Pterostylis
Pterostylis ist eine Pflanzengattung in der Familie der Orchideen (Orchidaceae). Die etwa 270 Arten[1] kommen schwerpunktmäßig in Australien vor, aber auch auf Neuseeland, Neukaledonien, Papua-Neuguinea und weiteren Inseln im Umkreis. Es sind kleine, ausdauernde, krautige Pflanzen, die als Geophyten mit einer Knolle die ungünstige Jahreszeit überdauern. Die Blätter können bei blühenden und nicht blühenden Exemplaren derselben Art unterschiedlich ausgebildet sein. Die Blüten haben eine überwiegend grünliche Farbe, sie locken mit ihrem Duft männliche Fliegen an. Bemerkenswert ist das zentrale Blütenblatt, die Lippe, das sich bei Berührung bewegt, das bestäubende Insekt einschließt und dazu zwingt, entlang des Staubblatts und der Narbe aus der Blüte zu entkommen. Einige Arten werden als Zierpflanzen verwendet. Die Gattung Pterostylis bildet eine eigene Subtribus Pterostylidinae.
| Pterostylis | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Blüte von Pterostylis curta, Typusart der Gattung | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Untertribus | ||||||||||||
| Pterostylidinae | ||||||||||||
| Pfitzer | ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Pterostylis | ||||||||||||
| R.Br. |
Beschreibung
_pl.3086_(1831).jpg.webp)


Erscheinungsbild
Die Pterostylis-Arten sind ausdauernde, krautige Pflanzen. Diese Geophyten bilden eine unterirdische Knolle, mit der sie die ungünstige Jahreszeit überdauern. Die meisten Arten wachsen terrestrisch wurzeln also meist im Erdboden, einige Arten können auch epiphytisch wachsen.
Wurzeln, Knollen und Ausläufer
Die unterirdischen Knollen bestehen – typisch für die Unterfamilie Orchidoideae – hauptsächlich aus Wurzelgewebe, das ein Stück Sprossachse umgibt, aus dem die Pflanze wieder austreibt.[2] Die Knollen sind fleischig und meist rundlich geformt. Einige Arten aus der Verwandtschaft um Pterostylis biseta besitzen deutlich abgeflachte Knollen, einige Arten um Pterostylis spathulata elliptische bis tränenförmige. Die Knollen haben eine Lebensdauer von 10 bis 12 Monaten und werden danach durch neue ersetzt. Je nach Art wird nur eine neue Knolle oder mehrere gebildet. Die neue Knolle sitzt am Ende einer dicken, ausläuferartigen Wurzel. Bei Arten, die mehrere neue Knollen bilden, sind diese Ausläufer meist recht lang und wachsen horizontal. Dadurch ist eine vegetative Vermehrung möglich, im Laufe der Zeit können sich so Kolonien von Pflanzen bilden. Arten mit nur einer neuen Knolle bilden diese meist dicht neben der alten an einem kurzen, oft vertikal wachsenden Ausläufer.[3] In den Knollen finden sich verschiedene Kristalle: Raphiden im Cortex, häufig in Idioblasten unter der Epidermis, selten auch im Mark; stabförmige Kristalle bei einigen Arten im Cortex.[4]
Die Knollen sind von einem Velamen radicum umgeben, einer aus ein bis drei Lagen abgestorbener Zellen bestehenden Schicht. Die äußeren Zellwände der obersten Schicht sind verdickt und verkorkt, gelegentlich entspringen der äußersten Zellschicht einzellige Haare. In der unter dem Velamen liegenden Exodermis können bei einigen Arten ebenfalls verkorkte Zellwände vorkommen. Die Knolle wird meist von einer einzelnen, unzerteilten, 8- bis 40-strahligen Stele durchzogen. Auch die Ausläufer sind von Velamen umgeben, hier befinden sich mehrzellige Haare. Diese Haare und die äußeren Zellschichten der Ausläufer sind an der typischen Mykorrhiza der Orchideen beteiligt. Bei einigen Arten finden sich unter der Epidermis Raphiden.[4]
Die Wurzeln sind dünn und faserig. Auch hier finden sich mehrzellige Haare sowie Mykorrhiza-Pilze. Raphiden sind im Cortex der Wurzeln bei den meisten Arten vorhanden, bei einigen auch in der Epidermis.[4]
Sprossachse und Blätter
Aus der Knolle entspringt eine einzelne Sprossachse. Der unterirdische Teil ist mit mehrzelligen Haaren besetzt, auch hier finden sich in den äußeren Zellschichten Mykorrhiza-Pilze. Im Cortex sind meist Raphiden oder stabförmige Kristalle zu finden.[4]
Die Laubblätter befinden sich entweder gedrängt in einer Rosette oder stehen in Abständen entlang der Sprossachse. Die in Rosetten angeordneten Laubblätter können je nach Art waagrecht am Boden anliegen oder eher aufrecht stehen, manchmal ist die Blattrosette auch einige Zentimeter über dem Substrat angeordnet. Rosettenblätter können gestielt oder ungestielt sein, während bei verteilt an der Sprossachse angeordneten Blättern keine Blattstiele vorkommen. Die obersten Blätter sind kleiner als die unteren, weiter oben am Spross können auch zu Hochblättern reduzierte, der Sprossachse anliegende Blätter vorkommen. Der Blattrand ist ganzrandig oder gewellt, bei Arten aus trockenen Regionen finden sich am Blattrand häufig durchsichtige, Silikat enthaltende Zellen. Die Nervatur besteht aus einer deutlich hervortretenden Mittelrippe, die Seitenadern laufen Richtung Blattspitze parallel zum Blattrand (acrodrom) und können an ihren Enden zusammenlaufen. Netzförmige Aderung kommt nicht vor.[3] Die Laubblätter sind kahl. Stomata kommen, je nach Art, nur auf der Blattunterseite oder auf beiden Blattseiten vor. Die Kutikula ist dünn, die Epidermis der Blätter enthält häufig verzweigte Kristalle, stabförmige Kristalle oder Raphiden. Das Palisadenparenchym besteht nur aus ein oder zwei Zellschichten, kann auch ganz fehlen. Das Schwammparenchym ist drei bis acht Zelllagen dick, an der Mittelrippe auch dicker. Auch im Mesophyll finden sich häufig Kristalle.[4]
Bei einigen Arten um Pterostylis rufa und Pterostylis mutica biegen sich die Blätter der Blattrosette, entnimmt man die Pflanzen aus dem Boden, nach kurzer Zeit nach unten. Wahrscheinlich presst die Pflanze die Blätter auf den Boden, um unter den Blätter Tau und Regen zu sammeln oder um den Wasserverlust der Blattunterseite zu verringern.[3]
Die Anordnung von Knolle, Blattrosette, beblättertem Spross und Blütenstand ist je nach Art unterschiedlich. Blüht eine Pflanze in einem Jahr nicht – etwa weil sie noch jung ist oder die Umweltbedingungen ungünstig sind – bildet sie aus der Knolle eine Blattrosette. Bei blühenden Pflanzen entspringt im Grundmuster aus der Knolle der Spross, formt eine Blattrosette, über der Blattrosette setzt sich die Sprossachse als Blütenstandsstiel fort. Bei einigen Arten zeigen blühende und nicht blühende Pflanzen unterschiedliche Wachstumsmuster: Blüht eine Pflanze in einem Jahr nicht, bildet sie eine Blattrosette, während blühende Pflanzen an der Sprossachse in Abständen verteilte Blätter besitzen. Als weitere Variation gibt es Arten, bei denen blühende Pflanzen einen Spross mit sehr reduzierten Blättern sowie dem Blütenstand bilden, seitlich davon wächst an einer Verzweigung des Sprosses eine Blattrosette.[3]
_pl._3172_(1832).jpg.webp)
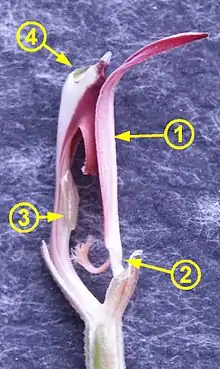
(1) – Lippe
(2) – Gelenk der Lippe
(3) – Säule – Narbe
(4) – Säule – Staubblatt
Blütenstände und Blüten
Der Blütenstand ist endständig und enthält eine oder mehrere Blüten. Die Blüten sind resupiniert, meist weisen sie vom Blütenstandsstiel weg, bei den Arten um Pterostylis parviflora sind sie jedoch nach innen, zum Blütenstandsstiel hin, orientiert.
