Kobralilie
Die Kobralilie (Darlingtonia californica) ist eine präkarnivore Pflanze und die einzige Art der monotypischen Gattung Darlingtonia aus der Familie der Schlauchpflanzengewächse (Sarraceniaceae). Sie ist eng mit den karnivoren Schlauchpflanzen (Sarracenia) verwandt.
| Kobralilie | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Kobralilie (Darlingtonia californica) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Darlingtonia | ||||||||||||
| Torr. | ||||||||||||
| Wissenschaftlicher Name der Art | ||||||||||||
| Darlingtonia californica | ||||||||||||
| Torr. |
Merkmale
Die Kobralilie ist eine immergrüne, mehrjährige, krautige Pflanze mit einem Rhizom von ein bis zwei Zentimetern Durchmesser. Sie wächst sehr langsam, erst nach zwei bis drei Jahren nimmt sie ihre typische Gestalt an und nach sieben bis zehn Jahren blüht sie erstmals.
Wurzel
Die Pflanzen haben keine Hauptwurzel, sondern bilden aus dem Rhizom heraus zahlreiche feine Haarwurzeln.
Blätter/Falle
Das direkt aus dem Rhizom heraus treibende Blattwerk der Pflanzen besteht ausschließlich aus den typischen Schlauchblättern, die eine lose Rosette bilden. Die Blätter sind schwach kegelförmig und verbreitern sich von der Basis an. Sie sind hohl und werden 60 bis 80 Zentimeter, gelegentlich bis zu einem Meter hoch, über ihre gesamte Länge zieht sich auf der Vorderseite des Blattes eine Flügelleiste. An ihrem oberen Ende sind sie von einer helmartigen Haube überdacht, an deren Vorderende ein umgedreht V-förmiges Anhängsel herabhängt. Hinter dieser „Schlangenzunge“ liegt eine kleine, zum Boden zeigende Öffnung, die von einem Peristom umkränzt ist. Die Blätter stehen entweder aufrecht oder wachsen fast waagerecht zum Boden, sodass die Schlangenzunge den Boden berührt und, anders als die aufrechten Blätter, hauptsächlich bodenlebende Insekten anzieht. Jedes Blatt ist bis zu 270 Grad in sich verdreht, sodass Öffnung und Schlangenzunge von der Rosette weg zeigen – vermutlich um Beute besser anzulocken.
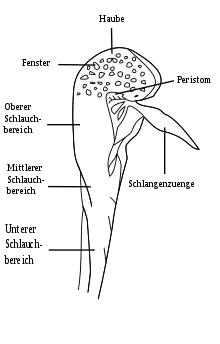
Die Blätter werden in sechs Zonen unterteilt: Zone 1 ist die „Schlangenzunge“, Zone 2 das Peristom, Zone 3 die Haube, Zonen 4 bis 6 jeweils tiefer liegende Abschnitte des eigentlichen Schlauches. Jede dieser Zonen hat beim Beutefang eine Funktion, für die sie entsprechend ausgestattet ist.
- Zone 1: Die Schlangenzunge. Sie ist meist auffällig rot eingefärbt und auf ihrer Rückseite mit zahlreichen Nektarien versehen, beides dient zur Anlockung der Beute. Eine gerichtete Behaarung der Oberfläche macht es dem Insekt dabei leichter, zur Schlauchöffnung hin voranzugehen, als sich von dieser abzuwenden.
- Zone 2: Die Öffnung. Sie wird von einem Peristom gebildet, das besonders große Mengen Nektar ausscheidet und so die Beute vom Anhängsel in den eigentlichen Schlauch lockt.
- Zone 3: Die Haube. Sie überdacht den Schlauch und die hervorkragende Öffnung, wodurch sie ein Entweichen der Beute nach oben verhindert. An ihrem höchsten Punkt ist sie gehäuft mit chlorophyllfreien Flecken versehen, die nahezu ungehindert das Außenlicht passieren lassen und so wie Fenster wirken. Gefangene Beutetiere versuchen nun, durch diese „Fenster“ die Falle zu verlassen, bei diesen „Fluchtversuchen“ stürzen sie nun in den aufrechten Schlauch.
- Zone 4: Oberer Schlauchbereich. Dieser sich direkt unterhalb der Haube befindende Abschnitt des Schlauches ist mit einer Wachsschicht und nach unten gerichteten Haaren versehen. Beides zwingt die Beute weiter abwärts.
