Biotopverbund
Der Biotopverbund oder die Biotopvernetzung ist die Schaffung eines Netzes von (Einzel-)Biotopen, welches das Überleben von Arten sichert. Der Biotopverbund ist gegeben, wenn ein funktionaler Kontakt zwischen Biotopen (Lebensräumen) besteht, der eine Vernetzung zwischen Populationen von Organismen ermöglicht. Er funktioniert dann, wenn die zwischen gleichartigen Lebensräumen liegende Fläche für Organismen überwindbar ist, oder durch Prozesse wie den Transport durch Weidetiere überbrückt wird, so dass ein beidseitiger Individuenaustausch möglich ist.
In Deutschland sind Biotopverbund und Biotopvernetzung u. a. durch das Bundesnaturschutzgesetz (§21) angestrebtes Ziel.[1] In Bayern wird seit dem Volksbegehren „Artenvielfalt & Naturschönheit in Bayern“ im Bayerischen Naturschutzgesetz in Artikel 19 ein konkret zu erreichender Flächenanteil Biotopverbund festgeschrieben.[2]
Bedeutung des Verbunds im Lebenszyklus
Vor allem im zoologischen Bereich wird für die Bedeutung des Biotopverbunds für definierte Einzelarten dessen Bedeutung im Lebenszyklus der Art zugrunde gelegt. Unterschieden werden[3][4]:
- Arten, die einen Biotopverbund für ihr normales Aufenthalts-, Jagd- oder Nahrungsrevier benötigen. Ohne Verbund werden ansonsten besiedelbare Bereiche als Lebensraum entwertet und/oder das Minimumareal für eine lebensfähige Population unterschritten. Dies betrifft vor allem Arten mit sehr großem Raumanspruch, meist als Räuber lebende Wirbeltiere (z. B. der Wolf oder die Wildkatze)
- Arten, die einen Biotopverbund für regelmäßige Wanderungen (Migrationen) benötigen. Ohne Verbund sind räumlich getrennte Teillebensräume voneinander isoliert. Betroffen sind z. B. Wanderungen zwischen Sommer- und Winterhabitat oder zwischen Fortpflanzungs- und Nahrungshabitat. Die Wanderungen können großräumig (z. B. Zwischenquartiere für Zugvögel oder Fledermäuse), auf mittlerer Raumebene (z. B. Sommer- und Winterhabitate des Rothirschs) oder kleinräumig (z. B. Amphibienwanderungen) sein.
- Arten, die einen Biotopverbund zur Ausbreitung (Dispersion) benötigen, um neue oder frei gewordene Habitat"inseln" kolonisieren zu können. Bei fehlendem Verbund sterben sie aus, indem über einen längeren Zeitraum nach und nach alle Lokalpopulationen nacheinander erlöschen, entweder aus zufälligen Gründen (stochastische Schwankungen), infolge von Sukzessionsvorgängen (Pionierarten) oder wegen großräumiger Arealverschiebungen (globale Erwärmung). Diskutiert (aber bisher sehr selten nachgewiesen) wird auch eine genetische Verarmung aufgrund mangelhaften Allelaustauschs.
Während der erste und der zweite Fall relativ leicht anschaulich sind, sind für den dritten Fall weitere Begründungen erforderlich. Meist wird angenommen, dass Beschränkungen der Dispersion kurzfristig keine offensichtlichen Auswirkungen zu besitzen scheinen, dass sich aber langfristig die Folgen mangelnden Biotopverbunds vor allem daraus ergeben werden. Bei Pflanzenarten ist überhaupt nur der dritte Weg bedeutsam.
Ökologische Grundlagen
Die Forderung nach einem Biotopverbund beruht einerseits auf Modellaussagen, die aus der ökologischen Theorie abgeleitet sind, andererseits auf empirischen Beobachtungen, d. h. Fallstudien, bei denen das Überleben von Populationen in mehr oder minder isolierten Habitaten untersucht wurde.
