Elektrische Orientierung
Elektrische Orientierung (auch: Elektrorezeption oder Elektrosensorik) bezeichnet die Fähigkeit einiger Lebewesen, ein elektrisches Feld über spezielle Rezeptoren wahrzunehmen. Dies wird auch als passive elektrische Orientierung bezeichnet.
Einige Lebewesen verschiedener Reiche können mithilfe elektrischer Organe schwache oder starke elektrische Ladungen erzeugen und an ihre Umgebung abgeben (Electric Organ Discharge, EOD). Zur Orientierung werden dann elektrische Felder erzeugt und empfangen (Sender-Empfänger-Kombination). Dies wird als aktive elektrische Orientierung bezeichnet. Zusätzlich kann die aktiv erzeugte Elektrizität zur Jagd, zur Verteidigung oder zur innerartlichen Kommunikation eingesetzt werden.
Sowohl Salz- als auch Süßwasser leiten elektrischen Strom aufgrund der gelösten beweglichen Ladungsträger (Ionen). Wegen der besonders guten elektrischen Leitfähigkeit von Salzwasser treten Fische mit aktiver elektrischer Orientierung überwiegend im Süßwasser auf, da sie sich ansonsten selbst verletzen könnten.[1]
Gruppen der Lebewesen
Einige Arten verschiedener Gruppen von Lebewesen sind zur elektrischen Orientierung befähigt, es kann jedoch sein, dass nahe verwandte Arten nicht dazu imstande sind. Die folgenden Tierarten sind daraufhin untersucht, sie stehen stellvertretend für nicht genannte Arten, erlauben aber keine Aussagen zur Befähigung nahestehender Arten.
Mikroorganismen
Für etliche Mikroorganismen wie Schizosaccharomyces pombe ist nachgewiesen, dass sie sich am elektrischen Feld ausrichten.[2]
Blütenpflanzen
Viele Blütenpflanzen sind zu aktiver elektrischer Orientierung befähigt. Viele Pollen tragen statische Ladungen,[3] daher kann die Pflanze die Ankunft von Pollen (durch Insekten) in der Blüte registrieren und die Blütenöffnung verbessern.[4][5] Das von ihr erzeugte elektrische Feld kann die Pflanze innerhalb von Sekunden ändern, um auf einfallenden Pollen oder Insekten zu reagieren.[6] Dieses Phänomen wird zur industriellen elektrostatischen Bestäubung genutzt.[3][7]
Insekten
Für Blüten besuchende Insekten wie die Honigbiene[8] oder die Dunkle Erdhummel[6] und viele andere Insekten[9] ist die Fähigkeit zur Elektrorezeption belegt. Das durch die Pflanze erzeugte elektrische Feld leitet das Insekt, wie der Duft oder die optisch wirkenden Farben, zur Blüte. Hummeln, Taufliegen, Schmeißfliegen und andere Insekten beeinflussen das elektrische Feld aktiv, da sie statische elektrische Ladung tragen.[9]
Rundmäuler
Neunaugen besitzen eine passive elektrische Orientierung und vermutlich besaßen die gemeinsamen Vorfahren der Wirbeltiere, Rundmäuler und Kiefermäuler, ein entsprechendes Sinnesorgan.[10]
Knorpelfische
Der passive elektrische Orientierungssinn ist innerhalb der Klasse der Knorpelfische bei vielen Haien und Rochen vorhanden.[11][12]
Innerhalb der Familie der Echten Rochen (Rajidae) besteht auch aktive elektrische Orientierung.[13]
Ein sehr gut untersuchtes Beispiel innerhalb der Familie der Zitterrochen ist der Marmor-Zitterrochen Torpedo marmorata.
Knochenfische
Viele Knochenfische haben die Fähigkeit zur passiven elektrischen Orientierung und können mit ihren Elektrorezeptoren Beute, Feinde und das Erdmagnetfeld feststellen. Ein Elektrosinn wurde bisher u. a. beim Südamerikanischen Lungenfisch, bei verschiedenen Störartigen (z. B. Löffelstör & den Schaufelstören), bei den Flösselhechten, vielen Welsartigen (wahrscheinlich sind alle elektrorezeptiv) und den afrikanischen Arten der Altwelt-Messerfische (Afrikanischer Fähnchen-Messerfisch und Afrikanischer Messerfisch), bei den asiatischen aber nicht. Bei Knochenfischen ist die aktive elektrische Orientierung eher von untergeordneter Bedeutung. Die Elektrorezeptoren entwickelten sich aus dem Seitenlinienorgan, das dabei seine ursprüngliche Funktion einbüßte. Elektrische Organe haben sich aber bei Bodenformen und Bewohnern trüber und lichtarmer Gewässer drei- bis viermal unabhängig voneinander entwickelt. Insgesamt besitzen neun Familien aus vier Ordnungen elektrische Organe, unter den Welsen die Zitterwelse, alle fünf Familien der Neuwelt-Messerfische (Gymnotiformes), die Nilhechte und der Großnilhecht aus der Ordnung der Knochenzünglerartigen und die Elektrischen Sterngucker (Astroscopus) (Familie Himmelsgucker (Uranoscopidae), die einzigen Barschverwandten mit elektrischen Organen),[14] zusammen ca. 400 Arten.
