Tiefsee-Anglerfische
Die Tiefsee-Anglerfische oder Tiefseeangler (Ceratioidei) sind eine Unterordnung von Tiefseefischen und mit etwa 160 Arten das artenreichste Taxon der Wirbeltiere im Bathypelagial (1000 bis 4000 Metern Tiefe).
| Tiefsee-Anglerfische | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
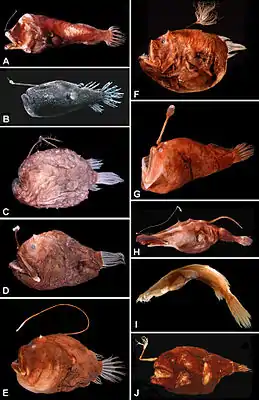
Tiefsee-Anglerfische aus verschiedenen Familien: | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Ceratioidei | ||||||||||||

Sie kommen in allen Weltmeeren unterhalb von 300 Meter Tiefe vor. Die Tiefsee-Anglerfische bestehen aus elf verschiedenen Familien und werden zur Ordnung der Armflosser (Lophiiformes) gezählt. Im Unterschied zu allen anderen Armflossern, die bodenbewohnend sind, leben die Tiefsee-Anglerfische pelagisch.
Abgesehen von morphologischen Untersuchungen an Totfängen sind die Tiefsee-Anglerfische wenig erforscht. Beobachtungen an lebenden Fischen sind selten.
Verbreitung
Tiefsee-Anglerfische kommen in der Tiefsee aller Ozeane vor, vom Arktischen Ozean bis zum Südlichen Ozean. Einige Arten haben eine weltweite Verbreitung, während das Verbreitungsgebiet anderer sehr klein und räumlich sowie auch vertikal sehr beschränkt ist. Im Meso- (200 bis 1000 Meter Tiefe) und Bathypelagial (1000 bis 4000 Meter) sind sie die Hauptcarnivoren und ökologisch wichtig.
Merkmale
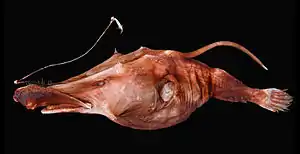
Tiefsee-Anglerfische sind plumpe Fische mit aufgedunsenen Körpern, großen Köpfen und großen, mit langen Fangzähnen besetzten Mäulern. Der Körper ist in den meisten Fällen schuppenlos, die Haut kann aber mit Papillen oder Stacheln bedeckt sein. Weibchen erreichen Körperlängen zwischen 6 cm und 1,2 m. Tiefsee-Anglerfische sind normalerweise dunkelbraun bis schwärzlich gefärbt, bei der Gattung Haplophryne aus der Familie der Linophrynidae ist die Haut unpigmentiert und transparent. Das Palatinum und das Basihyale, ein Element zwischen den Zungenbeinen, fehlen bei ausgewachsenen Weibchen. Auch die Pseudobranchie fehlt, Überreste sind nur noch bei einigen Gigantactis-Arten vorhanden. Das Supraoccipitale, ein Schädelknochen, liegt mittig hinter den Frontalia und zwischen den Parietalia. Die Frontalia sind nicht zusammengewachsen. Die unteren Pharyngealia sind reduziert und zahnlos. Tiefsee-Anglerfische besitzen 19 bis 24 Wirbel.
Tiefsee-Anglerfische zeigen einen extremen Sexualdimorphismus und unterscheiden sich damit von ihren benthischen (bodenbewohnenden) Verwandten aus der Ordnung der Armflosser. Die winzigen Männchen (Zwergmännchen), die man zunächst für Larven oder Jungfische hielt, erreichen oft nur 5 bis 10 Prozent der Weibchengröße. Bei Ceratias holboelli sind die Weibchen sechzig Mal größer als die Männchen und mit einer Länge von maximal 1,20 Metern die größten Tiefseeangler. Weibchen besitzen eine „Angel“ (Illicium) mit anhängendem „Köder“ (Esca), der üblicherweise mit einem Leuchtorgan ausgestattet ist. Bei Weibchen können Leuchtorgane auch an anderen Stellen vorhanden sein. Eine Schwimmblase fehlt.