Inseltheorie
(vgl. Inselbiogeographie) Eine wichtige Grundlage für den Biotopverbund stellt die Inseltheorie (eigentlich: Gleichgewichts-Theorie der Biogeographie von Inseln) dar. Es handelt sich um eine einflussreiche ökologische Theorie, die die amerikanischen theoretischen Ökologen Robert H. McArthur und Edward O. Wilson zuerst im Jahr 1963[5] aufgestellt und 1967 verallgemeinert haben.[6] Die Forscher betrachteten die Artenzahlen auf mehr oder weniger isoliert liegenden ozeanischen Inseln. Sie kamen auf ein in einer mathematischen Formel darstellbares Abhängigkeitsmuster, das sie biologisch als ein Gleichgewicht zwischen Einwanderung und (lokalem) Aussterben interpretieren konnten. Lokale Populationen unterliegen einem (stochastischen) Aussterberisiko, welches abhängig von der Populationsgröße ist, welche wiederum von der Inselgröße (über die Tragfähigkeit) abhängt. Kleine Populationen sterben nach einer mehr oder weniger langen Periode zufallsbedingt aus (zufällige Populationsschwankungen). Die Insel stellt danach einen freien Lebensraum für die Art dar. Sie kann schließlich neu besiedelt werden, wenn sie von einwandernden Individuen dieser Art entdeckt wird. Diese Kolonisierung hängt von den biologischen Eigenschaften der kolonisierenden Art, aber vor allem auch von der Isolation der Insel ab, die sich (neben ihrer Gestalt und einigen anderen Parametern) vor allem durch ihre Entfernung ergibt. Die tatsächliche Artenzahl einer Insel ergibt sich danach als ein Gleichgewichtszustand zwischen Aussterben und Neubegründung von Populationen. Der Artenbestand der Insel wechselt dabei (Artenwechsel, engl. species turnover). Die Artenzahl hängt von der Inselgröße und ihrer Entfernung zu anderen Inseln (oder einem Kontinent) ab.
Die Inseltheorie wurde bereits früh von ozeanischen Inseln auf isoliert liegende besondere Biotope übertragen, die als Habitat-Inseln in einem für das Überleben der betrachteten Art feindlichen "Ozean" interpretiert werden können. Daraus ergeben sich eine Reihe von Vorhersagen, die teilweise experimentell bestätigt werden konnten.
- Nimmt die Größe einer Habitat-Insel ab, sterben zahlreiche Arten aus, auch wenn sich die Habitatqualität der verbliebenen Fläche überhaupt nicht verschlechtert ("Relaxation"). In der Übergangszeit hat die Insel einen Arten-Überhang, der nach und nach zum Aussterben verdammt ist. Welche Arten tatsächlich überleben, hängt teilweise vom Zufall ab. Im Naturschutz spricht man auch von "Aussterbeschuld" (engl.: extinction debt). Zur Anwendung vgl. z. B.[7] und.[8]
- Sind die verbliebenen Habitat-Inseln so entfernt voneinander, dass eine Neukolonisation "leerer" Inseln unmöglich oder höchst unwahrscheinlich ist (Kolonisationsrate Null), sterben über kurz oder lang alle Arten auf allen Inseln aus (Sie überleben nur auf sehr großen Inseln und Kontinenten. Diese sind so groß, dass sie mehrere unabhängige Populationen besitzen, und ihre Populationsgröße ist so hoch, dass das stochastische Aussterberisiko auf nahe Null fällt.)
- Kann die Kolonisationsrate von Habitat-Inseln erhöht werden, können mehr Arten überleben, obwohl weder die Größe noch die Qualität der Habitat-Inseln erhöht wurde.
Metapopulationen
Eine Metapopulation beschreibt ein Netz von Einzel-Populationen (Subpopulationen), die teilweise, aber nicht vollständig, voneinander isoliert sind. Die Subpopulationen wechselwirken miteinander durch den Austausch von Individuen. Die o. g. Aussagen der Inseltheorie können genauso gut über einen populationsbiologischen Ansatz hergeleitet werden. Es ergeben sich aber darüber hinausgehende Hypothesen.