Säugetiere
Den passiven elektrischen Orientierungssinn findet man auch beim Schnabeltier.
Der auch in Flüssen vorkommende Guyana-Delfin besitzt einen passiven elektrischen Orientierungssinn.[15]
Organe
Elektrorezeptoren
Um elektrische Ströme oder die Veränderung selbst generierter elektrischer Felder zu erkennen, gibt es spezielle Poren in der Haut der Fische, an deren Basis die Elektrorezeptoren liegen. Diese Rezeptoren unterscheiden sich voneinander in der Hinsicht, dass sie unterschiedliche Aspekte von elektrischen Feldern wahrnehmen. Darüber hinaus sind einige auf eigene Entladungen abgestimmt, andere auf die von Artgenossen oder verwandten Arten.
Die Elektrorezeptoren haben keine eigenen Axone und sind somit sekundäre Sinneszellen. Sie sind von Haarsinneszellen abgeleitet und werden von Ästen des Nervus statoacusticus innerviert.
Man unterscheidet morphologisch zwei Arten von Elektrorezeptoren: ampulläre Rezeptoren und tuberöse Rezeptoren. Sie unterscheiden sich hauptsächlich dadurch, dass die ampullären Rezeptoren, wie z. B. die Lorenzinischen Ampullen bei Haien nach außen hin geöffnet sind. Hingegen sind die tuberösen Rezeptoren nicht nach außen geöffnet und deswegen auch weniger empfindlich.
Die apikale Membran der Rezeptorzellen hat einen geringen elektrischen Widerstand. So kann ein elektrischer Impuls durch die Membran die Rezeptorzellen passieren und die basale Rezeptorzellmembran depolarisieren. Dies löst den Einstrom von Calcium-Ionen aus, wodurch wiederum Transmitter freigesetzt werden. Die Transmittermoleküle beeinflussen postsynaptisch die Entladungsfrequenz der afferenten Nervenfasern, deren Terminalen an der basalen Rezeptormembran angeheftet sind. Die jeweilige Erhöhung oder auch Verminderung der Entladungsfrequenz hängt davon ab, wie stark das elektrische Feld, das der Fisch ausgesandt hat, beeinflusst wird, oder welche Art von elektrischen Impulsen eintrifft.
Lorenzinische Ampullen
Lorenzinische Ampullen sind die Elektrorezeptoren der Knorpelfische und dienen zur passiven Orientierung. Sie können geomagnetische Felder, von Tieren durch Muskelaktivität erzeugte elektrische Felder und große Meeresströmungen wahrnehmen.
Lorenzinische Ampullen sind schlauchförmige Ampullenorgane, die sich von den Seitenlinienorganen ableiten. Das eine Ende der Ampulle ist röhrenförmig zum Außenmedium offen, und das andere Ende bildet eine geschlossene, kugelförmige Kammer. Die Röhre ist mit einer elektrisch sehr leitfähigen gelatinösen Substanz gefüllt (Mucus), welche auch zum Schutz der Elektrorezeptoren vor dem Außenwasser dient, wohingegen die Röhrenwand einen hohen elektrischen Widerstand besitzt. Das macht die Röhre zu einem sehr guten elektrischen Leiter. Die Röhre verbindet die apikale Membrane der Elektrorezeptoren, von denen bis zu Hundert in der Kammerwand liegen, durch die gelatinöse Substanz mit dem Außenmedium.
Die Lorenzinische Ampulle ist ein hochempfindlicher Gleichspannungsmesser. Durch die Rezeptoren fließt ein ständiger Ruhestrom, der tonische Impulse auslöst. Die einzelnen Rezeptoren sind ionendicht mit ihren Nachbarzellen verknüpft, so dass nur durch deren apikale Membran Strom fließen kann. Ändert sich das elektrische Feld des Fisches, ändert sich die Ruheentladungsfrequenz nach oben oder unten (je nach Richtung des fließenden Stromes).
