Tapirus augustus
Tapirus (Megatapirus) augustus ist ein ausgestorbener, sehr großer Vertreter der Familie der Tapire, der innerhalb der Gattung Tapirus als Untergattung Megatapirus geführt wird. Fossilien dieses Unpaarhufers, die mehrere vollständige Schädel, weitere Gebissfragmente und Reste des Körpers umfassen, stammen aus dem Pleistozän und wurden weitgehend in Ost- und Südostasien (China, Vietnam) nachgewiesen.
| Tapirus augustus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Schädel von Tapirus augustus | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Pleistozän | ||||||||||||
| 1,8 Mio. Jahre bis 10.000 Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Untergattung | ||||||||||||
| Megatapirus | ||||||||||||
| Matthew & Granger, 1923 | ||||||||||||
| Wissenschaftlicher Name der Art | ||||||||||||
| Tapirus augustus | ||||||||||||
| Matthew & Granger, 1923 |
Merkmale
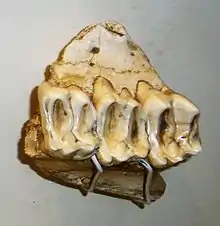
Megatapirus stellte den größten bekannten Vertreter der Tapire dar und besaß Rekonstruktionen zufolge etwa eine Kopf-Rumpf-Länge von 3,5 m und eine Schulterhöhe von 1,5 m. Megatapirus übertraf dabei die größte heute lebende Tapirart, den Schabrackentapir, in dessen Maßen jeweils um 25 %.[1][2][3] Im Skelettbau, vor allem im Postcranium ähnelte Megatapirus weitgehend den anderen Tapiren, zeichnete sich jedoch durch seine allgemeine Größe aus.[4] Der Schädel wurde bis zu 53 cm lang,[2] war dabei aber vergleichsweise kürzer als bei den anderen Tapirarten, und wies in der Seitenansicht einen gewölbten Stirnbereich auf. Wie alle Tapire besaß er einen kräftig ausgebildeten Scheitelkamm. Das Hinterhauptsbein wies eine nur kurze und rechtwinklige Form auf., Das Nasenbein lag wie bei allen Tapiren weit hinter dem Mittelkieferknochen und zeigte eine nur schwache Bildung.[5]
Das Gebiss umfasste im Oberkiefer die vollständige ursprüngliche Bezahnung der Säugetiere mit drei Schneidezähnen, einem Eckzahn, vier Prämolaren und drei Molaren, im Unterkiefer war der erste Prämolar nicht ausgebildet. Die Zahnformel für das Dauergebiss lautete somit: . Im Oberkiefer war der jeweils dritte Schneidezahn vergrößert ausgebildet und wies eine dolchartige Form auf. Der folgende Eckzahn, zu dem ein kleines Diastema bestand, hatte dagegen eine nur sehr kleine Form. Im Unterkiefer war die Situation genau umgekehrt mit einem großen Eckzahn und einem kleinen dritten Schneidezahn. Zu den nachfolgenden Backenzähnen bestand jeweils ein wesentlich größeres Diastema.[6][4] Die Prämolaren zeichneten sich durch eine starke Molarisierung aus, viel stärker als bei den anderen Tapirarten, und ähnelten damit deutlich den hinteren Backenzähnen. Allgemein waren die Backenzähne durch zwei quer verlaufende Zahnschmelzleisten (bilophodont) gekennzeichnet, während die Zahnkronen eine relativ niedrige Höhe besaßen (brachydont).[3]

Verbreitungsgebiet
Megatapirus war weitgehend in Ost- und Südostasien verbreitet. In China ist es überwiegend in den südlichen Landesteilen nachgewiesen, wo allein mehr als 20 Fundstellen bekannt sind, vereinzelt ist es auch im nördlichen China während warmklimatischer Phasen im Pleistozän nachgewiesen.[7] Bedeutende Funde stammen aus Daxin aus dem Mittelpleistozän, während die Reste aus Liujiang aus dem Jungpleistozän stammen und mit Fossilien eines frühen Menschen (Liujiang-Mensch) vergesellschaftet waren. Beide Fundplätze liegen im Autonomen Gebiet Guangxi. Weiterhin sind Reste von Megatapirus in Vietnam und Laos gefunden worden. Hier ist vor allem die Höhle von Tham Khuyen in Nordvietnam erwähnenswert, wo die Tapirart gemeinsam mit dem Riesenaffen Gigantopithecus und dem Homo erectus vorkam.[8][3]
Paläobiologie
Die deutlich bilophodont und niederkronig aufgebauten Backenzähne von Megatapirus sprechen zusammen mit charakteristischen Schliffmustern an den querverlaufenden Schmelzleisten für eine, den heutigen Tapiren ähnliche Ernährung durch weiche Pflanzenkost, wie Blätter, Zweige oder Früchte. Der Lebensraum umfasste weitgehend tropische Regenwälder, das gelegentlich weiter nördliche Auftreten könnte auch für eine Akzeptanz stärker saisonalisierter, aber immer noch warmer Klimate sprechen. Einige Forscher vertreten die Ansicht, dass Megatapirus eine ähnliche ökologische Nische wie die Flusspferde besetzte, die in China nicht sicher nachgewiesen sind.[7] In südlicheren Teilen Asiens, etwa in Thailand ist die Flusspferd-Gattung Hexaprotodon dagegen im Pleistozän bekannt.[9]
Systematik