- Zone 5: Mittlerer Schlauchbereich. Hier befinden sich die abwärts gerichteten Haare in besonderer Dichte, Wachsflecken finden sich kaum noch.
- Zone 6: Unterer Schlauchbereich. Diese Zone ist rein glatt und mit keinerlei Fangeinrichtung mehr ausgestattet, sie dient allein der Verdauung. In ihr befindet sich die Flüssigkeit der Pflanze, die für die Verdauung sorgt, diese wird von der Pflanze regelrecht in die Schläuche hineingepumpt, verstärkt nach einem Beutefang. Die Oberfläche des Schlauches ist durch eine fehlende Kutikula fähig zur Aufnahme der Nährstoffe, die durch eine symbiotische Bakterienfauna, aber auch durch Kommensalen gelöst werden.
Trotz dieser aufwändigen Fallenkonstruktion ist die Kobralilie im Vergleich zu ihren Verwandten, den Schlauchpflanzen, kein besonders guter Beutefänger. Auch ist sie selbst nicht zur Produktion von Verdauungsenzymen imstande, sondern angewiesen auf die Zersetzung des Fangs durch Bakterien in der Lösung. Das Beutespektrum umfasst aufgrund der zwei verschiedenen Blattformen (aufrecht stehend und am Boden liegend) sowohl Flug- als auch Bodeninsekten.

Blüten
Der mit Hochblättern besetzte, ab Mai sich ausbildende Blütenstand wird bis zu einem Meter hoch, damit überragt er, für eine Karnivore ungewöhnlich, die Fallen nur geringfügig; zum Blühzeitpunkt sind die Fallen allerdings noch nicht aktiv. Die einzelne, hängende, selbstfertile, jedoch schwach protandrische Blüte ist ungewöhnlich gebaut: Die gelblich-hellgrünen Kelchblätter sind etwas länger als die purpurnen Kronblätter. 1994 wurde allerdings ein Vorkommen von rund 30 Individuen entdeckt, bei dem die Kronblätter nicht purpurn sind, sondern von gleicher Farbe wie die Kelchblätter. Diese öffnen sich nicht, sondern bilden eine Art Kapsel, die für mögliche Bestäuber nur durch kleine, durch Einwölbungen der Kronblätter am Rand nahe der Spitze gebildete Öffnungen zugänglich ist. Für welche Art von Bestäubern diese komplexe Blüte gebildet ist, ist jedoch trotz teils langjähriger Beobachtungen der Art immer noch nicht eindeutig bekannt, vermutlich sind Spinnen daran beteiligt, der – für Menschen unangenehme – Duft der Blüte lässt aber auch die Vermutung zu, dass Fliegen beteiligt sind.
Frucht und Samen
Nach erfolgter Bestäubung hebt sich der Fruchtknoten allmählich, so dass die Kapselfrucht aufrecht steht. Die ausgebildete Frucht ist umgekehrt-eiförmig und enthält etwa zehn Wochen nach der Bestäubung mehrere hundert bis über tausend sandfarbene, behaarte Samen von zwei bis drei Millimetern Länge, der Embryo ist recht groß, Endosperm ist vorhanden. Durch ihre Behaarung sind diese schwimmfähig und treiben mit der Strömung von der Mutterpflanze fort (Bythisochorie).
Vegetative Vermehrung
Das Rhizom der Pflanzen bildet Ausläufer, die mit zunehmendem Alter einer Kolonie große Geflechte bilden können. Jede Nodie des Rhizoms wiederum kann eigenständig neue Wurzeln und Blätter ausbilden. An vielen Standorten ist die Vermehrungsrate durch Ausläufer erheblich höher als die durch Samen. An besonders nährstoffarmen Standorten dienen die Rhizomgeflechte möglicherweise auch zum Nährstoffaustausch zwischen den einzelnen Individuen einer Kolonie.