Die zwittrigen Blüten sind zygomorph und dreizählig. Von den drei äußeren Blütenhüllblättern (Sepalen) sind die beiden seitlichen an ihrer Basis miteinander verwachsen und bilden ein sogenanntes Synsepalum. Die vordere Hälfte dieser Blütenhüllblätter ist frei und endet lang ausgezogen. Diese Spitzen sind sehr variabel gestaltet: von relativ breit-dreieckig bis zu fadenförmig, die Enden teilweise noch keulenförmig verdickt oder mit einer längs verlaufenden Rinne versehen. Die Spitzen können in dieselbe Richtung weisen wie das Blütenblatt, an dem sie sitzen, oder verschieden gebogen, bei Pterostylis recurva fast im rechten Winkel abgeknickt sein. Bei Pterostylis woollsii werden die Spitzen des Synsepalums bis zu 15 cm lang. Das Synsepalum kann nach oben gerichtet sein oder nach unten weisen – ein Merkmal, nach dem sich die Gattung systematisch in zwei Gruppen aufteilen lässt. Seine Form kann flach, aber auch schüsselförmig ausgebuchtet sein. Das dritte, nach oben weisende Sepalum und die beiden seitlichen inneren Blütenhüllblätter (Petalen) haften aneinander an und formen eine Haube. Die seitlichen Petalen sind asymmetrisch sichelförmig gebogen, ihr Rand hängt meist in einer Rinne, die längs an den Seiten des Sepalums verläuft.
Das dritte Petalum, die Lippe, ist frei; im Gegensatz zu den anderen Blütenhüllblättern setzt sie nicht am Ende des Fruchtknotens an, sondern mit einer schmalen Basis (Nagel) an der Säule. Die Lippe ist ganzrandig oder dreilappig, sie kann an der Basis ein Anhängsel besitzen. Die Oberfläche der Lippe kann glatt sein, mit reflektierenden Zellen besetzt sein, mit rundlichen oder spitzen Zellen auf der Oberfläche, oder mit verschiedenen Haaren versehen sein. Die Lippe endet meist ganzrandig, kann aber auch eingeschnitten sein oder zweilappig, bei einigen Arten ist die ganze Spitze verdickt oder besitzt eine kugelige Verdickung. Das basale Anhängsel der Lippe ist bei verschiedenen Arten sehr unterschiedlich geformt, es reicht von fadenförmig, ganzrandig bis dreilappig oder rundlich-knorpelig. Bei Berührung bewegt sich die Lippe und beschreibt einen Bogen von etwa 30 bis 150° um ein Gelenk an ihrer Basis. Bei Arten mit nach unten weisendem Synsepalum ist die Lippe sichtbar, das Anhängsel nur wenig entwickelt, die Bewegung wird durch Berührung der Lippe ausgelöst. Weist das Synsepalum nach oben, ist die Lippe dadurch verdeckt, das basale Anhängsel der Lippe ist bei diesen Arten gut entwickelt und löst bei Berührung die Bewegung der Lippe aus. Nach der Drehbewegung verbleibt die Lippe zwischen fünf und 30 Minuten in der neuen Position bevor sie sich in die Ausgangsposition zurück bewegt. Nach weiteren etwa zehn Minuten kann die Bewegung erneut ausgelöst werden.[3]
Die gebogene Säule besitzt verschiedene, je nach Art anders geformte Anhängsel: seitliche Flügel, Haare oder Bewimperung. Die Flügel sind nach vorne gebogen und formen zusammen mit der Säule eine Röhre. An der Spitze der Säule steht ein Staubblatt, das vier gelbe, linealisch bis keulenförmige Pollinien von mehlartiger Konsistenz enthält. Zu den Pollenkörnern gibt es wenige Untersuchungen: dabei wurde eine netzförmige Ausformung der Oberfläche festgestellt. Ob die Pollenkörner einzeln[2][4] oder in Tetraden[3] vorliegen, wird unterschiedlich angegeben. Die Narbe ist gelappt. Das Trenngewebe zwischen Narbe und Staubblatt ist gut ausgebildet und enthält eine Klebdrüse (Viscidium).
Früchte und Samen

Bei einigen Arten verlängert sich der Blütenstiel bis zur Reife der Kapselfrucht. Die Kapselfrucht sind länglich-elliptisch mit drei längs verlaufenden Rippen. Die vertrockneten Blütenhüllblätter bleiben an der Kapselfrucht haften. Die Kapselfrucht öffnet sich mit drei Schlitzen und entlässt die Samen.[3]
Die Samen sind elliptisch, das papierartige Integument ist flügelartig aufgeweitet und umgibt den Embryo.[3] Die Samen lassen sich dem sogenannten Goodyera-Typ zuordnen, bei dem die Zellen des Integuments alle etwa gleich groß sind und etwa quadratisch oder etwas langgestreckt geformt sind. Zwischen den Zellen befinden sich auffällige Lücken.[2][5]
Protokorm
Bei der Keimung bildet sich zunächst eine auf Mykorrhiza-Pilze angewiesene Protokorm. Es gibt innerhalb der Gattung zwei Typen: bei den meisten Arten ist die Protokorm verkehrt-eiförmig, nach oben hin dicker und wächst nach oben. Bei Arten um Pterostylis rufa, Pterostylis barbata und Pterostylis recurva wächst die Protokorm zunächst nach unten (positiv geotrop) und biegt sich erst später nach oben.[3]
Karyologie
Nur für wenige Arten gibt es Zählungen des Chromosomensatzes. Bei den meisten untersuchten Arten beträgt sie 2n = 50, aber auch 2n = 42, 2n = 52 und 2n = 54 wurden nachgewiesen.[4]
Lebenszyklus
Die Samen von Pterostylis reifen etwa drei bis sechs Wochen nach der Bestäubung der Blüten. Die Keimung findet bei guten Bedingungen sofort statt, die Samen können aber auch die trockenen Sommermonate überdauern. Um zu keimen, muss der Samen auf einen Mykorrhiza-Pilz treffen, der den Embryo durch die Mikropyle erreicht, Pilzfäden wachsen in den Embryo und werden dort von der Orchidee verdaut. Die Protokorm wächst so in Abhängigkeit vom Pilz heran. Unter guten Bedingungen blühen die Pflanzen zwei oder drei Jahre nach der Keimung zum ersten Mal.[3]
Pterostylis kommt in Gegenden mit saisonalem Klima vor, die Pflanzen überdauern die heiße, trockene Jahreszeit mit ihren Knollen und wachsen während der Regenzeit. Meist dauert die Trockenzeit im Areal der Gattung nicht länger als zwei oder drei Monate. Für Arten aus dem südlichsten Bereich sowie Arten aus den Hochlagen der Gebirge ist die Ruhezeit durch Kälte vorgegeben. Während der Ruhezeit finden sich in der Knolle kaum Mykorrhiza-Pilze, beim Einsetzen des Wachstums bildet sich die Mykorrhiza an den neuen Wurzeln und am unterirdischen Teil der Sprossachse neu.[3]
Viele Arten wachsen in Ökosystemen, die durch häufige Brände gekennzeichnet sind. Im Gegensatz zu anderen australischen Orchideen blühen die Pflanzen nach Bränden nicht besser, sondern die Blüte fällt in solchen Jahren schwächer aus.[3]
Ökologie
Bestäubung