Die Flossenformel ist für männliche und weibliche Tiefsee-Anglerfische gleich. Innerhalb der Überfamilie ist sie sehr variabel. Für die meisten Familien gilt:
Dorsale 3–8, Anale 3–7, Pectorale 12–30, Ventrale 2–5, Caudale 9.
Davon abweichend haben die Neoceratiidae 11 bis 13 Flossenstrahlen in der Rückenflosse und 10 bis 13 in der Afterflosse. Bei den Melanocetidae sind es 12 bis 22 Flossenstrahlen in der Rückenflosse, bei den Caulophrynidae ebenfalls, sowie 5 bis 19 in der Afterflosse. Ctenochirichthys hat 28 bis 30 Brustflossenstrahlen. Die Schwanzflosse hat 8 Flossenstrahlen bei Cryptopsaras und 10 bei den Neoceratiidae. Sie ist bei fast allen Familien abgerundet, bei großen Individuen der Ceratiidae endet sie in runden, mit Haut bedeckten Verknöcherungen. Bauchflossen fehlen bei ausgewachsenen Tiefsee-Anglerfischen und sind nur noch bei den Larven und kurz nach der Metamorphose bei den Caulophrynidae vorhanden. Beckenknochen fehlen oder sind reduziert. Die Flossenstrahlen der unpaaren Flossen sind manchmal stark verlängert, vor allem bei den Weibchen von Caulophrynidae und Gigantactinidae, und oft nicht durch Flossenmembran verbunden, oder die Flossenmembran ist extrem dünn und transparent.
Alle Flossen werden nur von Weichstrahlen gestützt, eine von Flossenstacheln gestützte Rückenflosse fehlt. Der erste Rückenflossenstachel ist bei den Weibchen zu einer „Angel“ (Illicium) mit anhängendem „Köder“ (Esca) umgewandelt und befindet sich auf der Oberseite des Kopfes. Der zweite ist bis auf einen winzigen Rest reduziert und von Haut überwachsen, teilweise auch mit der Oberseite des Flossenträgers verwachsen und unmittelbar hinter dem Illicium liegend. Beide Rückenflossenstacheln werden von einem einzigen Flossenträger gestützt.
Weibchen
Der Körper der Weibchen ist meist rundlich und kurz, bei den Thaumatichthyidae, Centrophrynidae, Ceratiidae, Gigantactinidae, Neoceratiidae und einigen Oneirodidae aber langgestreckt und seitlich abgeflacht. Der Kopf nimmt normalerweise mehr als 40 Prozent der Standardlänge ein, aber nur ein Viertel bei den Arten mit langgestrecktem Körper. Wie bei anderen Armflossern ist das Maul extrem groß. Die Länge der Prämaxillare reicht von 10 Prozent der Standardlänge bei einigen Gigantactinidae bis zu 40 Prozent bei einigen Linophrynidae. Die Maulspalte steht fast horizontal bei Thaumatichthyidae, Centrophrynidae und Gigantactinidae, fast vertikal bei Melanocetidae und Ceratiidae. Kiemenreusenstrahlen sind auf den Kiemenbögen II und III als Holobranchien (Kiemenbögen, die sowohl an der vorderen als auch der hinteren Seite respiratorische Filamente tragen) vorhanden und reichen bei einigen Familien (Caulophrynidae, Himantolophidae, Melanocetidae, Diceratiidae, einige Oneirodidae, Centrophrynidae und Neoceratiidae) als Hemibranchien (respiratorische Filamente nur an einer Seite) auch auf Kiemenbogen I und IV.
Männchen
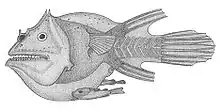
Männchen werden in den meisten Fällen nur wenige Zentimeter lang. Zwergmännchen aus der Familie Linophrynidae erreichen nur eine Länge von 6 bis 10 mm und zählen damit zu den kleinsten Wirbeltieren überhaupt. Sie sind noch kleiner als die Karpfenartigen der Gattung Paedocypris und die Grundelartigen der Gattung Schindleria. Der Körper der Tiefseeanglermännchen ist langgestreckt, der Kopf kürzer als 40 Prozent der Standardlänge, die Länge der Prämaxillare macht weniger als 15 Prozent der Standardlänge aus. Das Maul steht horizontal. Die kopfständigen Rückenflossenstacheln, die bei den Weibchen die Esca bilden, sind stark reduziert, die Überreste sind von Haut bedeckt; Leuchtorgane fehlen. Augen und Geruchsorgan sind bei den meisten Arten stark vergrößert. Letztere dienen dazu, die artspezifischen Pheromone aufzuspüren, die von den Weibchen abgegeben werden. Die normale Bezahnung geht während der Metamorphose verloren und wird durch hakenförmige Zähne in der Schnauzenspitze ersetzt, die dazu dienen, sich in der Haut der Weibchen festzubeißen.