Der finnische Populationsbiologe Ilkka Hanski hat eine einflussreiche Theorie aufgestellt, die berücksichtigt, dass die Populationsgröße einer Art bei der Besiedlung von Inseln oder Teilbiotopen (meist engl.: habitat patches) zweimal eingeht; sie erhöht die Kolonisationsrate und vermindert die Aussterberate.[9] In seinem Modell ergibt sich dadurch eine zweigipflige ("bimodale") Häufigkeitsverteilung: Wenige häufige Arten, von ihm "Kernarten" genannt, sind quasi auf allen Inseln präsent. Die meisten Arten ("Satellitenarten") sind seltener, als man bei rein zufälliger Verteilung erwarten würde. Durch diesen Zusammenhang steigt die Artenzahl mit zunehmender Anzahl von Inseln langsamer an. Nur die "Kernarten" sind auf nahezu allen Inseln präsent.
Der niederländische Ökologe Pieter J. den Boer hat in langjährigen Forschungsarbeiten den Einfluss der Dispersion auf die Laufkäferfauna niederländischer Heidefragmente untersucht.[10][11] Bei diesen räuberischen, bodenlebenden Käfern gibt es flugfähige Arten, nicht flugfähige Arten und Arten, bei denen ein Teil der Individuen flugfähig ist (flügeldimorphe Arten). Er konnte zeigen, dass in isolierten Heidefragmenten die Artenzahl langfristig abnimmt. Außerdem nimmt bei den verbleibenden der Anteil der flugfähigen Individuen ab. Den Boer interpretiert dies als evolutionären Trend aufgrund der Verinselung. Durch die Isolation der Heidefragmente nimmt die Wahrscheinlichkeit erfolgreicher Einwanderungen ab. Damit sind Arten kurzfristig im Vorteil, die nicht mehr in den (energetisch kostspieligen) Flugapparat investieren. Langfristig sind damit allerdings fast alle Arten auf lange Sicht zum Aussterben verdammt.
Bedeutsam im Metapopulations-Ansatz ist auch die Erhaltung der genetischen Variabilität der Arten. Auf kleinen Inseln und in durch wenige Kolonisatoren neu begründeten Populationen ist die Variabilität notwendigerweise viel geringer als in großen ("Gründereffekt"). Sie wird durch immigrierende Individuen aus anderen Populationen erhöht. Dieser Effekt ist auch dann bedeutsam, wenn diese Einwanderer eine Insel erreichen, die von der Art noch besiedelt ist. Der Verlust an genetischer Variabilität erhöht deutlich das Aussterberisiko einer Art, weil ihre Plastizität, auf Lebensraumveränderungen zu reagieren, verloren gehen kann. Steigt die Populationsgröße nach einem Beinahe-Aussterben wieder an, ist diese Population damit viel anfälliger als die ursprüngliche ("genetischer Flaschenhals").
Für sehr mobile Tierarten, z. B. viele Vogelarten, ist das Metapopulationsmodell in der Praxis bedeutungslos. Hier können Individuen problemlos zwischen entfernten Teilhabitaten wechseln. Sie bilden durch den häufigen Austausch eine einzige große Population.
Randeffekte
Randeffekte beschreiben den Einfluss der umliegenden Landschaft auf Biotopinseln. Im Gegensatz zu ozeanischen Inseln sind Biotopinseln ja in eine wesentlich ähnlichere Umgebung eingebettet. Randeffekte treten hier in mehreren Arten auf. Zum einen wirken intensive Flächennutzungen auch in geschützte Biotop-Inseln hinein, indem z. B. Pestizide oder Dünger eingeweht und eingeschwemmt werden können. Deshalb ist der Wert sehr kleiner oder schmaler Biotop-Inseln in der Praxis manchmal stark vermindert. Eine andere Art Randeffekt geht vom Artenbestand der Agrarlandschaft aus, der (im Gegensatz zu den ozeanischen Arten zwischen echten Inseln) durchaus auch die Biotop-Inseln kolonisieren kann. Randeffekten versucht der Naturschutz durch die Einrichtung von Pufferzonen entgegenzuwirken. Im Biotopverbund müssen sie berücksichtigt werden, um die Effektivität von Biotopinseln und -korridoren beurteilen zu können.