Die meisten Pterostylis-Arten werden von Insekten bestäubt (Entomophilie), bei einigen Arten kommt auch Selbstbestäubung vor. Die Blüten bieten keinen Nektar oder andere Vorteile für die Bestäuber, es handelt sich um Täuschblumen, zumindest bei einem Teil der Arten um Sexualtäuschblumen. Als Bestäuber wurden kleine Fliegen aus den Familien Mycetophilidae und Culicidae beobachtet. Obwohl bei kaum einer Pterostylis-Art ein für den Menschen wahrnehmbarer Blütenduft verströmt wird (Ausnahme sind Arten um Pterostylis parviflora, die deutlich nach Sperma riechen, besonders bei feuchtem oder schwülem Wetter), fliegen die Bestäuber die Blüten gegen den Wind an und folgen wahrscheinlich einem Duft. Die Aktivität der Fliegen an den Blüten ist bei gewittrigem Wetter oder bei deutlich fallendem Luftdruck am größten.[3]
Der Bestäubungsmechanismus unterscheidet sich, je nachdem ob das Synsepalum nach oben weist und die Lippe verdeckt oder ob es nach unten weist und die Lippe sichtbar ist. Im ersten Fall landet das Insekt außen auf der Blüte und kriecht nach innen, erreicht dort die Lippe und bewegt sich auf der Lippe weiter Richtung Blüteninneres. An einem bestimmten Punkt löst das Insekt die Bewegung der Lippe aus – die Lippe bewegt sich um ihr Gelenk und drückt das Insekt gegen die Säule. Der Weg aus der Blüte hinaus ist jetzt für das Insekt weitgehend versperrt. Verschiedene Teile der Blüte, wie die Lippe, die Säule, die Flügel der Säule, auch deren Behaarung, greifen ineinander und lassen für die kleinen Fliegen keine Lücke. Wohl durch die Bewegung der Lippe und den versperrten Weg alarmiert, versuchen die Insekten die Blüte schnell zu verlassen. Der einzige Weg führt entlang der Säule, vorbei an der Narbe, der Klebdrüse und dann am Staubblatt. Dabei werden zuerst eventuell am Insekt hängende Pollenkörner auf der Narbe platziert, dann das Insekt mit klebender Flüssigkeit bestrichen und schließlich am Staubblatt Pollen angeheftet. Durch diese Reihenfolge wird vermieden, dass eigener Pollen auf der Narbe der Blüte landet. Bei den Pterostylis-Arten mit sichtbarer Lippe geschieht die Bestäubung nur durch männliche Fliegen. Beim Anflug auf die Lippe fahren diese ihre Genitalien aus und landen direkt auf diesem Blütenblatt. Bei einigen Pflanzen ist die Lippe ähnlich wie das weibliche Insekt geformt. Die Bewegung der Lippe wird direkt durch die Landung der Fliege ausgelöst, das Insekt wird gegen die Säule gedrückt und muss die Blüte entlang des oben beschriebenen Wegs verlassen.[3]
Die Arten, bei denen Selbstbestäubung vorkommt, sind die Pollinien bröselig und fallen von selbst auf die darunter liegende Narbe.[3]
Samenausbreitung
Die Samen werden bei allen Arten durch den Wind ausgebreitet (Anemochorie). Obwohl die Samen sehr leicht sind, landet die Mehrzahl doch in relativ geringer Entfernung. Bei einigen Arten verlängert sich der Blütenstiel zwischen Bestäubung und Samenreife (Arten um Pterostylis parviflora, Pterostylis mutica). Bei den beiden Arten Pterostylis uliginosa und Pterostylis humilis verdickt und verlängert sich in dieser Zeit der ganze Blütenstandsstiel, was den Samen eine weitere Verbreitung erlaubt.[3]
Mykorrhiza
Alle Arten sind bei der Keimung auf Mykorrhiza-Pilze angewiesen. Auch bei ausgewachsenen Pflanzen findet man diese Pilze, vor allem am unterirdischen Teil des Sprosses. Die Wurzeln sind bei den Pterostylis-Arten sehr gering ausgebildet, was darauf deutet, dass die Pflanzen in ihrer Ernährung zu einem großen Teil auf die Mykorrhiza-Pilze angewiesen sind.[3] Die beteiligten Pilze gehören zur Gattung Ceratobasidium innerhalb der Tulasnellales. Eine genaue Bestimmung der Pilzart ist schwierig, aufgrund von DNA-Untersuchungen lassen sich verschiedene Verwandtschaftslinien unterscheiden, von denen viele nah mit Ceratobasidium cornigerum verwandt sind. Einige Gruppen innerhalb der Gattung Pterostylis lassen sich einer bestimmten Pilzart zuordnen, das heißt nah miteinander verwandte Pflanzenarten beherbergen denselben Pilz. Das trifft auf die Sektionen Catochilus und Stamnorchis zu, deren Arten nahezu identische Mykorrhiza-Pilze aufweisen, auch wenn sie geografisch weit voneinander entfernt wachsen. Arten aus den Sektionen Hymenochilus und Oligochaetochilus zeigen dieselbe Spezialisierung auf einen gemeinsamen, von anderen Pterostylis-Sektionen unterscheidbaren Pilz. Die Sektionen Catochilus und Stamnorchis sind miteinander nah verwandt, ebenso die Sektionen Hymenochilus und Oligochaetochilus, hier reflektiert also die Abhängigkeit von einer speziellen Pilzart die Verwandtschaftsverhältnisse. In nah miteinander verwandten Orchideenarten finden sich nah miteinander verwandte Pilze, und umgekehrt finden sich eine Pilzart nur in Orchideen, die nah miteinander verwandt sind. Das gilt nicht für die gesamte Gattung Pterostylis: es lassen sich auch Gegenbeispiele finden wo die Verwandtschaftsverhältnisse der Mykorrhiza-Partner keine Gemeinsamkeit aufweisen. So besitzen die Arten der Sektion Pterostylis etliche unterschiedliche Pilzpartner; andersherum finden sich in nicht besonders nah verwandten Arten aus den Sektionen Pharochilum, Urochilos und Parviflorae dieselben Pilze.[6]
Bei der Keimung der Samen lässt sich im Labor ebenfalls eine Zuordnung von bestimmten Pilzarten und Pterostylis-Arten feststellen. So keimen die Samen generell besser, wenn der Mykorrhiza-Pilz aus derselben oder einer nah verwandten Pterostylis-Art stammt. Die Abhängigkeit von einer bestimmten Pilzart bei der Keimung und als erwachsene Pflanzen korreliert miteinander: die Arten der Sektion Pterostylis, bei denen aus erwachsenen Pflanzen unterschiedliche Pilze isoliert wurden, können auch mit unterschiedlichen Pilzen recht gut keimen. In Sektionen, die als erwachsene Pflanze auf ganz bestimmte Pilze angewiesen sind, zeigen auch die Samen eine wesentlich bessere Keimrate mit ebendieser Pilzart.[6]
Vorkommen
Die Pterostylis-Arten kommen hauptsächlich in Australien vor, des Weiteren in Neuseeland, Neuguinea, Neukaledonien, Neubritannien, New Ireland und auf Seram. Der südlichste Punkt des Areals ist Stewart Island südlich von Neuseeland, der nördlichste Punkt liegt auf Neuguinea fast in Höhe des Äquators. Es werden Höhenlagen von Meereshöhe bis 3600 Metern auf Neuguinea besiedelt. Die Mehrzahl der Arten und die größte Diversität findet man im südlichen Australien. Auf Neuseeland sind etwa 30 Arten beheimatet, fünf auf Neukaledonien und nur vier in Melanesien.