Die Familie der Tapire (Tapiridae) ist ein relativ alter Vertreter der Säugetiere, die älteste bekannte Gattung ist Protapirus, die im Oligozän lebte. Die nächsten lebenden Verwandten der Tapire sind die Nashörner, die beiden Gruppen trennten sich aber molekulargenetischen Analysen zufolge bereits vor rund 47 Millionen Jahren.[10] Zusammen mit diesen und den ausgestorbenen Hyracodontidae und Amynodontidae formen sie die Unterordnung der Ceratomorpha.[5]
Innerhalb der Tapire entwickelten sich, auch aufgrund der Trennung der asiatischen und amerikanischen Vertreter, mehrere Linien. Im Pleistozän Ostasiens entstanden unter anderem zwei parallele Entwicklungslinien. Megatapirus entwickelte sich dabei über T. peii und T. sinensis und steht somit der Entwicklungsabfolge von T. sanyuanensis zu T. indicus (Schabrackentapir) gegenüber. Beide Linien zeichnen sich durch die Zunahme der Körpergröße aus, Unterschiede bestehen vor allem im Aufbau und in der Ausprägung der Backenzähne.[3] Megatapirus selbst formte sich in einer späteren Phase des Altpleistozäns und ist überwiegend aus dem Mittel- und Jungpleistozän bekannt. Gegen Ende der letzten Eiszeit starb der Tapirvertreter weitgehend aus, von einigen chinesischen Fundorten wie Chonqing und Zheijiang liegen Daten vor, die annehmen lassen, einige Restpopulationen hätten bis ins mittlere Holozän überlebt.[8]
Forschungsgeschichte
Tapirus augustus wurde 1923 von William Diller Matthew und Walter W. Granger anhand mehrerer aus dem Altpleistozän stammender Schädelfunde (Holotyp-Nummer AMNH 18433) aus der chinesischen Provinz Sichuan wissenschaftlich erstbeschrieben. Im gleichen Zug führten beide Autoren die Bezeichnung Megatapirus ein und wiesen auf den Status als Untergattung innerhalb der Gattung Tapirus hin.[2][11] In einer Studie von 1953 wurde Megatapirus als eigenständige Gattung eingeführt und angenommen, dass diese sich möglicherweise in Ostasien aus der Form Tapirus entwickelte,[4] was später mehrfach wiederholt ausgedrückt wurde.[3] Aufgrund des aber dadurch entstehenden paraphyletischen Ursprungs der Gattung Tapirus wurde Megatapirus 1998 mit Tapirus wieder gleichgesetzt und die Form als Untergattung geführt. Die einzige bekannte Art ist Tapirus (Megatapirus) augustus.[6][11]
Literatur
- Malcolm C. McKenna und Susan K. Bell: Classification of Mammals
- Spencer G. Lucas: Chinese Fossil Vertebrates
- Edwin H. Colbert, Michael Moral und Eli C. Minkoff: Colbert's Evolution of the Vertebrates: A History of the Backboned Animals Through Time
Einzelnachweise
- Darren Naish: The biggest tapir. 2009 ()
- William Diller Matthew und Walter Granger: New fossil mammals from the Pliocene of Sze-chuan, China. Bulletin of the American Museum of Natural History 48, 1923, S. 563–598
- Tong Haowen: Dental characters of the Quaternary tapirs in China, their significance in classification and phylogenetic assessment. Geobios 38, 2005, S. 139–150
- Edwin Harris Colbert und Dirk Albert Hooijer: Pleistocene mammals from the limestone fissures of Szechwan, China. Bulletin of the American Museum of Natural History 102, 1953, S. 1–134
- Luke T. Holbrook: Comparative osteology of early Tertiary tapiromorphs (Mammalia, Perissodactyla). Zoological Journal of the Linnean Society 132, 2001, S. 1–54
- Richard C. Hulbert Jr.: Late Miocene Tapirus (Mammalia, Perissodactyla) from Florida, with description of new species Tapirus webbi. Bulletin of the Florida. Museum of Natural History 45 (4), 2005, S. 465–494
- Tong Haowen: Occurrences of warm-adapted mammals in north China over the Quaternary Period and their paleoenvironmental significance. Science in China, Series D: Earth Sciences 50 (9), 2007, S. 1327–1340
- Julien Louys, Darren Curnoe und Haowen Tong: Characteristics of Pleistocene megafauna extinctions in Southeast Asia. Palaeogeography, Palaeoclimatology, Palaeoecology 243, 2007, S. 152–173
- C. Tougard: Biogeography and migration routes of large mammal faunas in South–East Asia during the Late Middle Pleistocene: focus on the fossil and extant faunas from Thailand. Palaeogeography, Palaeoclimatology, Palaeoecology 168 (3–4), 2001, S. 337–358
- Christelle Tougard, Thomas Delefosse, Catherine Hänni und Claudine Montgelard: Phylogenetic Relationships of the Five Extant Rhinoceros Species (Rhinocerotidae, Perissodactyla) Based on Mitochondrial Cytochrome b and 12S rRNA Genes. Molecular Phylogenetics and Evolution 19, 2001, S. 34–44
- Richard C. Hulbert Jr.: A new Early Pleistocene tapir (Mammalia, Perissodactyla) from Florida, with a review from Blancan tapirs from the state. Bulletin of the Florida Museum of Natural History 49 (3), 2010, S. 67–126