Dadurch, dass die einzelnen Eingänge der Ampullenorgane konvergent verschaltet werden, kann eine höhere Empfindlichkeit auf dem Rezeptorniveau erreicht werden (Siehe zentralnervöse Verarbeitung). Haie können mit den Lorenzinischen Ampullen elektrische Felder wahrnehmen, die nur 0,01 bis 0,05 Mikrovolt pro Zentimeter betragen.
- Bedeutung der Lorenzinischen Ampullen am Beispiel der Haie
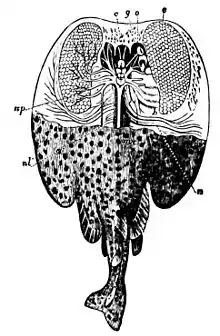
Bei Haien liegen die Lorenzinischen Ampullen an Kopf und Schnauze und sind als kleine, dunkle Porenöffnungen sichtbar. Die Ampullen dienen aufgrund ihrer Eigenschaften zur Elektroortung. Lebende Organismen verursachen unvermeidlich eine elektrische Spannung durch Muskelaktivität die sogar wahrnehmbar ist, wenn sie unter Sand vergraben sind. Allerdings kann eine solche Ortung nur in einem sehr geringen Radius stattfinden. So dienen die Lorenzinischen Ampullen wohl der Ortung der Beute im letzten Moment des Angriffes oder der Suche am Boden nach vergrabenen Lebewesen.
Zudem dienen die Ampullen auch als „geomagnetischer Kompass“, da Meeresströmungen unweigerlich elektrische Magnetfelder hervorrufen, die der Hai spüren kann. Somit können sich die Haie zum Beispiel zu einer bestimmten Zeit an einem ganz bestimmten Ort zur Paarung versammeln.
Einigen Erkenntnissen zufolge dienen die Lorenzinischen Ampullen auch zur Thermorezeption.[16] Abkühlung erhöht die Entladungsrate, wohingegen eine Erwärmung die Frequenz der Aktionspotentiale kurzzeitig senkt.
Tubulusorgane
Zur Wahrnehmung der Veränderung der eigens ausgesandten elektrischen Felder besitzen die Neuwelt-Messerfische (Gymnotiformes) Tubulusorgane, die auf ihre hochfrequenten elektrischen Entladungen abgestimmt sind. Diese sind ähnlich den Lorenzinischen Ampullen in der Haut versenkt, jedoch durch Deckzellen kapazitiv mit dem Außenmedium (Wasser) gekoppelt. Somit können hochfrequente Wechselfelder wahrgenommen werden. Da das Organ am Rand mit „tight junctions“ abgeschirmt ist, wirken die Deckzellen wie eine Linse, die den Spannungsgradienten direkt auf die Rezeptorzellen leitet. Diese Rezeptorzellen liegen auf dem Tubulusgrund auf und sind mit etwa 95 % ihrer Membranoberfläche dem Tubulusinnenraum ausgesetzt. Die Reizschwelle der Rezeptorzellen liegt immer bei der eigenen Entladungsfrequenz des Tieres, wobei die Methode der Abstimmung noch nicht erklärt werden kann.
Es gibt zwei Typen von Tubulusrezeptoren. Zum einen die P-Rezeptoren, die empfindlich auf kleine Amplitudenänderungen des elektrischen Feldes reagieren. Sie sind mit effektorischen sowie inhibitorischen Neuronen verschaltet, wobei eine Amplitudenzunahme oder -abnahme entweder zu einer Erregung oder zu einer Hemmung führt. So können, wie bei der lateralen Inhibition in der Wirbeltierretina, (elektrische) Kontraste verstärkt und somit schwache Feldänderungen besser wahrgenommen werden. Bei den P-Rezeptoren erfolgt eine schnelle Adaption auf elektrische Reize.
Der zweite Rezeptortyp sind die T-units. Sie sind Zeitcodierer, die jeweils einen einzelnen Spike absenden, wenn ein elektrisches Feld die Null-Linie zwischen positiv und negativ durchläuft. Bei den T-units erfolgt keine Adaption.
Durch die beiden Rezeptortypen können die Gymnotiden nicht nur zwischen ohmschen und kapazitiven Widerständen in ihrem elektrischen Feld differenzieren, sondern auch Interferenzen mit elektrischen Wechselfeldern von anderen elektrischen Fischen analysieren.
Mormyromasten und Knollenorgane
Die Knollenorgane der Nilhechte (Mormyridae) sind Zeitcodierer, die wie die T-Units der Neuwelt-Messerfische funktionieren. Sie senden auch einzelne Spikes bei einer Durchquerung der Null-Linie des elektrischen Felds. Die Knollenorgane dienen zur Kommunikation mit anderen Fischen.