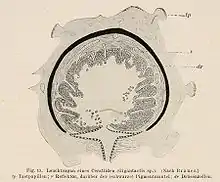
Leuchtorgane
Weibliche Tiefsee-Anglerfische unterscheiden sich von ihren in flacheren Meeresregionen lebenden Verwandten z. B. den vor allem in Korallenriffen lebenden Anglerfischen (Antennariidae), durch den Besitz eines Leuchtorgans an der als Beuteattrappe fungierenden Esca. Nur die Weibchen der Familie Caulophrynidae, der Gigantactiniden Gattung Rhynchactis und die von Neoceratias spinifer sind ohne Leuchtorgane. Form, Größe und Anzahl der Anhänge und Filamente der Esca sind ein wichtiges Merkmal, um die verschiedenen Arten voneinander zu unterscheiden.
Die innere Struktur der Esca ist kompliziert und beinhaltet eine verwirrende Vielfalt von Bläschen, die mit lumineszierenden Bakterien gefüllt sind, von Licht absorbierenden Schichten, Pigmenten, reflektierendem Gewebe, röhrenförmigen, lichtleitenden Strukturen, Nerven, Blutgefäßen und Muskelfasern. Es spricht einiges dafür, dass die Esca auch Pheromon-produzierende Drüsen enthält, die dazu dienen, Männchen anzulocken.
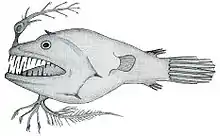
Neben dem Leuchtorgan der Esca besitzen alle Arten der Gattung Linophryne (Linophrynidae) eine kompliziert gebaute Bartel am Zungenbein (Hyoid). Das Licht der Bartel stammt nicht von lichtproduzierenden Bakterien, sondern von einem zwischen den Zellen liegenden, fotogenen Granulat. Das Leuchtorgan der Hyoidbartel hat sich aus dem Mesoderm entwickelt, während das Licht der Esca praktisch außerhalb des Körpers, von Bakterien produziert wird. Die unabhängige Entwicklung von zwei verschiedenen lichtproduzierenden Mechanismen bei der Gattung Linophryne ist einzigartig in der Tierwelt.
„Sexualparasitismus“

In einigen Familien und Gattungen – bei den Caulophrynidae, Ceratiidae, Neoceratiidae, Linophrynidae und den Oneiroden-Gattungen Bertella und Leptacanthichthys – wachsen Haut und Blutkreislauf der Partner zusammen; die Männchen leben fortan fest mit den Weibchen verbunden.
Das dauerhafte Anwachsen erfolgt durch zwei getrennte Auswüchse von der Spitze des Ober- und Unterkiefers des Männchens, die beide schließlich mit der Haut des Weibchens verschmelzen. Bei einigen Arten wächst die Haut des Weibchens in das männliche Maul und kleidet schließlich den gesamten Schlund aus. So entsteht ein länglicher, zylindrischer Stiel, mit dem das Männchen an das weibliche Gewebe festgewachsen ist. Die Köpfe der Männchen können bei anderen Arten auch großflächig – von der Unterkieferspitze bis zum Hinterschädel – mit der Haut der Weibchen verschmelzen. Sie sind von da an völlig unfähig, sich selbständig zu ernähren, und werden ähnlich wie Embryonen in der Gebärmutter der Säugetiere durch den Blutkreislauf des Weibchens ernährt. Angewachsene Zwergmännchen legen noch deutlich an Größe zu, auch verglichen mit der Länge frei lebender Männchen der gleichen Art. Sie laichen mit den Weibchen und sterben mit dem Tod der Weibchen. Bei vielen Arten ist ein angewachsenes Zwergmännchen pro Weibchen die Normalität, bei anderen können es mehrere sein. Der Rekord beobachteter Exemplare liegt bei acht Zwergmännchen an einem Weibchen.