Vor allem im temperaten Klima besiedeln die Pterostylis-Arten sehr verschiedene Standorte. Grasland, Gebüsche, offene Wälder bis hin zu Regenwäldern werden besiedelt. Die Böden können aus sauren Sanden, Kalkstein oder Granit bestehen, auch in Mooren und Sümpfen sowie an Flussufern finden sich einige Arten. Das trockene Innere Australiens wird allerdings von Pterostylis nicht erreicht. Die ökologische Amplitude der Arten ist dabei ganz unterschiedlich: einige Arten sind weit verbreitet, Pterostylis aspera, Pterostylis curta und Pterostylis nutans etwa findet man auch in angepflanzten Kiefern-Plantagen. Viele Arten haben sich aber auf Sonderstandorte spezialisiert, so wächst etwa Pterostylis tenuissima nur in kalkhaltigen Sümpfen nahe der Küste.[3][4]
Systematik und botanische Geschichte
Erste Spuren in der wissenschaftlichen Literatur hinterließ die Gattung Pterostylis in einem illustrierten Werk zu Joseph Banks' Reise. Dort sieht man eine Zusammenstellung aus drei Blüten, wahrscheinlich von Pterostylis revoluta und Blättern von Pterostylis acuminata. In einem unveröffentlichten Manuskript Daniel Solanders vergab dieser den wissenschaftlichen Namen Arethusa tetrapetala für die heute als Pterostylis revoluta bekannte Art. Frühe Herbarexemplare finden sich im J.-E.-Smith-Herbarium der Linnean Society of London. Sie wurden zwischen 1788 und 1794 von John White sowie 1792 von Jacques Labillardière gesammelt. 1806 beschrieb Labillardière eine Art unter dem Namen Disperis alata.[3]
_-_page_326_detail.jpg.webp)
George Caley und Robert Brown waren die ersten Sammler, die eine größere Anzahl an Arten und Exemplaren in ihren Herbarien zusammentrugen. Beide waren im Südosten Australiens zwischen 1800 und 1810 tätig, vielleicht sammelten sie gemeinsam, in Browns Herbarium finden sich jedenfalls einige Pflanzen, die Caley sammelte. Robert Brown beschrieb 1810 die Gattung mit 19 Arten in seinem Werk Prodromus floræ Novæ Hollandiæ. Typusart ist Pterostylis curta.[3] Der Gattungsname Pterostylis setzt sich aus den altgriechischen Bestandteilen πτερόν pteron, deutsch ‚Flügel‘ oder ‚Feder‘, und στῦλος stylos, deutsch ‚Säule‘ oder ‚Griffel‘ zusammen; er bezieht sich auf die seitlich des Staubblatts an der Säule befindlichen, deutlichen Flügel.[7]
Externe Systematik
Frühe Einordnungen der Gattung Pterostylis innerhalb der Familie der Orchideen gab es verschiedene: Lindley stellte sie 1840 in die Tribus Arethuseae, Endlicher folgte ihm 1842 in dieser Sichtweise. George Bentham sah 1873 die Gattung in der Tribus Neottieae, später platzierte er sie in dem 1883 gemeinsam mit Hooker herausgegebenen Werk Species plantarum in der Tribus Diurideae. Pfitzer stellte 1887 eine Subtribus Pterostylidinae auf, in der er die Gattungen Pterostylis, Caleana und Drakaea versammelte. Schlechter bezweifelte die nahe Verwandtschaft dieser drei Gattungen und errichtete 1926 die Subtribus Drakaeinae, so dass Pterostylis als einzige Gattung der Subtribus Pterostylidinae innerhalb der Diurideae verblieb.[3] Diese Einteilung blieb im Wesentlichen die nächsten 70 Jahre stabil. Auch Dressler folgte ihr, bemerkte aber, dass Pterostylis von den anderen Diurideae deutlich zu unterscheiden sei und die nächsten Verwandten innerhalb der Tribus unklar seien.[2]
Ab 1995 führten zuerst Studien der Embryonalentwicklung, der Samen und dann DNA-Studien zu einer gänzlich anderen Sichtweise. Demnach ist Pterostylis nicht näher mit der Tribus Diurideae verwandt, sondern gehört zur Tribus Cranichideae. Ganz ähnlich war der Befund zu der Subtribus Chloraeinae sowie zu Achlydosa – zuerst zu den Diurideae gezählt, stellten die neueren Untersuchungen sie zu den Cranichideae. Die genauen Verwandtschaftsverhältnisse der basalen Gruppen der Tribus Cranichideae sind noch recht unklar,[4] ein Kladogramm aus einer Studie von Salazar, Chase, Soto Arenas und Ingrouille sieht folgendermaßen aus:[8]
| ||||||||||||||||||||||||||||||||||||||||||||||
|
Diurideae | ||||||||||||||||||||||||||||||||||||||||||||||
Interne Systematik


Schon bei der Erstbeschreibung der Gattung ordnete Brown die ihm bekannten Arten in vier Gruppen. Die ersten drei der Brown'schen informellen Gruppen wurden 1830 von Don formal als Sektionen beschrieben. Weitere, teilweise recht unterschiedliche interne Gliederungen der Gattung nahmen Lindley 1840, Reichenbach 1871, Bentham 1873, Pfitzer 1889 und Rupp 1933 vor. Die von anderen Autoren zahlreich neu beschriebenen Arten wurden nur selten in eine dieser Gliederungen eingefügt. In gärtnerischen Kreisen haben sich für einzelne leicht abgrenzbare Gruppen eigene Namen eingebürgert.[3]
In den Jahren 2001 und 2002 wurden in kurzer Folge zwei Bearbeitungen zur Systematik der Gattung Pterostylis veröffentlicht. In seiner Übersichtsarbeit Genera et Species Orchidalium teilte Szlachetko die Gattung in drei Teile auf: Pterostylis und die zwei neu beschriebenen Oligochaetochilus und Plumatichilos.[9] Die Monographie von Jones und Clements musste, da früher aufgestellte Namen Vorrang haben, auf die Arbeit von Szlachetko Rücksicht nehmen. Die beiden australischen Forscher übernahmen aber nicht Szlachetkos Konzept, sondern präsentierten eine Unterteilung von Pterostylis in 16 einzelne Gattungen.[3]
In der Folge wurde das splitting insbesondere australischer Orchideen-Taxa als „taxonomic turmoil down-under“[10] kritisiert. Obwohl Jones und Clements kladistische Methoden und DNA-Untersuchungen angewandt hatten, stellten weitere Arbeiten die Monophylie einiger der neuen Gattungen in Frage.[11] Eine Reihe von australischen Institutionen, etwa die Beteiligten am Australian Plant Census und an dem Australian Plant Names Index sowie die meisten Herbarien beschlossen, die Gattung Pterostylis in ihrer „klassischen“ Form zu behalten und den Arbeiten von Jones und Clements nicht zu folgen. Als Alternative stellten Janes und Duretto eine Gliederung vor, die Pterostylis nicht in mehrere Gattungen unterteilt, sondern mit internen Rangstufen wie Untergattung und Sektion versieht.[12]
Wenn man die Gattung Pterostylis nach Janes und Duretto 2010 in einem breiteren Sinne auffasst dann sind Synonyme für Pterostylis R.Br. nom. cons.: Diplodium Sw., Oligochaetochilus Szlach., Plumatichilos Szlach., Bunochilus D.L.Jones & M.A.Clem., Crangonorchis D.L.Jones & M.A.Clem., Eremorchis D.L.Jones & M.A.Clem., Hymenochilus D.L.Jones & M.A.Clem., Linguella D.L.Jones & M.A.Clem., Petrorchis D.L.Jones & M.A.Clem., Pharochilum D.L.Jones & M.A.Clem., Ranorchis D.L.Jones & M.A.Clem., Speculantha D.L.Jones & M.A.Clem., Stamnorchis D.L.Jones & M.A.Clem., Taurantha D.L.Jones & M.A.Clem., Urochilus D.L.Jones & M.A.Clem., ×Taurodium D.L.Jones & M.A.Clem.[12][1]
_pl._3401_(1835).jpg.webp)


_pl_6351_(1878).jpg.webp)
_pl._3400_(1835).jpg.webp)
_pl.3085_(1831).jpg.webp)
.jpg.webp)



.jpg.webp)
.jpg.webp)
.jpg.webp)

.jpg.webp)

Die Gattung Pterostylis wird demnach in zwei Untergattungen geteilt, die jeweils etwa die Hälfte der Arten enthalten. Die Untergattung Pterostylis wird weiter in drei Sektionen unterteilt, die Untergattung Oligochaetochilus in sieben Sektionen:
- Untergattung Pterostylis:
- Charakterisiert durch nach oben gerichtete seitliche Sepalen (schräg nach oben bei Pterostylis porrecta), die Lippe ist ungelappt, ohne Bewimperung, das basale Anhängsel pinselförmig. Der Blütenstand ist einblütig oder, wenn mehrblütig, mit kurzen Blütenstielen und Blüten, die zum Blütenstandsstiel hin weisen. Einige Arten formen Kolonien durch vegetative Vermehrung.