Die Mormyromasten sind ähnlich aufgebaut wie die Tubulusorgane der Neuwelt-Messerfische, doch differenzierter. Sie besitzen A-Rezeptoren die in die Kammerwand eingebettet und nur mit der apikalen Membran dem Umgebungswasser ausgesetzt sind. Die A-Rezeptoren sind Amplitudencodierer. Darüber hinaus gibt es in einer unter der Hauptkammer liegenden inneren Kammer B-Rezeptoren die einzeln komplett vom Außenwasser umgeben sind (100 % der Membranoberfläche). Diese sind kapazitiv mit dem Wasser gekoppelt und reagieren sehr empfindlich auf Veränderungen der Wellenformen des elektrischen Felds. Sie sind Zeitcodierer. Die Mormyromasten dienen zur Elektrolokalisation.
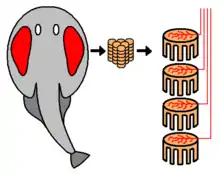
Elektroplax
Elektrische Entladungen werden in elektrischen Organen, den sogenannten Elektroplaxen, erzeugt. Das Elektroplax ist aus Elektrocyten aufgebaut, welche als elektrische Platten zu Säulen zusammengeschaltet sind. Die Elektrocyten werden überwiegend aus Schwanz-, Rumpf- und Halsmuskelzellen gebildet und bestehen aus modifizierten Muskelfasern, die keine Myofibrillen mehr enthalten und deshalb kontraktionsunfähig sind. Jede Elektrocyte entsteht durch Verschmelzung von mehreren Muskelfasern, wodurch sich erklären lässt, wieso die Elektrocyten deutlich größer als Muskelfasern sind.[17] Beim Zitterwels sind die Elektrocyten 20–40 μm dick und ca. 1 mm lang.[18] Mehrere elektrische Organe können pro Individuum vorkommen: so besitzt der Zitteraal beispielsweise drei Organe, die bei gemeinsamer Entladung besonders starke Impulse generieren.
Die eine Seite einer sogenannten elektrischen Platte ist glatt und ähnelt einer neuromuskulären Endplatte. Sie ist mit motorischen Spinalnerven verbunden (innerviert). Diese haben eine sehr hohe Synapsendichte. Die Erzeugung von Elektrizität in den einzelnen Elektrocyten basiert auf der Membranpotentialdifferenz der beiden Seiten der elektrischen Platte.
Die andere Seite ist papillös und mit vielen Kapillaren versehen, die zur Versorgung der elektrischen Zellen mit Sauerstoff und Nährstoffen dienen, da die elektrischen Entladungen viel Energie verbrauchen. Durch die hohe Synapsendichte auf der glatten Seite und viele Acetylcholinrezeptoren in deren postsynaptischen Membranen ist eine blitzartige Invertierung (Umkehrung) des Membranpotentials dieser Seite möglich. Diese erfolgt mit großer Präzision und synchron. Dazu gibt es besondere Neuronen verschiedener Dicke und Länge, die zur Synchronisation dienen (siehe 3.b.iii).
Durch die Umkehrung des Membranpotentials der innervierten Seite besteht eine Potentialdifferenz von bis zu 140 mV zwischen beiden Seiten der Elektrocyte. Damit eine solche Spannung zwischen den beiden Seiten der Zelle aufgebaut werden kann und sie als eine Art „Batterie“ fungiert, sind beide Seiten auf bisher noch unbekannte Weise voneinander isoliert. Da diese 140 mV eine kaum wahrnehmbare Spannung darstellen, werden größere Spannungen durch serielle (hintereinander) Anordnung von vielen tausend elektrischen Platten erzeugt. So addieren sich die Spannungsbeiträge der einzelnen Platten und können mehrere hundert bis fast tausend Volt betragen.
Höhere Stromstärken werden durch die parallele (nebeneinander) Anordnung der elektrischen Platten erreicht. Deswegen sind elektrische Organe bei Fischen entweder großflächig oder trommelförmig aufgebaut. So ist entweder die Erzeugung einer größeren Stromstärke oder Spannung möglich.
Die elektrischen Organe können sich in verschiedenen Körperregionen der Fische befinden und unterschiedliche Größen annehmen, was unter anderem mit der Funktion der elektrischen Entladungen zusammenhängt. Beim Zitteraal dient fast die gesamte Rumpfmuskulatur zur Erzeugung von Elektrizität, wohingegen beim Sterngucker nur einer der sechs Augenmuskeln zum elektrischen Organ wurde.