Randeffekte auf Populationsebene können vom Naturschutz unter Umständen beabsichtigt sein. In diesen Fällen soll das (zu schützende oder neu zu schaffende) Netz von Linienbiotopen nicht in erster Linie naturnahe Biotop-Inseln miteinander vernetzen, sondern gezielt den Wert der dazwischen liegenden Agrarlandschaft verbessern, indem hier Arten der Agrarlandschaft selbst Refugien oder Teilhabitate finden (vgl. z. B.[12]) Obwohl dieses Ziel legitim und praktikabel sein kann, verschwimmt die Argumentation in der Praxis manchmal dadurch so, dass kaum noch auszumachen ist, was das eigentliche Ziel der Biotopvernetzung sein soll.
Randeffekte können in größerem Maßstab wirksam sein als oft intuitiv angenommen. Untersuchungen in Neuseeland haben gezeigt, dass selbst bei kleinen bodenlebenden Käfern noch in einem Kilometer Entfernung von der Grenze eines Schutzgebiets nachweisbare Effekte auf die Fauna auftraten.[13] Stärkere Effekte sind naturgemäß bei Arten, die die Schutzgebietsgrenzen regelmäßig überschreiten, zu erwarten, z. B. Carnivoren.[14] Neben den negativen sind auch die positiven Randeffekte schon tatsächlich nachgewiesen worden: Es kann zu einer Anreicherung angrenzender Lebensräume mit Arten kommen.[15] In Fließgewässern versucht man in Deutschland, sich diesen Effekt unter dem Namen "Strahlwirkung" zunutze zu machen, wobei belastbare empirische Belege zur Wirksamkeit aber noch ausstehen.
Formen des Biotopverbunds
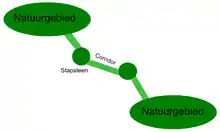
Der Biotopverbund definiert sich also über seine Wirksamkeit auf ausgewählte Zielarten. Streng genommen werden nicht Biotope vernetzt, sondern Populationen. Der Biotopverbund muss also je nach Biologie der betrachteten Art (v. a. ihrer Mobilität und Ausbreitungsbiologie) andere Anforderungen erfüllen. Für mobile Arten, insbesondere flugfähige oder solche mit flugfähigen Lebensstadien (v. a. Vögel, die meisten Insekten, Pflanzen mit windverbreiteten Samen) wird ein Verbund meist über Trittsteinbiotope angestrebt. Dies sind kleine, nicht notwendigerweise miteinander verbundene, Biotop-"Inseln" innerhalb der umliegenden Landschaft. Ihre Entfernung sollte sich nach der Mobilität der Zielart bemessen. Für immobile, bodengebundene Arten (z. B. viele Säugetiere, aber auch Pflanzen des Waldbodens ohne effiziente Fernverbreitungsmechanismen) wird ein Biotopverbund über Biotopkorridore angestrebt, d. h. linienhafte Strukturen, die die zu verbindenden Zielbiotope physisch miteinander verbinden. Dies können z. B. Heckenstreifen zwischen zwei Wald-"Inseln" oder eine oder mehrere Wald-"Inseln" (sogenannte Waldbrücken) zwischen großen Waldgebieten sein.[16] Bedeutsame Aufgaben für den Biotopverbund sind darüber hinaus unüberwindbare Landschaftsbarrieren, vor allem große Straßen (Autobahnen).[17] Diese sind auch für recht mobile, aber nicht flugfähige Arten, für die die normale Agrarlandschaft keine Zäsur darstellt, meist unüberwindliche Hindernisse. Dies ist besonders bedeutsam für große Säugetierarten. Entgegen der Intuition können solche Zäsuren je nach Ausgestaltung aber auch für flugfähige Arten (z. B. Fledermäuse) als Barrieren wirken. Ein Biotopverbund wird hier über besondere Verbindungselemente (z. B. Grünbrücken) auf Landschaftsebene angestrebt.