- Sektion Pterostylis:
- Blühende und nicht blühende Pflanzen gleichen sich in den vegetativen Merkmalen, die Blätter stehen in einer grundständigen Rosette oder sind locker spiralig angeordnet. Die Blätter am Blütenstandsstiel sind zu kleinen Hochblättern reduziert. Bei allen Arten kommt vegetative Vermehrung vor. Die Blüten erscheinen einzeln, die seitliche Sepalen sind nach oben oder schräg nach oben gerichtet, sie enden lang und dünn ausgezogen, zwischen den seitlichen Sepalen und dem oberen Sepal ist eine deutliche Lücke zu sehen.
- Die Sektion Pterostylis enthält etwa 50 Arten mit einem weiten Verbreitungsgebiet, sie fehlt allerdings im westlichen Australien. Diese Sektion ist in der Untergattung Pterostylis die basale Klade, das heißt, sie ist das Schwestertaxon der beiden anderen Sektionen. Diese Sektion enthält mit Pterostylis curta die Typusart der Gattung, deshalb trägt sie denselben Namen wie die Gattung.
- Sektion Foliosae G.Don:
- Blühende und nicht blühende Pflanzen sind je nach Art vegetativ gleich oder unterschiedlich ausgebildet. Bei nicht blühenden Pflanzen stehen die Blätter in einer grundständigen Rosette oder locker spiralig, bei blühenden Pflanzen können die Blätter in einer Rosette oder verteilt entlang der Sprossachse stehen. Alle Arten vermehren sich auch vegetativ. Die Blüten erscheinen einzeln, die seitlichen Sepalen sind nach oben gerichtet, sie enden fadenförmig ausgezogen, zwischen den seitlichen Sepalen und dem oberen Sepal ist keine Lücke.
- Die Sektion Foliosae enthält ebenfalls etwa 50 Arten, das Verbreitungsgebiet umfasst Australien und Neuseeland. Diese Sektion ist das Schwestertaxon der Sektion Parviflorae. Typusart der Sektion ist Pterostylis grandiflora. Clements und Jones unterteilen diese Sektion in die Gattungen Crangonorchis, Diplodium, Eremorchis, Linguella und Taurantha.
- Sektion Parviflorae (Benth.) Janes & Duretto:
- Blühende und nicht blühende Pflanzen unterscheiden sich in ihren vegetativen Merkmalen: bei nicht blühenden Pflanzen stehen die Blätter in einer grundständigen Rosette, bei blühenden Pflanzen werden ein oder mehrere seitliche Blattrosetten gebildet. Am Blütenstandsstiel befinden sich Laubblätter oder reduzierte Hochblätter. Vegetative Vermehrung kommt nicht vor. Der Blütenstand ist mehrblütig, die Blütenstiele sind kurz, die Blüten weisen zum Blütenstandsstiel hin. Die seitlichen Sepalen sind nach oben gerichtet, sie enden relativ kurz und stumpf dreieckig.
- Die Sektion Parviflorae enthält elf Arten mit einem Verbreitungsgebiet im südöstlichen Australien. Typusart der Sektion ist Pterostylis parviflora. Clements und Jones unterteilen diese Sektion in die Gattungen Petrorchis und Speculantha.
- Sektion Pterostylis:
- Untergattung Oligochaetochilus:
- In dieser Untergattung weisen die seitlichen Sepalen nach unten, selten schräg nach unten oder zuerst aufwärts mit dann nach unten gebogener Spitze. Die Lippe kann gelappt sein, Bewimperung kann vorhanden sein, das basale Anhängsel kann pinselförmig, bandförmig oder dreilappig sein oder ganz fehlen. Der Blütenstand ist ein- oder mehrblütig, die Blüten weisen nach außen. Vegetative Vermehrung kommt nicht vor.
- Sektion Hymenochilus (D.L.Jones & M.A.Clem.) Janes & Duretto:
- Blühende und nicht blühende Pflanzen gleichen sich in den vegetativen Merkmalen, die Blätter stehen in einer grundständigen Rosette. Die Blätter am Blütenstandsstiel sind zu kleinen Hochblättern reduziert. Der Blütenstand ist mehrblütig, die Blüten zeigen nach außen. Die seitlichen Sepalen weisen nach unten, sie enden kurz dreieckig. Die Lippe ist ungeteilt, sehr kurz bewimpert, das basale Anhängsel ist band- oder schnabelförmig.
- Die Sektion Hymenochilus enthält 16 Arten, das Verbreitungsgebiet erstreckt sich über das südliche und östliche Australien sowie Neuseeland. Sie ist Schwestertaxon zu einer Klade aus den anderen sechs Sektionen der Untergattung Oligochaetochilus. Typusart der Sektion ist Pterostylis muticus.
- Sektion Catochilus Benth.:
- In dieser Sektion gleichen sich blühende und nicht blühende Pflanzen, die Blätter stehen in einer grundständigen Rosette sowie spiralig an der Sprossachse, nach oben kleiner werdend bis zu reduzierten Hochblättern. Der Blütenstand ist einblütig. Die seitlichen Sepalen weisen nach unten, sie enden lang linealisch ausgezogen. Die Lippe ist ungeteilt, mit langen, gelben Haaren, das basale Anhängsel ist schnabelförmig.
- Vier Arten bilden diese Sektion, deren Schwestertaxon innerhalb der Untergattung nicht geklärt ist. Das Verbreitungsgebiet umfasst das südliche Australien und Neuseeland. Typusart ist Pterostylis barbata. Szlachetko sowie Clements und Jones sehen diese Sektion im Gattungsrang unter dem Namen Plumatichilos.
- Sektion Oligochaetochilus (Szlach.) Janes & Duretto:
- Blühende und nicht blühende Pflanzen gleichen sich in den vegetativen Merkmalen, die Blätter stehen in einer grundständigen Rosette. Die Blätter am Blütenstandsstiel sind zu kleinen Hochblättern reduziert. Der Blütenstand ist mehrblütig, die Blüten zeigen nach außen. Die seitlichen Sepalen weisen nach unten, sie enden lang fadenförmig ausgezogen. Die Lippe ist ungeteilt, mit auffälligen weißen Haaren besetzt, ein basales Anhängsel fehlt.
- Mit 47 Arten die größte Sektion der Untergattung, wie bei der vorigen Sektion ist das Schwestertaxon nicht klar. Verbreitungsgebiet ist Australien. Typusart der Sektion ist Pterostylis rufa.
- Sektion Stamnorchis (D.L.Jones & M.A.Clem.) Janes & Duretto:
- Blühende und nicht blühende Pflanzen sind vegetativ unterschiedlich ausgebildet. Bei nicht blühenden Pflanzen stehen die Blätter in einer grundständigen Rosette, bei blühenden Pflanzen nur verteilt entlang der Sprossachse. Der Blütenstand ist mehrblütig, die Blüten weisen nach außen. Die beiden seitlichen Sepalen weisen, soweit sie zusammengewachsen sind, schräg nach oben, die freien Spitzen sind nach unten gebogen, sie enden lang fadenförmig ausgezogen. Die Lippe ist dreilappig, ohne auffällige Behaarung, mit einem pinselförmigen basalen Anhängsel.