Man unterscheidet zwischen stark und schwach elektrischen Fischen. Die stark elektrischen Fische (Zitterfische) benutzen ihre Bioelektrizität als Waffe, die schwach elektrischen Fische (Messeraale, Nilhechte – siehe aktive Elektroortung) stoßen elektrische Signale zur Orientierung und Kommunikation aus.
Bei der Entladung wird der Fisch zu einem Dipol, wobei entweder das Kopf- oder das Schwanzende der Plus-Pol ist. Dies hängt von der Lage der innervierten Membran der Elektroplaxen ab. Die Entladung kann entweder mono oder biphasisch ablaufen. Dies hängt von der Membraneigenschaft der papillösen (nicht innervierten) Membran ab: Ist diese nicht chemisch oder elektrisch erregbar, ist die Entladung monophasisch. Ist sie dagegen erregbar, verläuft die Entladung biphasisch. Auf die Entladung des innervierten Membranabschnitts folgt die Entladung des nicht innervierten Membranabschnitts. Die Entladung erhält so zwei Spitzen (Plus und Minus), was vor allem bei schwach elektrischen Fischen zu beobachten ist.
Zentralnervöse Verarbeitung
Die Elektrorezeptoren werden durch Neuronen des peripheren Seitenlinienorgans innerviert. Die einzelnen Rezeptorzellen werden konvergent verschaltet um die elektrische Empfindlichkeit zu erhöhen. Dennoch bleiben die verschiedenen Rezeptortypen (Zeit- und Amplitudencodierung) getrennt, bis sie im Mittelhirn differenziell verschaltet werden. Also der Zeitpunkt der Nulldurchgänge und die Amplitudenzu- oder -abnahme.
Im ELL (electrosensory lateral line lobe), in den die Elektrorezeptoren projizieren, treffen die Signale der verschiedenen Rezeptoren parallel ein und es erfolgt eine topographische Abbildung der elektrosensitiven Haut. Die Kontraste werden durch antagonistische Felder erhöht.
Die Zeitcodierung an verschiedenen Hautstellen wird verglichen und ermittelt, wie sich die Phase des gestörten elektrischen Felds verschiebt. Lokale Veränderungen werden so genau differenziert. Darüber hinaus wird dieses „elektrische Bild“, im Hirn des Fisches (tectum opticum) über das visuelle Bild „gelegt“, ergänzt dieses also.
Neuronale Rückkopplung
Man unterscheidet zwischen negativer und positiver Rückkopplung. Die negative Rückkopplung erfolgt durch antagonistische Erregung durch die Zellen der Körnerschicht (über die Parallelfasern) und sorgt dafür, dass die Elektrorezeptoren nicht auf die eigenen elektrischen Entladungen reagieren und dass eine Adaption bei sich wiederholenden Frequenzen erfolgt. So wird die Rezeptorkapazität für singuläre (neue = wichtige) Signale freigehalten. Bei der positiven Rückkopplung geht es um eine Verstärkung der lokalen elektrischen Erregung und der elektrischen Kontraste. Dies ist sehr wichtig für die Elektrolokation.
Funktionen der aktiven Elektroortung
Elektrische Felder
Das elektrische Feld breitet sich um den Dipol (Fisch) von Kopf zu Schwanzspitze aus (Grafik). Gegenstände, die sich im elektrischen Feld des Fisches befinden, erzeugen eine Amplituden- und Phasenverschiebung seines elektrischen Feldes, da sich ihre elektrische Leitfähigkeit von der des Wassers unterscheidet.
Geringerer elektrischer Widerstand als Wasser (kapazitiv) führt zu einer Verdichtung der lokalen elektrischen Feldlinien. Höherer elektrischer Widerstand (resistiv, ohmsch) führt zu einer Ausdünnung.
Allerdings ist die Reichweite der Elektrolokation auf einen sehr geringen Umkreis, der in etwa der halben Fischlänge entspricht, begrenzt. Die Modulationen des elektrischen Felds durch Objekte sind sehr gering und die Fische müssen direkt und mehrmals an ihnen vorbeischwimmen; trotzdem sind die elektrischen Bilder nur verschwommen.
Die Entfernung zu einem Objekt wird durch die Maximalamplitude und die Kontrastschärfe festgestellt. Weiter entfernte Objekte besitzen eine geringere Maximalamplitude und sind „verschwommener“ (undeutliche elektrische Kontraste). Anhand der kapazitiven Widerstände können die Tiere zwischen belebten und unbelebten Objekten unterscheiden.