Der Bau von Grünbrücken kann im Zusammenhang mit einer Strategie des Biotopverbunds gesehen werden, die nicht so sehr bei den Inseln, sondern mehr bei den dazwischen liegenden Räumen ansetzt. Es soll durch gezielte Maßnahmen die Durchlässigkeit der Landschaft zwischen den Biotopinseln erhöht werden (meist engl. "connectivity"). Obwohl diese Strategie nach den Theorien der Ökologie gut fundiert wäre, stellen sich in der Praxis ganz erhebliche Probleme, wenn z. B. die großflächige Extensivierung einer landwirtschaftlichen Bodennutzung gefordert wird. In der Praxis spielt dieser Ansatz (außer bei der Überwindbarkeit linearer Hindernisse, v. a. Verkehrstrassen) noch keine Rolle.
Ein Sonderfall des Biotopverbunds, der meist abseits der sonstigen Diskussion betrachtet wird, ist die Aufhebung der Isolation von Fließgewässer-Abschnitten, z. B. durch Wehre, Wasserkraftanlagen, Stauseen oder verrohrte Gewässerstrecken. Man spricht hier von der „ökologischen Durchgängigkeit“ der Gewässer. Fehlt die Durchgängigkeit, können z. B. Wanderfischarten Gewässeroberläufe nicht erreichen, die eigentlich als Lebensraum geeignet wären. Auch für wirbellose Arten können sich Probleme ergeben, weil nach Aussterbeereignissen Teilstrecken des Gewässers unter Umständen nicht neu besiedelt werden können. Durch die Verabschiedung der europäischen Wasserrahmenrichtlinie haben sich die Mitgliedsstaaten verpflichtet, die ökologische Durchgängigkeit aller Fließgewässer wiederherzustellen, wo dies möglich ist. Typische Maßnahmen zur Herstellung der Durchgängigkeit umfassen z. B. den Bau von Fischaufstiegsanlagen (Fischtreppen) an Wasserkraftanlagen und Wehren, der Bau von Umgehungsgerinnen („Bypässen“) oder der Rückbau von Verrohrungen und Abstürzen (Sohlsprüngen).
Anwendung in Naturschutz und Landschaftsplanung
Die Forderung nach einer Biotopvernetzung wurde in der Naturschutzdiskussion erst seit den 1980er Jahren stärker. Vorher hatte der Naturschutz sich auf die Erhaltung besonders wertvoller Einzelbiotope konzentriert. Aufgrund seines geringen gesellschaftlichen Einflusses hatte er wohl in der Praxis auch keine andere Wahl. Die Bedeutung des Biotopverbunds wurde den Naturschützern vor allem durch die abnehmende Lebensraumeignung der "normalen" Agrarlandschaft aufgrund der intensiveren Produktionsweisen bewusst. Dadurch wurde immer deutlicher, dass die meisten Arten in kleinen Naturschutzgebieten allein auf Dauer nicht zu erhalten sein werden. In der internationalen Diskussion wurde das Thema bei den Bemühungen zur Erhaltung bedrohter natürlicher Lebensräume, z. B. des tropischen Regenwalds, akut. Hier wurde in der Fachwelt eine Debatte um die Größe und den Zuschnitt der Schutzgebiete geführt, die unter dem Schlagwort SLOSS ("single large or several small") zusammengefasst wird. Heute ist die Bedeutung des Biotopverbunds im amtlichen und ehrenamtlichen Naturschutz weithin anerkannt.
Die Erfolge bei den praktischen Bemühungen um die Umsetzung der Biotopvernetzung werden nach wie vor kontrovers diskutiert. Die Biotopvernetzung wurde vor allem Anfang der 1990er Jahre zu einem Mode- und Schlagwort, durch die das eigentliche Anliegen eher verunklart worden ist. Insbesondere ist eine Tendenz zu erkennen, in der planerischen Umsetzung die Tatsache aus den Augen zu verlieren, dass eine Vernetzung von "Biotopen" eigentlich nur eine abgekürzte Schreibweise für die Vernetzung von Populationen von Tier- und Pflanzenarten darstellen soll. Durch die Definition abstrakter "Biotoptypen" ist der Zusammenhang in der Praxis gelegentlich verloren gegangen.
Förderung von Linienbiotopen in der Agrarlandschaft
Eine besondere Bedeutung bei der Biotopvernetzung haben Linienbiotope in der Ackerlandschaft. Zu den Linienbiotopen zählen Ackerrandstreifen, Raine, Lesesteinwälle, Böschungen, Wege und Straßenränder, Hecken, Alleen und Fließgewässer. Linienbiotope tragen besonders in einer stark ausgeräumten Landschaft mit geringem oder fehlendem Wald- und Grünlandanteil zur Mannigfaltigkeit und Vernetzung der inselartigen Biotope bei.