Verbreitung
Die Art ist endemisch im Nordwesten der USA. Sie findet sich nur in Westoregon und Nordkalifornien, ein einzelnes Vorkommen am Chase Lake bei Seattle im Staate Washington gilt als angesalbt. Hauptsächlich findet sie sich in Gebirgslagen (Kaskadenkette, Sierra Nevada, Klamath Mountains, Siskiyou Mountains, Trinity Mountains) auf bis zu 2500 m NN, steigt aber in Oregon bei entsprechenden Bedingungen auch bis auf Meeresniveau unmittelbar an die Pazifikküste herab (z. B. in Florence, Oregon, wo sich mit dem nur sieben Hektar großen Darlingtonia Botanical Wayside das einzige Naturschutzgebiet Oregons befindet, das nur einer einzigen Art gewidmet ist). Die Zahl der angegebenen Vorkommen schwankt dabei zwischen 200 und 250, die sich einigermaßen gleichmäßig auf Oregon und Kalifornien aufteilen.
Habitate

Kobralilien wachsen bevorzugt an Standorten mit felsigem Untergrund (meist Serpentin), geringem Nährstoffangebot und hohem Schwermetallanteil (dem gegenüber die Pflanzen tolerant sind). Die Vorkommen sind häufig dicht und ausgedehnt, das größte bekannte Vorkommen in den Siskiyou Mountains umfasst mehrere Tausend Pflanzen. Häufige Standorte sind Feuchtwiesen und Moore, aber auch Flussufer oder Böden aus reinem (kalkfreiem) Sand in Kiefernwäldern. Alle Standorte sind südlich oder südwestlich ausgerichtet und liegen in Zonen hoher Niederschläge (1000 bis 2000 mm).
Für das Gedeihen der Pflanze ist ein hoher Spiegel kühlen Grundwassers und reichlich fließendes Wasser bei zugleich gut drainiertem Boden von Bedeutung, welche für die notwendige kühle Wurzeltemperatur sorgen. Ebenso wichtig sind offene und sonnige Standorte, die im Verbreitungsgebiet eigentlich durch periodisch auftretende Brände gewährleistet bleiben. Diese Pflanzenart ist bedingt winterhart und liegt an einigen Standorten zu dieser Zeit unter Schnee und Eis. Die Art findet sich in Gesellschaft unter anderem mit Torfmoosen, dem Rundblättrigen Sonnentau, dem Gemeinen Fettkraut, Pinguicula macroceras ssp. nortensis sowie Panther-Lilien (Lilium pardalinum).
Gefährdung und Status
Seit 1981 ist die Kobralilie im Anhang 2 des Washingtoner Artenschutzabkommens (CITES) gelistet, wodurch der Handel mit Wildpflanzen genehmigungspflichtig und streng reglementiert ist. Darüber hinaus unterliegt die Art auch unterschiedlichen Unterschutzstellungen auf staatlicher, regionaler und lokaler Ebene; all diesen ist gemeinsam, dass sie die Art als nicht unmittelbar gefährdet, aber wegen ihres kleinen Verbreitungsgebietes und den speziellen Standortanforderungen als relativ verletzlich einstufen.
Als hauptsächlich gefährdende Faktoren gelten Bergbauaktivitäten (das Verbreitungsgebiet ist vielfach reich an Nickel, Chrom und Cobalt), Holzfällaktivitäten, Straßenbau sowie Freizeit- und Siedlungsdruck (letzteres vor allem in Oregon), wodurch ebenso Habitate zerstört werden wie mittelbar durch die Unterdrückung von Bränden, die als natürliche Mahd wirken. Die bis in die 80er Jahre stärkste Bedrohung war das Absammeln der Pflanzen durch Händler oder Sammler, diese hat sich seither durch die diversen Unterschutzstellungen und das gestiegene Umweltbewusstsein von Sammlern erheblich verringert. Immerhin einige Vorkommen befinden sich in Schutzgebieten und sind so als ungefährdet anzusehen.
Lebensgemeinschaften
Zahlreiche Insektenarten leben in und an Kobralilien, einige von ihnen in sehr enger Lebensgemeinschaft. Häufige Kommensalen sind Metriocnemus edwardsii (eine Fliege), Eperigone trilobata (eine Spinnenart) sowie Sarraceniopus darlingtoniae, eine Milbenart, die ausschließlich in den Schläuchen von Kobralilien vorkommt.