- Die Sektion ist monotypisch mit der einzigen Art Pterostylis recurva aus dem Südwesten Australiens. Das Schwestertaxon ist unklar.
- Sektion Pharochilum (D.L.Jones & M.A.Clem.) Janes & Duretto:
- Blühende und nicht blühende Pflanzen sind vegetativ unterschiedlich, bei nicht blühenden Pflanzen stehen die Blätter in einer grundständigen Rosette, bei blühenden Pflanzen sind seitlich ein oder zwei Blattrosetten ausgebildet, während an der blühenden Sprossachse nur reduzierte Hochblätter stehen. Der Blütenstand ist mehrblütig, die Blüten weisen nach außen. Die beiden seitlichen Sepalen weisen schräg nach unten, sie enden lang fadenförmig ausgezogen. Die Lippe ist dreilappig mit großen Seitenlappen, ohne Behaarung und ohne basales Anhängsel.
- Monotypische Sektion mit Pterostylis daintreana als einziger Art, sie besitzt ein Areal im Osten Australiens. Das Schwestertaxon ist unklar.
- Sektion Urochilus (D.L.Jones & M.A.Clem.) Janes & Duretto:
- Blühende und nicht blühende Pflanzen unterscheiden sich vegetativ: nicht blühende Exemplare bilden eine basale Blattrosette, bei blühenden Pflanzen stehen die Blätter am Spross verteilt. Der Blütenstand ist mehrblütig, die Blüten weisen nach außen. Die seitlichen Sepalen zeigen nach unten, sie enden lang fadenförmig ausgezogen oder kurz dreieckig. Die Lippe besitzt höchstens sehr kleine Haare, sie ist dreilappig mit einem dreigeteilten basalen Anhängsel.
- Diese Sektion umfasst vier Arten aus dem südlichen Australien. Schwestertaxon ist die Sektion Squamatae. Die Typusart ist Pterostylis vittata. Clements und Jones unterteilen diese Sektion in die Gattungen Ranorchis und Urochilus.
- Sektion Squamatae G.Don:
- Die Arten der Sektion Squamatae ähneln äußerlich denen der Sektion Urochilus. Unterscheidungsmerkmal ist das basale Anhängsel der Lippe, das bei Squamatae fehlt. Die 27 Arten stammen alle aus dem südöstlichen Australien. Typusart ist Pterostylis longifolia. Clements und Jones sehen diese Sektion im Gattungsrang unter dem Namen Bunorchis.[12]
- Sektion Hymenochilus (D.L.Jones & M.A.Clem.) Janes & Duretto:
Liste der Arten und Hybriden
Eine Liste der anerkannten Arten der Gattung Pterostylis mit ihrer Verbreitung findet sich bei R. Govaerts:[1]
- Pterostylis abrupta D.L.Jones
- Pterostylis aciculiformis (Nicholls) M.A.Clem. & D.L.Jones
- Pterostylis acuminata R.Br.
- Pterostylis aenigma D.L.Jones & M.A.Clem.
- Pterostylis aestiva D.L.Jones
- Pterostylis agathicola D.L.Jones, Molloy & M.A.Clem.
- Pterostylis agrestis (D.L.Jones) G.N.Backh.: Sie wurde 2010 aus dem australischen Bundesstaat Victoria erstbeschrieben.[1]
- Pterostylis alata (Labill.) Rchb. f.
- Pterostylis allantoidea R.S.Rogers
- Pterostylis alobula (Hatch) L.B.Moore
- Pterostylis alpina R.S.Rogers
- Pterostylis alveata Garnet
- Pterostylis amabilis (D.L.Jones & L.M.Copel.) D.L.Jones: Diese Neukombination erfolgte 2015. Sie kommt im australischen Bundesstaat New South Wales vor.[1]
- Pterostylis anaclasta (D.L.Jones) Janes & Duretto
- Pterostylis anatona D.L.Jones
- Pterostylis aneba D.L.Jones
- Pterostylis angulata (D.L.Jones & C.J.French) D.L.Jones & C.J.French
- Pterostylis angusta A.S.George
- Pterostylis antennifera (D.L.Jones) D.L.Jones
- Pterostylis aphylla Lindl.
- Pterostylis aquilonia D.L.Jones & B.Gray
- Pterostylis arbuscula (D.L.Jones & C.J.French) D.L.Jones & C.J.French
- Pterostylis arenicola M.A.Clem. & J.Stewart
- Pterostylis areolata Petrie
- Pterostylis arfakensis (J.J.Sm.) D.L.Jones & M.A.Clem.
- Pterostylis aspera D.L.Jones & M.A.Clem.
- Pterostylis atrans D.L.Jones
- Pterostylis atriola D.L.Jones
- Pterostylis atrosanguinea (D.L.Jones & C.J.French) D.L.Jones & C.J.French
- Pterostylis auriculata Colenso
- Pterostylis australis Hook. f.
- Pterostylis banksii R.Br. ex A.Cunn.
- Pterostylis baptistii Fitzg.
- Pterostylis barbata Lindl.,
- Pterostylis barringtonensis (D.L.Jones) G.N.Backh.
- Pterostylis basaltica D.L.Jones & M.A.Clem.
- Pterostylis bicolor M.A.Clem. & D.L.Jones
- Pterostylis bicornis D.L.Jones & M.A.Clem.
- Pterostylis biseta Blackmore & Clemesha
- Pterostylis boormanii Rupp
- Pterostylis borealis (D.L.Jones) D.L.Jones
- Pterostylis brumalis L.B.Moore
- Pterostylis bruniella (R.J.Bates) J.M.H.Shaw: Sie wurde 2017 aus South Australia erstbeschrieben.[1]
- Pterostylis brunneola D.L.Jones & C.J.French: Sie wurde 2014 aus dem australischen Bundesstaat Western Australia erstbeschrieben.[1]
- Pterostylis bryophila D.L.Jones
- Pterostylis bureaviana Schltr.
- Pterostylis calceolus M.A.Clem.
- Pterostylis campestris (R.J.Bates) J.M.H.Shaw
- Pterostylis cardiostigma D.Cooper
- Pterostylis caulescens L.O.Williams
- Pterostylis cernua D.L.Jones, Molloy & M.A.Clem.
- Pterostylis chaetophora M.A.Clem. & D.L.Jones
- Pterostylis cheraphila D.L.Jones & M.A.Clem.
- Pterostylis chlorogramma D.L.Jones & M.A.Clem.
- Pterostylis chocolatina (D.L.Jones) G.N.Backh.
- Pterostylis ciliata M.A.Clem. & D.L.Jones
- Pterostylis clavigera Fitzg.
- Pterostylis clivicola (D.L.Jones) G.N.Backh.
- Pterostylis clivosa (D.L.Jones) D.L.Jones
- Pterostylis cobarensis M.A.Clem.
- Pterostylis coccina Fitzg.
- Pterostylis collina (Rupp) M.A.Clem. & D.L.Jones
- Pterostylis commutata D.L.Jones
- Pterostylis concava D.L.Jones & M.A.Clem.
- Pterostylis concinna R.Br.
- Pterostylis conferta (D.L.Jones) G.N.Backh.
- Pterostylis corpulenta (D.L.Jones) D.L.Jones
- Pterostylis crassa (D.L.Jones) G.N.Backh.
- Pterostylis crassicaulis (D.L.Jones & M.A.Clem.) G.N.Backh.
- Pterostylis crassichila D.L.Jones
- Pterostylis crebra (D.L.Jones) D.L.Jones
- Pterostylis crebriflora (D.L.Jones & C.J.French) D.L.Jones & C.J.French
- Pterostylis cucullata R.Br.
- Pterostylis curta R.Br.
- Pterostylis cycnocephala Fitzg.
- Pterostylis daintreana F.Muell. ex Benth.
- Pterostylis decurva R.S.Rogers
- Pterostylis depauperata F.M.Bailey
- Pterostylis despectans (Nicholls) M.A.Clem. & D.L.Jones
- Pterostylis dilatata A.S.George
- Pterostylis diminuta (D.L.Jones) G.N.Backh.
- Pterostylis divaricata (D.L.Jones & L.M.Copel.) J.M.H.Shaw
- Pterostylis diversiflora (R.J.Bates) J.M.H.Shaw
- Pterostylis dolichochila D.L.Jones & M.A.Clem.
- Pterostylis dubia R.Br.
- Pterostylis ectypha (D.L.Jones & C.J.French) D.L.Jones & C.J.French
- Pterostylis elegans D.L.Jones
- Pterostylis elegantissima (D.L.Jones & C.J.French) D.L.Jones: Diese Neukombination erfolgte 2015. Sie kommt im australischen Bundesstaat Western Australia vor.[1]
- Pterostylis erecta T.E.Hunt
- Pterostylis eremaea (D.L.Jones & C.J.French) D.L.Jones & C.J.French
- Pterostylis erythroconcha M.A.Clem. & D.L.Jones
- Pterostylis exalla (D.L.Jones) G.N.Backh.
- Pterostylis excelsa M.A.Clem.
- Pterostylis exquisita (D.L.Jones) D.L.Jones
- Pterostylis extranea (D.L.Jones) Janes & Duretto
- Pterostylis faceta (D.L.Jones, C.J.French & M.A.Clem.) D.L.Jones & C.J.French
- Pterostylis ferruginea (D.L.Jones) G.N.Backh.
- Pterostylis fischii Nicholls
- Pterostylis flavovirens (D.L.Jones) R.J.Bates
- Pterostylis foliata Hook.f.
- Pterostylis frenchii (D.L.Jones) A.P.Br.
- Pterostylis fuliginosa (D.L.Jones & C.J.French) D.L.Jones & C.J.French
- Pterostylis furcata Lindl.
- Pterostylis furva (D.L.Jones) D.L.Jones
- Pterostylis galgula (D.L.Jones & C.J.French) D.L.Jones & C.J.French
- Pterostylis gibbosa R.Br.
- Pterostylis glyphida (D.L.Jones) G.N.Backh.
- Pterostylis graminea Hook.f.
- Pterostylis grandiflora R.Br.
- Pterostylis hamata Blackmore & Clemesha
- Pterostylis hamiltonii Nicholls
- Pterostylis heberlei (D.L.Jones & C.J.French) D.L.Jones & C.J.French
- Pterostylis hians D.L.Jones
- Pterostylis hildae Nicholls
- Pterostylis hispidula Fitzg.
- Pterostylis humilis R.S.Rogers
- Pterostylis incognita (D.L.Jones) G.N.Backh.
- Pterostylis insectifera M.A.Clem.
- Pterostylis irsoniana Hatch
- Pterostylis irwinii D.L.Jones
- Pterostylis jonesii G.N.Backh.
- Pterostylis laxa Blackmore
- Pterostylis lepida (D.L.Jones) G.N.Backh.
- Pterostylis leptochila M.A.Clem. & D.L.Jones
- Pterostylis lineata (D.L.Jones) G.N.Backh.
- Pterostylis lingua M.A.Clem.
- Pterostylis littoralis (D.L.Jones) R.J.Bates
- Pterostylis loganii (D.L.Jones) G.N.Backh.
- Pterostylis longicornis (D.L.Jones & C.J.French) D.L.Jones & C.J.French
- Pterostylis longicurva Rupp
- Pterostylis longifolia R.Br.
- Pterostylis longipetala Rupp
- Pterostylis lustra D.L.Jones
- Pterostylis macilenta (D.L.Jones) G.N.Backh.
- Pterostylis macrocalymma M.A.Clem. & D.L.Jones
- Pterostylis macrosceles (D.L.Jones & C.J.French) D.L.Jones
- Pterostylis macrosepala (D.L.Jones) G.N.Backh.
- Pterostylis major (D.L.Jones) G.N.Backh.
- Pterostylis maxima M.A.Clem. & D.L.Jones
- Pterostylis melagramma D.L.Jones
- Pterostylis meridionalis (D.L.Jones & C.J.French) D.L.Jones & C.J.French
- Pterostylis metcalfei D.L.Jones
- Pterostylis micromega Hook. f.
- Pterostylis mirabilis (D.L.Jones) R.J.Bates
- Pterostylis mitchellii Lindl. in T.L.Mitchell
- Pterostylis montana Hatch
- Pterostylis monticola D.L.Jones
- Pterostylis multiflora (D.L.Jones) G.N.Backh.
- Pterostylis mutica R.Br.
- Pterostylis mystacina (D.L.Jones) Janes & Duretto
- Pterostylis nana R.Br.
- Pterostylis nichollsiana (D.L.Jones) D.L.Jones
- Pterostylis nigricans D.L.Jones & M.A.Clem.
- Pterostylis nutans R.Br.
- Pterostylis oblonga D.L.Jones
- Pterostylis obtusa R.Br.
- Pterostylis oliveri Petrie
- Pterostylis ophioglossa R.Br.
- Pterostylis orbiculata (D.L.Jones & C.J.French) D.L.Jones & C.J.French
- Pterostylis ovata M.A.Clem.
- Pterostylis paludosa D.L.Jones, Molloy & M.A.Clem.
- Pterostylis papuana Rolfe
- Pterostylis parca (D.L.Jones) G.N.Backh.
- Pterostylis parva D.L.Jones & C.J.French: Die 2015 erstbeschriebene Art kommt in Western Australia vor.[1]
- Pterostylis parviflora R.Br.
- Pterostylis patens Colenso
- Pterostylis pearsonii (D.L.Jones) Janes & Duretto
- Pterostylis pedina (D.L.Jones) Janes & Duretto
- Pterostylis pedoglossa Fitzg.
- Pterostylis pedunculata R.Br.
- Pterostylis perculta (D.L.Jones & C.J.French) D.L.Jones
- Pterostylis petrosa D.L.Jones & M.A.Clem.
- Pterostylis picta M.A.Clem.
- Pterostylis planulata D.L.Jones & M.A.Clem.
- Pterostylis platypetala D.L.Jones & C.J.French: Die 2015 erstbeschriebene Art kommt in Western Australia vor.[1]
- Pterostylis plumosa Cady
- Pterostylis porrecta D.L.Jones, Molloy & M.A.Clem.
- Pterostylis praetermissa M.A.Clem. & D.L.Jones
- Pterostylis prasina (D.L.Jones) G.N.Backh.
- Pterostylis pratensis D.L.Jones
- Pterostylis precatoria (D.L.Jones, C.J.French & M.A.Clem.) D.L.Jones & C.J.French
- Pterostylis procera D.L.Jones & M.A.Clem.
- Pterostylis psammophilus (D.L.Jones) R.J.Bates
- Pterostylis puberula Hook.f.
- Pterostylis pulchella Messmer
- Pterostylis pusilla R.S.Rogers
- Pterostylis pyramidalis Lindl.
- Pterostylis recurva Benth.
- Pterostylis reflexa R.Br.
- Pterostylis repanda (M.A.Clem. & D.L.Jones) J.M.H.Shaw
- Pterostylis revoluta R.Br.
- Pterostylis riparia D.L.Jones
- Pterostylis robusta R.S.Rogers
- Pterostylis roensis M.A.Clem. & D.L.Jones
- Pterostylis rogersii E.Coleman
- Pterostylis rubenachii D.L.Jones
- Pterostylis rubescens (D.L.Jones) G.N.Backh.