Förderung des großräumigen Biotopverbunds
Eine nationale Strategie zum Biotopverbund wurde in Deutschland zuerst durch einen Beschluss der Ministerkonferenz für Raumordnung am 27. November 1992 angestrebt. Eine Arbeitsgruppe unter Beteiligung des Bundesamts für Naturschutz erarbeitete eine fachliche Strategie.[18] In der Europäischen Union wird das Thema unter dem Schlagwort „green infrastructure“ seit 2008 vertiefend debattiert (eine Zusammenfassung der bisherigen Ansätze unter[19]). Grünes Band Deutschland stellt ein aktuelles Projekt zur großräumigen, linienhaften Verbindung verschiedener Biotope dar. Es ist Bestandteil der europäischen Initiative Grünes Band Europa.[20] Die Niederlande streben seit ca. 20 Jahren ein nationales Biotopverbundnetz unter dem Namen Ecologische Hoofdstructuur(EHS) an.[21] In Frankreich werden seit 2008 zwei zusammenhängende Schutzgebietsnetze unter dem Titel Trame verte et bleue (TVB) angestrebt.
Der Naturschutzbund Deutschland NABU stellte 2007 die nationale Strategie eines Bundeswildwegeplans auf, um vor allem der Zerschneidungswirkung der Autobahnen entgegenzuwirken.[22] Der Bund für Umwelt und Naturschutz Deutschland BUND stellte gezielt für die Zielart Wildkatze einen Wildkatzenwegeplan auf,[23] dessen Realisierung er in mehreren Projekten, z. B. in Thüringen, anstrebt.
Politische Umsetzung
Im deutschen Bundesland Baden-Württemberg legte der amtierende Landesminister für Ländlichen Raum und Verbraucherschutz in Baden-Württemberg Alexander Bonde (Bündnis 90/Die Grünen Baden-Württemberg) im April 2012 ein Konzept zur Biotopvernetzung im Land vor.[24] Seit 1995 besteht in Bayern das Bayern-Netz-Natur.
Nachteile und Kritik des Biotopverbunds
Der Biotopverbund als Strategie des Naturschutzes gilt allgemein als fachlich gut fundiert und anerkannt. Dennoch gibt es in zahlreichen Einzelfällen Kritik an der Anwendung.
Eine eher generelle Kritik kommt zu dem Schluss, dass der Erfolg zahlreicher bisher durchgeführter Maßnahmen der Biotopvernetzung zweifelhaft oder nicht nachweisbar gewesen ist. Aufgrund der Knappheit von Naturschutzmitteln seien diese verschwendet worden und besser in den direkten Erhalt hochwertiger Biotope investiert worden. Die meisten Naturschützer sehen diese Kritik als nicht berechtigt an. Dennoch ist darauf hinzuweisen, dass der konkrete Erfolg von biotopvernetzenden Maßnahmen (wie aller Naturschutzmaßnahmen) überprüft und die Maßnahmen ggf. verbessert werden müssen. Obwohl es in sorgfältigen Studien möglich war, den Effekt von Korridoren nachzuweisen,[25] ist das tatsächliche Scheitern zahlreicher Maßnahmen, die den Biotopverbund stützen sollten, leider gut dokumentiert.
Probleme der Biotopvernetzung sind darüber hinaus in Einzelfällen plausibel.
- Durch die Vernetzung können sich Seuchen und Krankheitserreger schneller ausbreiten. In Fließgewässern wird z. B. die Beibehaltung von Wehren gefordert, die Reliktvorkommen des heimischen Edelkrebses (Astacus astacus) bisher vor der Krebspest geschützt haben.
- Die Biotopkorridore können die Ausbreitung unerwünschter Arten, z. B. Neophyten und Neozoen, genauso fördern wie diejenige der Zielarten. Sie können sich dadurch homogenisierend auswirken.