Die Mormyride nehmen mit ihren Mormyromasten Differenzen der Signale der A- und B-Rezeptoren wahr und können somit zwischen resistiven und kapazitiven Objekten unterscheiden: Bei resistiven Objekten heben sich beide Signale auf, bei kapazitiven gibt es eine starke Differenz der Signale von A- und B-Rezeptoren. Allerdings nimmt mit höherer Leitfähigkeit des Wassers die Sensitivität der Elektrolokation ab, da die Amplitude des elektrischen Feldes des Fisches gleichzeitig abnimmt. So muss durch positive neuronale Rückkopplung die Modulation der Amplituden „verstärkt“ werden. Außerdem ist die Amplitudenmodulation selbst bei optimalen Verhältnissen sehr gering. Aus diesem Grund wird die Elektrolokation meistens als zusätzliches Hilfsmittel in Kombination mit dem Geruchssinn benutzt, wenn die optischen Sinnesorgane nicht mehr genügend Informationen liefern können (Nacht/trübe Gewässer). So kann man vermuten, dass die kommunikative Funktion der elektrischen Entladungen biologisch bedeutsamer ist als die Lokation.
Orientierung
Zur aktiven elektrischen Orientierung sendet der Fisch selbst ein elektrisches Feld aus und nimmt durch Rezeptoren seine Veränderung wahr. Aktive elektrische Ortung ist dann vorteilhaft, wenn zum Beispiel der Seitenliniensinn des Fisches keine ausreichende Orientierung bietet, wie zum Beispiel in sehr schnell und turbulent fließenden Gewässern, oder da wo visuelle Wahrnehmung schwierig ist (trübe Gewässer, Dunkelheit). Allerdings ist die aktive elektrische Ortung auf einen kleinen Umkreis beschränkt. Die Stärke des elektrischen Feldes nimmt mit der vierten Potenz der Entfernung ab und die Detektionsrate nimmt quadratisch zur Distanz ab.
Es gibt zwei verschiedene Großgruppen der schwach elektrischen Fische, die sich aktiv elektrisch orientieren. Das sind zum einen die Gymnotiden (Messeraale), südamerikanische Süßwasserfische, die sinusförmige elektrische Entladungen erzeugen können. Dafür muss eine große Zahl von elektrischen Zellen gleichzeitig erregt werden. Sie besitzen Tubulusorgane, mit denen sie die Änderung ihrer selbst erzeugten elektrischen Felder registrieren. Zum anderen sind es die Mormyriden, afrikanische Süßwasserfische, die pulsförmige elektrische Entladungen erzeugen, welche mono-, bi- oder triphasisch sein können. Bis zu 100 Entladungen pro Sekunde sind möglich.
Aufgrund der teils sehr hohen Entladungsfrequenz ist die neuronale Verschaltung von der Medulla oblongata, dem verlängerten Mark, zu den Elektroplaxen durch verschieden lange und dicke Axon-Stiele synchronisiert. Diese Synchronisation basiert somit auf dem Prinzip des elektrischen Widerstands. Die dickeren „Leitungen“ besitzen einen geringeren elektrischen Widerstand und verlaufen direkt zu den weiter entfernten Elektroplaxen. Die dünneren leiten elektrische Impulse langsamer und verlaufen „auf Umwegen“ zu den näheren elektrischen Zellen.
Die Mormyriden besitzen Knollenorgane und Mormyromasten zur Elektrorezeption.

Kommunikation
Dadurch, dass parallel die Veränderung von Phasen, Amplituden (Tubulär, Mormyromasten, Knollenorgane) und Gleichspannungspotential (Ampullen) gemessen und diese neuronal verstärkt und kombiniert wird, können die Tiere zwischen Artgenossen und Artfremden, sowie Geschlechtspartnern unterscheiden. Diese Unterscheidung erfolgt anhand der Entladungsform und Wiederholungsrate, da der Grundtyp art- und geschlechtsspezifisch ist und die Amplitude in Zusammenhang mit der Länge der Pausen eine Individualerkennung ermöglicht.
Daran, dass dominante Tiere eine höhere Entladungsfrequenz aufweisen, kann man erkennen, dass so auch eine soziale Interaktion stattfindet.
Zur Balz und in Fällen von Aggression zerhacken die Männchen ihre kontinuierlichen Entladungen zu kurzen Salven (Chirps). Diese Chirps dienen für Weibchen als notwendiger Schlüsselreiz für das Ablaichen. Eine Erhöhung der Sendefrequenz weist jedoch auf Aggression hin.
Der Umkreis der elektrischen Kommunikation beträgt jedoch maximal einen Meter.
Bioelektrizität als Waffe
Stark elektrische Fische werden als Zitterfische bezeichnet. Zitterfische besitzen zwar in der Regel keine tuberösen Elektrorezeptoren, gehören aber zu den elektrischen Fischen und sind somit der Vollständigkeit halber zu erwähnen.
Ein besonderes Beispiel ist der Zitteraal. Sein kompletter Schwanzmuskel (ca. 70 % des Körpers) ist zur Erzeugung von Elektrizität umfunktioniert. Aufgrund dieser Größe sind bis zu 6000 Elektrocyten seriell und parallel miteinander verschaltet, was zu hohen Spannungen führt. Ausgewachsene Exemplare können so mit dem Elektroplax für etwa zwei Millisekunden Spannungen von mindestens 600 Volt, die größten Vertreter bis 860 Volt und 1 Ampere Stromstärke generieren was einer kurzfristigen Leistung von 860 Watt entspricht.[19][20][21] Der Zitteraal kann sowohl starke als auch schwache elektrische Entladungen abgeben, die zum einen dem Beutefang, zum anderen der Revierabgrenzung und dem Auffinden von Fortpflanzungspartnern dienen. Dabei fungiert der Aal als elektrischer Dipol mit dem positiven Pol am Kopf und dem negativen Pol an der Schwanzspitze.
Dabei sind die Tiere vor Stromschlägen durch sich selbst oder andere Individuen durch die isolierende Haut oder zusätzliche Fettschichten um lebenswichtige Organe geschützt.[22]
Zitterrochen können elektrische Entladungen von 60 bis 230 V und über 30 A ausführen, um ihre Opfer zu lähmen. Beim Gefleckten Zitterrochen kann die Stromstärke 50 A erreichen, wobei dieser extrem hohe Wert mit einer Spannung von 'nur' 60 V nachgewiesen wurde.
Quellen
- T. H. Bullock, C. D. Hopkins, A. N. Popper, R. R. Fay: Electroreception. Springer Handbook of Auditory Research. Springer Verlag, Berlin 2005, ISBN 0-387-23192-7.
- R. Eckert: Tierphysiologie. Thieme Verlag, Stuttgart 2002.
- W. Heiligenberg: Neural nets in electric fish. MIT Press, Cambridge, MA 1991.
- G. Heldmaier, G. Neuweiler: Vergleichende Tierphysiologie. 1. Band, Springer Verlag, Berlin 2003.
Sekundärliteratur
- R. W. Turner, L. Maler, M. Burrows: Electroreception and electrocommunication. In: J Exp Biol. Band 202, 1999, S. 1167–1458.
- G. von der Emde: Electroreception. In: D. H. Evans (Hrsg.): The Physiology of Fishes. 2. Auflage. CRC Press, Boca Raton (Florida) 1998, S. 313–343.
Einzelnachweise
- Günther Sterba: Süsswasserfische der Welt. 2. Auflage. Urania, Leipzig/Jena/Berlin 1990, ISBN 3-332-00109-4.
- Gerard H. Markx, Burçak Alp, Alastair McGilchrist: Electro-orientation of Schizosaccharomyces pombe in high conductivity media. In: Journal of Microbiological Methods. 50, Nr. 1, 2002, S. 55–62, doi:10.1016/S0167-7012(02)00012-X.
- Y. Dai, S. E. Law: Modeling the transient electric field produced by a charged pollen cloud entering a flower. In: Industry Applications Conference, 1995. Thirtieth IAS Annual Meeting, IAS '95., Conference Record of the 1995 IEEE.
- Yiftach Vaknin, S. Gan-Mor, A. Bechar, B. Ronen, D. Eisikowitch: The role of electrostatic forces in pollination. In: Plant Systematics and Evolution. 222, Band 1–4, 2000, S. 133–142.
- Yiftach Vaknin, Samuel Gan-mor, Avital Bechar, Beni Ronen, Dan Eisikowitch: Are flowers morphologically adapted to take advantage of electrostatic forces in pollination? In: New Phytologist. 152, Nr. 2, 2001, S. 301–306, doi:10.1046/j.0028-646X.2001.00263.x.
- Dominic Clarke: Detection and learning of floral electric fields by bumblebees. In: Science. Online-Vorabveröffentlichung vom 21. Februar 2013, doi:10.1126/science.1230883
- S. Edward Law: Agricultural electrostatic spray application: a review of significant research and development during the 20th century. In: Journal of Electrostatics. 51, 2001, S. 25–42, doi:10.1016/S0304-3886(01)00040-7.
- Daniel Robert u. a.: News and events. In: Eur J Ophthalmol. 10, Nr. 2, 2000, S. 132–136.
- Victor Manuel Ortega-Jimenez, Robert Dudley: Spiderweb deformation induced by electrostatically charged insects. In: Scientific Reports. 3, Nr. 2108, Juli 2013, doi:10.1038/srep02108.