- Pterostylis rufa R.Br.
- Pterostylis russellii T.E.Hunt
- Pterostylis sanguinea D.L.Jones & M.A.Clem.
- Pterostylis sargentii C.R.P.Andrews
- Pterostylis saxicola D.L.Jones & M.A.Clem.
- Pterostylis saxosa (D.L.Jones & C.J.French) D.L.Jones & C.J.French
- Pterostylis scabra Lindl.
- Pterostylis scabrida Lindl.
- Pterostylis scapula (D.L.Jones) D.L.Jones
- Pterostylis scitula D.L.Jones & C.J.French: Die 2015 erstbeschriebene Art kommt in Western Australia vor.[1]
- Pterostylis scoliosa D.L.Jones
- Pterostylis serotina (D.L.Jones, C.J.French & M.A.Clem.) D.L.Jones & C.J.French
- Pterostylis setifera M.A.Clem.
- Pterostylis sigmoidea (D.L.Jones & C.J.French) D.L.Jones & C.J.French
- Pterostylis silvicultrix (F.Muell.) D.L.Jones & M.A.Clem.
- Pterostylis sinuata (D.L.Jones) Janes & Duretto
- Pterostylis smaragdyna D.L.Jones & M.A.Clem.
- Pterostylis spatalium J.M.H.Shaw
- Pterostylis spathulata M.A.Clem.
- Pterostylis spissa (D.L.Jones) G.N.Backh.
- Pterostylis splendens D.L.Jones & M.A.Clem.
- Pterostylis squamata R.Br.
- Pterostylis stenochila D.L.Jones
- Pterostylis stenosepala (D.L.Jones) G.N.Backh.
- Pterostylis stricta Clemesha & B.Gray
- Pterostylis subtilis D.L.Jones
- Pterostylis tanypoda D.L.Jones, Molloy & M.A.Clem.
- Pterostylis tasmanica D.L.Jones
- Pterostylis taurus M.A.Clem. & D.L.Jones
- Pterostylis tenuicauda Kraenzl.
- Pterostylis tenuis (D.L.Jones) G.N.Backh.
- Pterostylis tenuissima Nicholls
- Pterostylis thulia (D.L.Jones) Janes & Duretto
- Pterostylis timorensis Schuit. & J.J.Verm.
- Pterostylis timothyi (D.L.Jones) Janes & Duretto
- Pterostylis torquata D.L.Jones
- Pterostylis tristis Colenso
- Pterostylis trullifolia Hook. f.
- Pterostylis truncata Fitzg.
- Pterostylis tryphera (D.L.Jones & C.J.French) D.L.Jones & C.J.French
- Pterostylis tunstallii D.L.Jones & M.A.Clem.
- Pterostylis turfosa Endl. in J.G.C.Lehmann
- Pterostylis uliginosa D.L.Jones
- Pterostylis umbrina (D.L.Jones) G.N.Backh.
- Pterostylis venosa Colenso
- Pterostylis ventricosa (D.L.Jones) G.N.Backh.
- Pterostylis vernalis (D.L.Jones) G.N.Backh.
- Pterostylis vescula (D.L.Jones) D.L.Jones
- Pterostylis virens (D.L.Jones & C.J.French) D.L.Jones & C.J.French
- Pterostylis viriosa (D.L.Jones) R.J.Bates
- Pterostylis vitrea (D.L.Jones) Bostock
- Pterostylis vittata Lindl.
- Pterostylis voigtii D.L.Jones & C.J.French: Die 2015 erstbeschriebene Art kommt in Western Australia vor.[1]
- Pterostylis wapstrarum D.L.Jones
- Pterostylis williamsonii D.L.Jones
- Pterostylis woollsii Fitzg.
- Pterostylis xerampelina (D.L.Jones & C.J.French) D.L.Jones & C.J.French
- Pterostylis xerophila M.A.Clem.
- Pterostylis zebrina (D.L.Jones & C.J.French) D.L.Jones
- Pterostylis ziegeleri D.L.Jones
Nothospecies:
- Pterostylis × aenigma D.L.Jones & M.A.Clem. = Pterostylis cucullata × Pterostylis foliata
- Pterostylis ×conoglossa Upton
- Pterostylis ×furcillata Rupp = Pterostylis alveata × Pterostylis ophioglossa
- Pterostylis ×ingens (Rupp) D.L.Jones = Pterostylis falcata × Pterostylis nutans
- Pterostylis× toveyana Ewart & Sharman = Pterostylis alata × Pterostylis concinna
Siehe auch
Literatur
- Alec M. Pridgeon, Phillip Cribb, Mark W. Chase, Finn Rasmussen (Hrsg.): Genera Orchidacearum. Orchidoideae (Part 2). Vanilloideae. Band 3/2. Oxford University Press, New York/Oxford 2003, ISBN 0-19-850711-9, S. 156–164 (englisch).
- David L. Jones, Mark A. Clements: A Review of Pterostylis (Orchidaceae) (= Australian Orchid Research. Band 4). Australian Orchid Foundation, 2002, ISBN 0-642-54904-4 (englisch).
Einzelnachweise
- Rafaël Govaerts (Hrsg.): Pterostylis. In: World Checklist of Selected Plant Families (WCSP) – The Board of Trustees of the Royal Botanic Gardens, Kew, abgerufen am 28. März 2020.
- Robert L. Dressler: Phylogeny and Classification of the Orchid Family. Cambridge University Press, Cambridge 1993, ISBN 0-521-45058-6, S. 133 (englisch).
- David L. Jones, Mark A. Clements: A Review of Pterostylis (Orchidaceae). Australian Orchid Foundation, 2002, ISBN 0-642-54904-4.
- Alec M. Pridgeon, Phillip Cribb, Mark W. Chase, Finn Rasmussen (Hrsg.): Genera Orchidacearum. Orchidoideae (Part 2). Vanilloideae. 3/2, Oxford University Press, New York und Oxford 2003, ISBN 0-19-850711-9, S. 156–164.
- Robert L. Dressler: Phylogeny and Classification of the Orchid Family. Cambridge University Press, Cambridge 1993, ISBN 0-521-45058-6, S. 52 (englisch).
- J. Tupac Otero, Peter H. Thrall, Mark Clements, Jeremy J. Burdon, Joseph T. Miller: Codiversification of orchids (Pterostylidinae) and their associated mycorrhizal fungi. In: Australian Journal of Botany. Band 59, 2011, S. 480–497, doi:10.1071/BT11053 (englisch).
- Hubert Mayr: Orchideennamen und ihre Bedeutung. Berger, 1996.
- Gerardo A. Salazar, Mark W. Chase, Miguel A. Soto Arenas, Martin Ingrouille: Phylogenetics of Cranichideae with emphasis on Spiranthinae (Orchidaceae, Orchidoideae): evidence from plastid and nuclear DNA sequences. In: American Journal of Botany. Band 90, Nr. 5, 2003, S. 782 (englisch).
- Dariusz L. Szlachetko: Genera et Species Orchidalium. I. In: Polish Journal of Botany. Band 46, Nr. 1, 2001, S. 11–26.
- Stephen D. Hopper: Taxonomic turmoil down-under. Recent developments in Australian orchid systematics. In: Annals of Botany. Band 104, 2009, S. 447–455, doi:10.1093/aob/mcp090 (englisch).
- Jasmine K. Janes, Dorothy A. Steane, René E. Vaillancourt, Marco F. Duretto: A molecular phylogeny of the subtribe Pterostylidinae (Orchidaceae). Resolving the taxonomic confusion. In: Annals of Botany. Band 23, 2010, S. 248–259, doi:10.1071/SB10006 (englisch).
- Jasmine K. Janes, Marco F. Duretto: A new classification for subtribe Pterostylidinae (Orchidaceae), reaffirming Pterostylis in the broad sense. In: Australian Systematic Botany. Band 23, 2010, S. 260–269, doi:10.1071/SB09052 (englisch).