- Biotopvernetzung kann die Abwanderung von Individuen in suboptimale Lebensräume fördern, die als Populationssenken wirken, und damit eine zu erhaltende Population schwächen.
- Die tatsächliche Ausbreitungsbiologie und Ausbreitungsgeschwindigkeit zahlreicher Arten ist unzureichend bekannt. Damit ist das Design von Biotopkorridoren oft nur auf Grundlage von Vermutungen durchführbar. Unter Umständen werden in Einzelfällen Lebensräume vernetzt, die tatsächlich keiner Vernetzung bedürfen.
Biotopvernetzung und Klimawandel
Als zusätzliches Argument für die Biotopvernetzung wird in der Debatte seit etwa 2005 die Milderung (meist engl. "mitigation") von Auswirkungen des menschengemachten Klimawandels angeführt.[26][27] Durch den Klimawandel verändern sich zahlreiche Biotope so, dass sie vermutlich einen Teil ihres heutigen Artenbestands verlieren werden. Da das Verbreitungsgebiet fast aller Arten gut nachweisbar mit klimatischen Faktoren korreliert, ist anzunehmen, dass sich das Gesamtareal vieler Arten verschieben wird. Sie verlieren mehr oder weniger große Teile ihres bisherigen Verbreitungsgebiets; sie könnten dies aber durch Erweiterung an anderer Stelle unter Umständen wettmachen, z. B. indem sich ihr Areal nordwärts oder bergauf verschiebt. Dies setzt freilich voraus, dass sie dieses neue (zunächst nur potentielle) Areal auch tatsächlich erreichen können.
In diesem Zusammenhang wird über eine Stärkung des Biotopverbunds debattiert, um bedrohten Arten solche Ausweich- und Wanderkorridore bereitzustellen. Tatsächliche Maßnahmen sind in diesem Zusammenhang bisher nicht erfolgt. Ob diese überhaupt erfolgversprechend wären, wird in der Forschung kontrovers debattiert. Eines der ersten Projekte, das Biotopverbund und Klimafolgen zusammenführen soll, wird derzeit in den Niederlanden vorbereitet, der West European climate corridor am Rhein (Province Gelderland).
Literatur
- Uwe Wegener (Hrsg.): Naturschutz in der Kulturlandschaft, Schutz und Pflege von Lebensräumen. Ulm 1998, ISBN 3-437-35250-4.
- E. Jedicke (Hrsg.): Biotopverbund: Grundlagen und Maßnahmen einer neuen Naturschutzstrategie. 2. Auflage. Ulmer Verlag, Stuttgart 1994, ISBN 3-8001-3324-5.
Weblinks
- Feldrandhecke selbst anlegen
- Bundesministerium für Umwelt, Naturschutz und nukleare Sicherheit: Das Bundesprogramm Wiedervernetzung. Stand: 16. Oktober 2014.
- Bundesamt für Naturschutz: Kohärenz und Biotopverbund.
- Sielmanns Biotopverbund Bodensee, Heinz Sielmann Stiftung
- Sielmanns Biotopverbund Harz-Eichsfeld-Werratal, Heinz Sielmann Stiftung
- Andreas Seiler: Lebensraumkorridore in Europa. Was machen unsere Nachbarn? IENE Infra Eco Network Europe. April 2010.
Einzelnachweise
- § 21 BNatSchG
- BayNatSchG: Art. 19 Biotopverbund, Biotopvernetzung, Arten- und Biotopschutzprogramm - Bürgerservice. Abgerufen am 17. Februar 2020.
- DJV-Projekt: "Überwindung von Barrieren". In: jagdnetz.de. Deutscher Jagdschutz-Verband e. V. (DJV) Vereinigung der deutschen Landesjagdverbände, abgerufen am 6. Februar 2011.
- H. Reck, K. Hänel, M. Hermann, J. Sachteleben: Verbände-Vorhaben „Überwindung von Barrieren“. Zielarten des überörtlichen Biotopverbundes. Zeigerarten für Zerschneidung und Verinselung. Vorentwurf. (PDF; 303 kB) (Nicht mehr online verfügbar.) September 2007, archiviert vom Original am 8. März 2012; abgerufen am 6. Februar 2011.