- David Bodznick, R. Glenn Northcutt: Electroreception in lampreys: evidence that the earliest vertebrates were electroreceptive. In: Science 212, Nr. 4493, 1981, S. 465–467.
- Ad J. Kalmijn: Electro-orientation in sharks and rays: theory and experimental evidence. (PDF) No. SIO-Ref-73-39. SCRIPPS Institution of Oceanography La Jolla California, November 1973.
- Marcelo Camperi u. a.: From morphology to neural information: The electric sense of the skate. In: PLOS Computational Biology. Band 3, 2007, S. 1083–1096, e113. doi:10.1371/journal.pcbi.0030113.
- Wilfried Westheide, Gunde Rieger: Spezielle Zoologie. Teil 2: Wirbel- oder Schädeltiere. Springer-Verlag, 2014, ISBN 978-3-642-55436-0, S. 224 (eingeschränkte Vorschau in der Google-Buchsuche [abgerufen am 15. Dezember 2017]).
- Kurt Fiedler: Lehrbuch der Speziellen Zoologie, Band II, Teil 2: Fische. Gustav Fischer Verlag Jena 1991, ISBN 3-334-00339-6, S. 112–119.
- Nicole U. Czech-Damal u. a.: Electroreception in the Guiana dolphin (Sotalia guianensis). In: Proceedings of the Royal Society B. Band 279, Nr. 1729, 2011, S. 663–668, doi:10.1098/rspb.2011.1127
- G. N. Akoev, N. O. Vol'pe: Analysis of the mechanism of perception of temperature stimuli by the ampullae of Lorenzini of Black Sea skates. In: Fiziol Zh SSSR Im I M Sechenova. Band 64, 1978, S. 1681–1688.
- G. A. Unguez, H. H. Zakon: Phenotypic conversion of distinct muscle fiber populations to electrocytes in a weakly electric fish. In: J Comp Neurol. Band 399, 1998, S. 20–34. PMID 9725698
- T. Schikorski: Funktionelle Neuroanatomie des elektromotorischen Systems des Zitterwelses Malapterurus electricus. Dissertation. Fachbereich Biologie der Johann Wolfgang Goethe-Universität, Frankfurt am Main 1993, DNB 94312669X.
- Amazon Shocker Facts. In: Nat Geo WILD. 6. Juni 2014 (nationalgeographic.com [abgerufen am 9. Dezember 2016]).
- Monster Eel, Monster Shock? (Nicht mehr online verfügbar.) In: Nat Geo TV Blogs. 21. Juli 2014, archiviert vom Original am 10. Dezember 2016; abgerufen am 9. Dezember 2016. Info: Der Archivlink wurde automatisch eingesetzt und noch nicht geprüft. Bitte prüfe Original- und Archivlink gemäß Anleitung und entferne dann diesen Hinweis.
- How do electric eels generate a voltage and why do they not get shocked in the process? In: Scientific American. (scientificamerican.com [abgerufen am 9. Dezember 2016]).
- Kathrin Passig, Aleks Scholz, Kai Schreiber: Das neue Lexikon des Unwissens: Worauf es bisher keine Antwort gibt. Rowohlt E-Book, 2011, ISBN 978-3-644-11011-3 (eingeschränkte Vorschau in der Google-Buchsuche [abgerufen am 6. Mai 2016]).
Weblinks
- C. C. Bell: Mormyromast electroreceptor organs and their afferent fibers in mormyrid fish. III. Physiological differences between two morphological types of fibers. In: Journal of Neurophysiology. Band 63, Nr. 2, 1990, S. 319–332, doi:10.1152/jn.1990.63.2.319.
- A. Roth: Zur Funktionsweise der Elektrorezeptoren in der Haut von Welsen (Ictalurus): Der Einfluß der Ionen im Süßwasser. In: Zeitschrift für vergleichende Physiologie. Band 75, 1971, S. 303–322, doi:10.1007/BF00340683.
- Johannes Günther, Gaby Markwirth, Olaf Zimmer: Elektrorezeptoren.
- Otto Schlappal, Andreas Schwarzkopf, Jan Trautmann, Torsten Roßmann: Elektrische Fische: Signalerzeugung & Rezeption. Funktion, Aufbau und Einsatz elektrischer Organe. Technische Universität, Darmstadt 2008 (PDF (Memento vom 18. September 2017 im Internet Archive) – 746 kB).
- Sönke von den Berg: Gnathonemus petersii oder der Elefantenrüsselfisch. (Nicht mehr online verfügbar.) In: Elektr. Fische Teil 1/5. 2008, archiviert vom Original am 25. Oktober 2018.