Das Anwachsen ist seit den 1920er-Jahren bekannt und wird in vielen angelsächsischen Veröffentlichungen als Sexualparasitismus bezeichnet, hat jedoch mit der Definition von Parasitismus nichts zu tun. Die physiologischen Vorgänge, die den Vorgang erlauben, sind bisher nicht erforscht worden und könnten für Biomedizin, Immunologie und Endokrinologie von Bedeutung sein.[1] Laut aktueller Forschung ist die Eiweißabwehr bei den Fischen mit Sexualparasitismus nicht ausgeprägt.[2]
Äußere Systematik

Von den fünf Haupttaxa der Armflosser sind die Tiefsee-Anglerfische phylogenetisch am stärksten abgeleitet. Die Verwandtschaftsverhältnisse nach einer aktuellen Studie zeigt folgendes Kladogramm:[3]
| Armflosser (Lophiiformes) |
| ||||||||||||||||||||||||
Innere Systematik

Die Unterordnung der Tiefsee-Anglerfische beinhaltet etwa 160 Arten aus elf Familien. Folgendes Kladogramm zeigt alle Familien und die wahrscheinlichen Verwandtschaftsverhältnisse zueinander:[4]
| Ceratioidei |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sonstiges
Der im Golf von Mexiko entdeckte Lasiognathus dinema wurde als hässlichste neue Art von 18.000 im Jahr 2015 erstmals beschriebenen Arten bezeichnet.[5]
Quellen
Die Informationen dieses Artikels entstammen zum größten Teil der unter Weblinks angegebenen Seite des Anglerfischexperten Theodore W. Pietsch beim Tree of Life Web Project. Darüber hinaus wurden folgende Quellen benutzt:
Literatur
- Kurt Fiedler: Fische (= Lehrbuch der Speziellen Zoologie. Bd. 2: Wirbeltiere. Tl. 2). Gustav Fischer, Jena 1991, ISBN 3-334-00338-8.
- Joseph S. Nelson: Fishes of the World. 4th edition. John Wiley & Sons, Hoboken NJ u. a. 2006, ISBN 0-471-25031-7.
- E. O. Wiley, G. David Johnson: A teleost classification based on monophyletic groups. in Joseph S. Nelson, Hans-Peter Schultze, Mark V. H. Wilson: Origin and Phylogenetic Interrelationships of Teleosts. Verlag Dr. Friedrich Pfeil, München 2010, ISBN 978-3-89937-107-9
Weblinks
- Theodore W. Pietsch & Christopher P. Kenaley: Ceratioidei. Seadevils, Devilfishes, Deep-sea Anglerfishes, 2007 (The Tree of Life Web Project)
- Hildegard Kaulen: Immunsystem von Tiefseefischen: Bis dass der Tod sie scheidet. In: FAZ.net. 8. August 2020.
Einzelnachweise
- M. P. Black, M. S. Grober: Group sex, sex change, and parasitic males: sexual strategies among the fishes and their neurobiological correlates. In: Annual Review of Sex Research. Band 14, Nr. 1, 2003, S. 160–184 (online [PDF; 3,4 MB]).
- Sexualparasitismus mit medizinischer Relevanz - wissenschaft.de. In: wissenschaft.de. 30. Juli 2020 (wissenschaft.de [abgerufen am 2. August 2020]).
- Ricardo Betancur-R, Edward O. Wiley, Gloria Arratia, Arturo Acero, Nicolas Bailly, Masaki Miya, Guillaume Lecointre und Guillermo Ortí: Phylogenetic classification of bony fishes. BMC Evolutionary Biology, BMC series – Juli 2017, doi:10.1186/s12862-017-0958-3
- Masaki Miya, Theodore W. Pietsch, James W. Orr, Rachel J. Arnold, Takashi P. Satoh, Andrew M. Shedlock, Hsuan-Ching Ho, Mitsuomi Shimazaki, Mamoru Yabe, Mutsumi Nishida: Evolutionary history of anglerfishes (Teleostei: Lophiiformes): a mitogenomic perspective. In: BMC Evolutionary Biology, Band 10, Nr. 1, Feb 2010, S. 58. doi:10.1186/1471-2148-10-58.
- http://science.orf.at/stories/2775782/ Schaurig, schön und skurril, orf.at, 23. Mai 2016, abgerufen 23. Mai 2016.