Systematik
Innerhalb der Gattung gibt es nur die eine Art, ohne weitere Unterarten oder Varietäten, allerdings ist eine Albino-Form bekannt. Zur Abgrenzung von den eng verwandten Schlauchpflanzen dient neben morphologischen Merkmalen wie dem Blütenaufbau, der Fähigkeit zur Regulierung des Standes der Verdauungsflüssigkeit in den Schläuchen und der Präkarnivorie vor allem das von den Schlauchpflanzen im Osten und Süden der USA isolierte und beträchtlich entfernte Verbreitungsgebiet. Letzteres wird auch als bestätigendes Merkmal der Klassifikation als eigene Gattung herangezogen.
Die Phylogenie der Familie ist nur wenig erforscht, molekulargenetische Untersuchungen ergaben, dass die Kobralilie ein Schwestertaxon der beiden anderen Gattungen der Familie darstellt[1]:
| |||||||||||||||||||
Botanische Geschichte
Entdeckt wurde die Pflanze erst 1841 in Feuchtwiesen südlich des Mount Shasta durch William Dunlop Brackenridge, Mitglied einer botanischen Expedition der US-Regierung. Die Erstbeschreibung erfolgte 1850 durch John Torrey, wurde aber erst 1853 veröffentlicht, der botanische Gattungsname (den sie durch einen schönen Zufall mit einer Natterngattung im Tierreich teilt) verweist auf den amerikanischen Arzt und Botaniker William Darlington (1782–1863), das Artepitheton auf den ersten Fund in Kalifornien.
1871 wurde die Kobralilie in den Kew Gardens in Kultur genommen, heutzutage findet sie sich in vielen Botanischen Gärten und privaten Karnivorensammlungen, vereinzelt auch in Alpin- und Felsgärten.
In den 70er Jahren des 19. Jahrhunderts erforschte Rebecca M. Austin als erste die Kobralilie. Über mehrere Jahre hinweg betrieb sie intensive Feldforschung und teilte von 1875 an bis 1877 ihre Ergebnisse brieflich dem Botaniker William Marriott Canby mit, der sie darin unterstützte und ermutigte. Ihre Aufzeichnungen wurden allerdings bis heute nur auszugsweise veröffentlicht.
1891 stellte sich heraus, dass der Name Darlingtonia bereits 1825 von de Candolle für ein Mimosengewächs vergeben worden war. Dadurch war der Name ungültig, woraufhin die Gattung von Greene in Chrysamphora umbenannt wurde. Nachdem aber die nun als Darlingtonia geführte Gattung 1954 in die Gattung Desmanthus eingegliedert wurde, war quasi der Weg frei und eine ICBN-Kommission erlaubte die Rückbenennung der Chrysamphora californica in Darlingtonia californica, weil dieser der etabliertere war (nomen conservandum).
Nachweise
- Wilhelm Barthlott, Stefan Porembski, Rüdiger Seine, Inge Theisen: Karnivoren. Biologie und Kultur fleischfressender Pflanzen. Eugen Ulmer GmbH & Co., Stuttgart 2004, ISBN 3-8001-4144-2.
- Guido J. Braem: Fleischfressende Pflanzen. Arten und Kultur. 2., durchgesehene Auflage. Weltbild, Augsburg 2002, ISBN 3-426-66762-2.
- Christine Leigh Elder: Reproductive Biology of Darlingtonia californica. 1997, (Arcata CA, Humboldt State University, Master's Thesis, 1997; Digitalisat (PDF; 2,33 MB)).
- Aaron M. Ellison, Elizabeth J. Farnsworth: The Cost of Carnivory for Darlingtonia californica (Sarraceniaceae): Evidence from Relationships among Leaf Traits. In: American Journal of Botany. Bd. 92, Nr. 7, 2005, ISSN 0002-9122, S. 1085–1093, JSTOR 4126151.
- Norman James Fashing: Biology of Sarraceniopus darlingtoniae. In: Phytophaga. Bd. 14, 2004, ISSN 0393-8131, S. 299–305, (online).
- Website zur Kobralilie
- Ausführliche Website zur Kobralilie in Englisch
- Informationen zur Pflanze vom U.S. Fish and Wildlife Service
- Darlingtonia californica in der Roten Liste gefährdeter Arten der IUCN 2006. Eingestellt von: Schnell, D., Catling, P., Folkerts, G., Frost, C., Gardner, R., et al., 2000. Abgerufen am 12. Mai 2006.
Einzelnachweise
Die Informationen dieses Artikels entstammen zum größten Teil den unter Nachweise angegebenen Quellen, darüber hinaus werden folgende Quellen zitiert: