqres.2008.03.005
- J. M. Enk, D. R. Yesner, K. J. Crossen, D. W. Veltre und D. H. O’Rourke: Phylogeographic Analysis of the mid-Holocene Mammoth from Qagnax Cave, St. Paul Island, Alaska. Palaeogeography, Palaeoclimatology, Palaeoecology 273 (1–2), 2009, S. 184–190
- V. Louise Roth: Pleistocene dwarf elephants from the California Islands. In: Jeheskel Shoshani und Pascal Tassy (Hrsg.): The Proboscidea. Evolution and palaeoecology of the Elephants and their relatives. Oxford, New York, Tokyo, 1996, S. 249–253
- Gerrit D. Van Den Bergh: The Late Neogene elephantoid-bearing faunas of Indonesia and their palaeozoogeographic implications; a study of the terrestrial faunal succession of Sulawesi, Flores and Java, including evidence for early hominid dispersal east of Wallace’s Line. Scripta Geologica 117, 1999, S. 1–419
- Georgi N. Markov und Haruo Saegusa: On the validity of Stegoloxodon Kretzoi, 1950 (Mammalia: Proboscidea). Zootaxa 1861, 2008, S. 55–56
- G. H. R. von Koenigswald: Fossil mammals from the Philippines. Proceedings of the Fourth Far-Eastern Prehistory and the Anthropology Division of the 8th Pacific Science Congresses, Quezon City, 1956, S. 339–369
- Gerrit D. van den Bergh, John de Vos, F. Aziz und M. J. Morwood: Elephantoidea in the Indonesian region: new Stegodon findings from Flores. In: G. Cavarretta, P. Gioia, M. Mussi und M. R. Palombo (Hrsg.): La terra degli Elefanti, The World of Elephants. Consiglio Nazionale delle Ricerche, Rom, 2001, S. 623–627
- Gerrit D. Van Den Bergh, Rokhus Due Awe, M. J. Morwood, T. Sutikna, Jatmiko und E. Wahyu Saptomo: The youngest stegodon remains in Southeast Asia from the Late Pleistocene archaeological site Liang Bua, Flores, Indonesia. Quaternary International 182, 2008, S. 16–48, doi:10.1016/j.quaint.2007.02.001
- Samuel T. Turvey, Jennifer J. Crees, James Hansford, Timothy E. Jeffree, Nick Crumpton, Iwan Kurniawan, Erick Setiyabudi, Thomas Guillerme, Umbu Paranggarimu, Anthony Dosseto und Gerrit D. van den Bergh: Quaternary vertebrate faunas from Sumba, Indonesia: implications for Wallacean biogeography and evolution. Proceedings of the Royal Society B 284, 2017, S. 20171278, doi:10.1098/rspb.2017.1278
- D. A. Hooijer: A Pygmy Stegodon from the Middle Pleistocene of Eastern Java. Zoologische Mededelingen 33, 1954, S. 91–102 ()
- Fachroel Aziz und Gerrit D. van den Bergh: A dwarf stegodon from Sambungmacan (Central Java, Indonesia). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen 98 (3), 1995, S. 229–241
- Th. Verhoeven: Stegodon-Fossilien auf der Insel Timor. Anthropos 59 (3./4), 1964, S. 634
- Julien Louys, Gilbert J. Price und Sue O'Connor: Direct dating of Pleistocene stegodon from Timor Island, East Nusa Tenggara. PeerJ 4, 2016, S. e1788, doi:10.7717/peerj.1788
- Edmund Naumann: Stegodon mindanensis, eine Art von Uebergangs-Mastodonten. Zeitschrift der Deutschen Geologischen Gesellschaft 42, 1890, S. 166–169 ()
- K. Martin: Fossile Saugethierreste von Java und Japan. Jaarboek van het mijnwezen in Nederlandsch Oost-Indië 16 (1), 1887, S. 25–69
- Miklós Kretzoi: Stegeloxodon nov. gen., a loxodonta elefántok esetleges ázsiai öse (Stegoloxodon nov. gen., a possible Asiatic ancestor of true loxodonts). Földtani Közlöny, 80, 1950, S. 405–408 ()
- D. A. Hooijer: Pleistocene Vertebrates from Celebes. XII. Notes on pygmy Stegodonts. Zoologische Mededelingen 40, 1964, S. 37–44 ()
- D. A. Hooijer: Pleistocene Vertebrates from Celebes. IV. Archidiskodon celebensis nov.spec. Zoologische Mededelingen 30, 1949, S. 205–226 ()
- D. A. Hooijer: A Stegodon from Flores. Treubia 24, 1957, S. 119–129
- Earl of Cranbrook, J. Payne und Charles M. U. Leh: Origin of the elephants Elephas maximus L. of Borneo. Sarawak Museum Journal 63, 2007, S. 95–125
- Reeta Sharma, Benoit Goossens, Rasmus Heller, Rita Rasteiro, Nurzhafarina Othman, Michael W. Bruford und Lounès Chikhi: Genetic analyses favour an ancient and natural origin of elephants on Borneo. Scientific Reports 8, 2018, S. 880, doi:10.1038/s41598-017-17042-5
- Theophil Noack: A dwarf form of the African elephant. Annals and Magazine of Natural History 17, 1906, S. 501–503 ()
- L. Blancou: A propos des formes naines de l’éléphant d’Afrique. Mammalia 26, 1962, S. 343–361
- E. Bourdelle und F. Petter: Note relative à un éléphant nain du Gabon. Mammalia 13, 1950, S. 144–153
- Pierre Pfeffer: Sur la validité de formes naines de l’éléphant d’Afrique. Mammalia 24, 1960, S. 556–576
- Colin P. Groves und Peter Grubb: Are there pygmy elephants?. Elephant 2 (4), 2000, S. 8–10
- Régis Debruyne, Arnaud Van Holt, Véronique Barriel und Pascal Tassy: Status of the so-called African pygmy elephant (Loxodonta pumilio (NOACK 1906)): phylogeny of cytochrome b and mitochondrial control region sequences. Comptes Rendus Biologies 326, 2003, S. 687–697