- Robert H. MacArthur, Edward O. Wilson: An Equilibrium Theory of Insular Zoogeography. Evolution. Band 17, Nr. 4, Dezember 1963, S. 373–387 (englisch, unm.edu [PDF; 1,6 MB]).
- Robert H. MacArthur, Edward O. Wilson: The Theory of Island Biogeography. Princeton University Press, Princeton 1967.
- Ilkka Hanski: Extinction debt in boreal forests. (PDF)
- Aveliina Helm u. a.: Extinction debt in Estonian calcareous grasslands ("alvars"). (PDF)
- Ilkka Hanski: Dynamics of regional distribution: the core and satellite species hypothesis. In: Oikos. 38, Copenhagen 1982, S. 210–221.
- P. J. Den Boer: Dispersal power and survival : carabids in a cultivated countryside. (= Miscellaneous papers. 14). Landbouwhogeschool, Wageningen 1977, OCLC 923343497.
- P. J. Den Boer: Density limits and survival of local populations in 64 carabid species with different powers of dispersal. In: Journal of Evolutionary Biology. 3(1/2), Basel 1990, S. 19–48.
- Biotopvernetzungskonzept Baden-Württemberg (landwirtschaft-mlr.baden-wuerttemberg.de)
- Robert M. Ewers, Raphael K. Didham: Pervasive impact of large-scale edge effects on a beetle community. In: Proceedings of the National Academy of Science (PNAS). 105(14), 2008, S. 5426–5429.
- Rosie Woodroffe, Joshua R. Ginsberg: Edge effects and the extinction of populations inside protected areas. In: Science. 280, 1998, S. 2126–2128. (PDF)
- Lars A. Brudviga, Ellen I. Damschena, Joshua J. Tewksbury, Nick M. Haddad, Douglas J. Levey: Landscape connectivity promotes plant biodiversity spillover into non-target habitats. In: Proceedings of the National Academy of Science (PNAS). 106(23), S. 9328–9332. (pnas.org)
- Verhindern Radler den Gen-Austausch? In: Frankenpost. 13. Mai 2019.
- Lenore Fahrig, Trina Rytwinski: Effects of Roads on Animal Abundance. (ecologyandsociety.org)
- R. Burkhardt u. a.: Empfehlungen zur Umsetzung des § 3 BNatSchG 'Biotopverbund'. Ergebnisse des Arbeitskreises 'Länderübergreifender Biotopverbund' der Länderfachbehörden mit dem BfN. (= Naturschutz und biologische Vielfalt. Heft 2). 2004, ISBN 3-7843-3902-6.
- Towards Green Infrastructure for Europe. In: Proceedings of the European Commission workshop 2009. green-infrastructure-europe.org (Memento vom 13. Juli 2011 im Internet Archive)
- The Green Belt Initiative. (Memento vom 20. Dezember 2010 im Internet Archive)
- V. Ecologische Hoofdstructuur. (uni-muenster.de (Memento vom 9. Juli 2016 im Internet Archive))
- Bundeswildwegeplan des NABU (PDF)
- Wildkatzenwegeplan des BUND (wildkatzenwegeplan.de)
- phi: Damit die Gottesanbeterin weiß, wo’s lang geht. In: badische-zeitung.de, Nachrichten, Südwest, 28. April 2012. (2. Mai 2012)
- Ellen I. Damschen, Nick M. Haddad, John L. Orrock, Joshua J. Tewksbury, Douglas J. Levey: Corridors increase plant species richness at large scales. In: Science. 313, 2006, S. 1284–1286. (sciencemag.org)
- Axel Ssymank, Sandra Balzer, Karin Ullrich: Biotopverbund und Kohärenz nach Artikel 10 der Fauna-Flora-Habitat-Richtlinie. Bundesamt für Naturschutz, 2005. (PDF)
- M. Kettunen, A. Terry, G. Tucker, A. Jones: Guidance on the maintenance of landscape features of major importance for wild flora and fauna - Guidance on the implementation of Article 3 of the Birds Directive (79/409/EEC) and Article 10 of the Habitats Directive (92/43/EEC). Institute for European Environmental Policy (IEEP), Brussels 2007. (PDF)